XX(X) 1 –7 © The Author(s) 2012 Reprints and permission: sagepub.com/journalsPermissions.nav DOI: 10.1177/0963721411434977 http://cdps.sagepub.com
You’ve probably heard it said that the average person uses only 10% of his or her brain. Although this claim is unfounded, it is true that brain-imaging studies have shown that most tasks differentially activate only small patches of the brain—often far less than 10% of the overall gray matter. However, if we could see the results from these imaging studies pooled together (see, e.g., Cabeza & Nyberg, 2000), or, better yet, if we could watch in real time the dynamic and sweeping shifts of activity that occur as the brain performs a complex cognitive task (Kounios et al., 2008), it would be readily apparent that humans can, and do, use the entire spa-tial extent of the brain and the incredible range of mental faculties that it supports.
Still, the seemingly scattered results from individual brain-imaging studies offer an important lesson: Although the full resources of our brain are available to us at any given moment, we don’t (and shouldn’t) use them all at once. But how do our brains learn which specific areas to activate and when to acti-vate them? A fundamental feature of human cognition is the ability to selectively recruit specific brain circuits so that only those areas specialized to deal with the momentary demands of a task are engaged. The ability to dynamically and selec-tively activate specific brain areas in a coordinated fashion is a function of the brain’s learning and cognitive-control architec-ture. That architecture is the focus of this article.
A Triarchic Theory of Learning
Brain research has revealed patterns of experience-dependent change (or plasticity) that occur in the brain over the course of learning. One of the most important clues to understanding the organization of the human brain is the fact that experience does not have a uniform effect across brain regions. Instead, different brain systems demonstrate different patterns of plas-ticity. As people learn and become more skilled at performing specific tasks, they typically use less of the brain’s overall resources, but some areas “drop out” much earlier than others (Chein & Schneider, 2005). In brain areas responsible for stor-ing task-relevant representations, learnstor-ing can have the oppo-site effect of increasing the magnitude and extent of local activity (Ungerleider, Doyon, & Karni, 2002).
By exploring the timeline of changes in the brain over the course of learning (and by drawing on supporting evidence from research on brain connectivity), we have developed a framework for understanding the shifting role of specific brain networks across separate stages of learning—from the initial Corresponding Author:
Jason M. Chein, Temple University, 825 Weiss Hall, 1701 N. 13th St., Philadelphia, PA 19122
E-mail: [email protected]
The Brain’s Learning and
Control Architecture
Jason Chein
1and Walter Schneider
21Department of Psychology, Temple University and 2Learning Research and Development Center,
Department of Psychology, University of Pittsburgh Abstract
Many brain-imaging studies are designed with the goal of isolating brain regions responsible for a specific mental function. The results, which reveal islands of activity scattered about the brain, can give the impression that the brain is just a disorganized collection of specialized processing centers. However, examination of how brain activity changes as a new skill is learned reveals a structured learning architecture composed of three hierarchically organized systems, each with a distinct role in learning and each characterized by a distinct pattern of learning-dependent plasticity. These systems are a representation system, which supports associative learning; a cognitive control network, which allocates attention during the execution of newly learned behaviors; and a metacognitive system, which guides the establishment of new behavioral routines, monitors the quality of ongoing behaviors, and oversees the transitions from one behavior to another. The combined involvement of these systems allows humans to learn rapidly and to flexibly transfer existing knowledge to novel contexts.
Keywords
formation (acquisition) of a new behavior, through a period of controlled execution of that behavior, and finally to a state in which the learned behavior can be carried out with relative automaticity (see Fig. 1).
