Contribution of the Belgian school of embryology to the concept of neural induction by the organizer
Full text
Figure

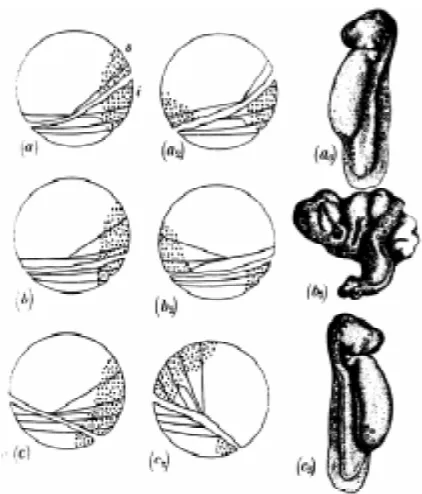
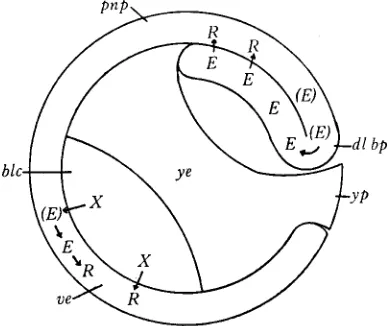
Related documents
National Conference on Technical Vocational Education, Training and Skills Development: A Roadmap for Empowerment (Dec. 2008): Ministry of Human Resource Development, Department
• Follow up with your employer each reporting period to ensure your hours are reported on a regular basis?. • Discuss your progress with
The Integrated Catchment Model INCA has been applied to the whole Thames System to assess a range of scenarios and mitigation measures (Crossman et al., 2013) and the likely
Table B.1.: The set of stimuli word pairs (translated into English) used in the perception experiment — 50 concepts from 10 concept classes and the corresponding (valid or
She says in Texas people don’t use the phrase to run the sweeper when they mean to vacuum the floor, they just say, I have to vacuum the floor (although I’m thinking since in
This study aimed to analyze the performance of all of Palestinian Commercial Banks for the year 2014 using CAMEL approach, to evaluate Palestinian Banks’ capital adequacy,
This paper addresses the effect of coexistence of FDD-LTE networks in 2.6GHz frequency band employing MIMO-OFDM antenna systems in Rayleigh channel.. The
It was decided that with the presence of such significant red flag signs that she should undergo advanced imaging, in this case an MRI, that revealed an underlying malignancy, which