This framework, which combines findings from past research on executive control (Anderson, Fincham, Qin, & Stocco, 2008; Baddeley, 1996; Miller & Cohen, 2001; Moscovitch, 1992; Shiffrin & Schneider, 1977) with recent findings from brain sciences, accounts for the remarkable capacity, efficiency, and flexibility of human learning. Humans
are unmatched by other organisms (and artificial learning sys-tems) in their ability to both maintain a vast and intricately organized system of knowledge and rapidly acquire and adjust to new knowledge. For instance, we can not only learn highly complex skills, such as reading, but can also readily alter those skills to accommodate a single instruction, such as to read the last sentence of this paragraph backward. !enod lleW
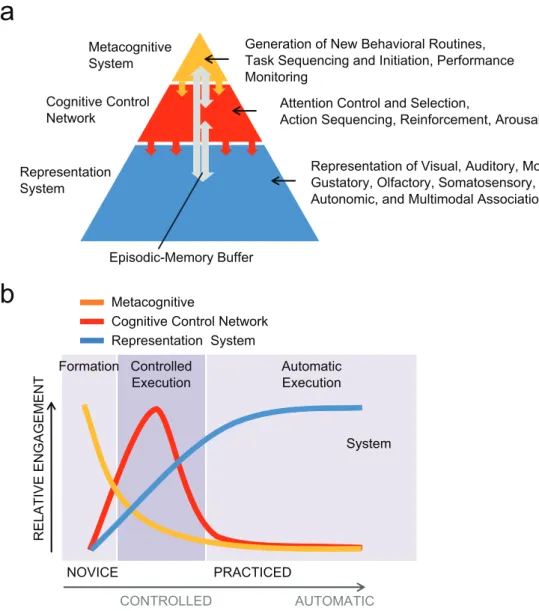
Evidence suggests that the human brain achieves this feat through three hierarchically organized learning systems (Fig. 1a): a representation system, a cognitive control network, and
Generation of New Behavioral Routines, Task Sequencing and Initiation, Performance Monitoring Metacognitive System Cognitive Control Network Representation System Episodic-Memory Buffer
Attention Control and Selection,
Action Sequencing, Reinforcement, Arousal Representation of Visual, Auditory, Motor, Gustatory, Olfactory, Somatosensory, Autonomic, and Multimodal Associations
a
b
CONTROLLED NOVICE AUTOMATIC PRACTICED Automatic Execution Controlled Execution Formation RELATIVE ENGAGEMEN T Metacognitive System Cognitive Control NetworkRepresentation System
Fig. 1. Hierarchical arrangement and major functions of the metacognitive system, the cognitive control network, and the representation system (a), all of which are linked by an episodic-memory buffer. Activity in the three learning systems shifts as people learn and execute skills (b). In an initial formation stage, associated with inexperienced task performance, the metacognitive system aids the learner in establishing the strategies and behavioral routines that support the execution of the task. Once these routines are established, the learner moves into a controlled-execution
stage, during which the cognitive control network is primarily engaged. This system guides action selection and sequencing via attentional control, arousal, and reinforcement signals. During these earlier learning stages, associative mechanisms within the representation system slowly strengthen the key input–output relationships that underlie task execution, gradually allowing cognitive-control resources to disengage as task performance becomes practiced and, eventually, automatic.
a metacognitive system. Each system plays a pivotal and fun-damentally different role in information processing and learn-ing (see Table 1). Performance of a novel task typically proceeds with emphasis shifting from the metacognitive system (formation stage) to the cognitive control network (controlled-execution stage) and, finally, to the representation system (automatic-execution stage; see Fig. 1b). However, we discuss the three systems in the reverse order, which allows us to most clearly highlight their unique properties (the reverse order also reflects their likely sequence of evolutionary devel-opment; see Table 1).
Although we discuss each system separately, we emphasize from the outset that it is their combined and continuously interactive behavior that enables the complexity and flexibility of human learning, and that considering one system in isola-tion from the others may obscure their synergistic nature (a point to which we return in the final section of this article). Ongoing interactions among the three systems are supported by several pathways in the brain, with the medial temporal lobe and associated structures (referred to in our framework as the episodic-memory buffer; see Figs. 1a and 2) functioning as an especially important shared hub in the wiring plan (McClelland, McNaughton, & O’Reilly, 1995).
The Representation System
We refer to the brain’s essential substrate for learning about the world as the representation system. It is within this system that memories and basic knowledge are acquired and stored
through the integration of sensory, perceptual, motor, and mul-timodal (i.e., from different sources) associations. The repre-sentation system is composed of many thousands of localized circuits, or modules, which are broadly distributed across the brain’s surface (Fig. 2 shows only a small patch of the representation system that is implicated in simple visual processing).
The fundamental role of the representation system is to capture the regularities and contingencies of people’s experi-ences by making simple associations: Input set X goes with output Y—for example, seeing a face from any of several angles can lead to identification of a specific individual. The representation system serves as the principle repository for accrued knowledge. There is considerable evidence that learn-ing in the representation system operates on the basic principle that when brain cells fire together (because they are simultane-ously activated by particular inputs), they wire together; this idea is referred to as Hebbian theory or connectionism.
Importantly, the representation system is a slow learner. That is, the neural changes that support associative learning in the representation system occur gradually and incremen-tally—over numerous episodes of learning, often over many days. This slow learning system is helped along by faster learning mechanisms in the hippocampus (a structure known to be crucial in declarative learning and memory; see, e.g., McClelland et al., 1995) and basal ganglia (structures known to support procedural learning; see, e.g., Pascual-Leone, Grafman, & Hallett, 1995). Slow learning in the representation system is important because it assures that the critical information stored
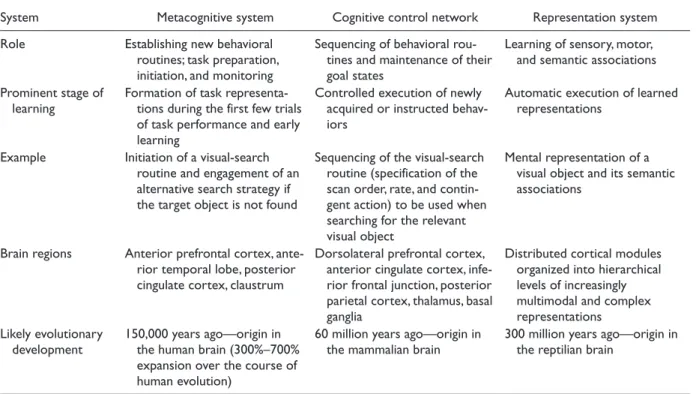
Table 1. Brain Systems Supporting Human Learning
System Metacognitive system Cognitive control network Representation system
Role Establishing new behavioral
routines; task preparation, initiation, and monitoring
Sequencing of behavioral rou-tines and maintenance of their goal states
Learning of sensory, motor, and semantic associations Prominent stage of
learning Formation of task representa-tions during the first few trials
of task performance and early learning
Controlled execution of newly acquired or instructed behav-iors
Automatic execution of learned representations
Example Initiation of a visual-search
routine and engagement of an alternative search strategy if the target object is not found
Sequencing of the visual-search routine (specification of the scan order, rate, and contin-gent action) to be used when searching for the relevant visual object
Mental representation of a visual object and its semantic associations
Brain regions Anterior prefrontal cortex,
ante-rior temporal lobe, posteante-rior cingulate cortex, claustrum
Dorsolateral prefrontal cortex, anterior cingulate cortex, infe-rior frontal junction, posteinfe-rior parietal cortex, thalamus, basal ganglia
Distributed cortical modules organized into hierarchical levels of increasingly multimodal and complex representations
Likely evolutionary
development 150,000 years ago—origin in the human brain (300%–700%
expansion over the course of human evolution)
60 million years ago—origin in
in the system isn’t corrupted by isolated or anomalous experi-ences (McClelland et al., 1995). However, an important fea-ture of the representation system is that it can eventually learn to process information without intervention from attentional mechanisms belonging to the cognitive control network, thereby speeding responses to familiar or emotionally salient stimuli and ultimately leading to automaticity (Schneider & Chein, 2003).
Though several excellent studies have demonstrated plas-ticity in the representation system (Ungerleider et al., 2002), experience-dependent changes in the system can be difficult to spot using brain-imaging techniques because essentially the same set of representation modules are engaged to support both novice and skilled (automatic) performance (Chein & Schneider, 2005) and because learning accrues slowly in this system. In contrast, the profound influence of learning on activity in the cognitive control network is readily observed in neuroimaging studies.
The Cognitive Control Network
The cognitive control network is a domain-general system responsible for monitoring and directing information process-ing in the representation system. The cognitive control network guides attention to information that is most relevant for further processing. It thus forms a system for working memory that allows people to maintain goals and to bias mental processing in accord with those goals and the current context. The cognitive control network also possesses mechanisms that allow people to keep irrelevant information out of mind and to update and sequence action so that they can carry out complex, goal-directed behavioral routines. Though it often behaves as a coor-dinated network, the cognitive control network is made up of many separate brain areas (Table 1; Fig. 2), each with special-ized roles (Cole & Schneider, 2007; Schneider & Chein, 2003). Perhaps the most prominent node of the cognitive control network is the lateral prefrontal cortex. Some accounts of
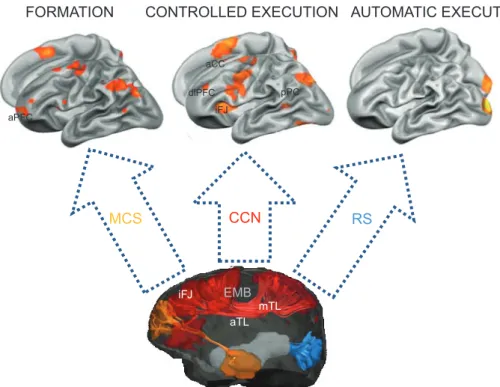
EMB aPFC dlPFC pPC aCC mTL aTL iFJ aPFC dlPFC aCC iFJ pPC CONTROLLED EXECUTION FORMATION MCS CCN RS AUTOMATIC EXECUTION
Fig. 2. Key brain areas and internal connection pathways of the three learning systems in the brain (some areas and pathways not depicted). The illustrations of functional MRI activation maps (top) show how activity shifts across stages of learning in a simple visual-discrimination task. Initial performance, or
formation, is associated with increased activity in the anterior prefrontal cortex (aPFC), a key component of the metacognitive system (MCS). After the first few trials, activity declines in the aPFC and increases in the interconnected regions of the cognitive control network (CCN)—the dorsolateral prefrontal cortex (dlPFC), the anterior cingulate cortex (aCC), the posterior parietal cortex (pPC), and the inferior frontal junction (iFJ)—to support controlled execution of the task. After considerable practice, automatic execution of the task becomes possible. At this point, activity declines in cognitive-control areas and task performance is primarily dependent on areas of the representation system (RS) that support the learned visual input to response output relationships (note that the RS activity in this illustration is specific to low-level visual processing). The illustration of a diffusion-weighted image (bottom) shows some of the major internal connection pathways that enable coordinated action within each learning system and integration across systems through the episodic-memory buffer (EMB).
prefrontal organization and function treat this area not as the center for a specific control process but simply as another sub-strate for the representation of learned associations (see, e.g., Wood & Grafman, 2003). It may thus seem odd to conceive of the cognitive control network as a distinct learning system. Why draw the distinction between learning in the cognitive control network and learning in the representation system?
One important difference is the relative complexity of the information represented within the cognitive control network. Rather than simple associations, lateral prefrontal representa-tions seem to reflect intricate, multistep routines that guide sequenced behaviors. Perhaps an even more fundamental dis-tinction is that whereas the representation system represents specific learned associations, the complex routines represented in the cognitive control network can incorporate varying infor-mation. That is, any of several values may be assigned to the variables in a cognitive-control-network routine. For example, if a represented behavior is “continue to read this paper until X,” the value of X might be “an e-mail arrives” or “it is 4 o’clock.” This variable-binding property of the cognitive control network is essential because it allows learned behav-iors to be applied in novel environments and to be adapted to accommodate novel information (Rougier, Noelle, Braver, Cohen, & O’Reilly, 2005).
Somewhat paradoxically, activity in the cognitive control network can change because of learning that occurs in the rep-resentation system rather than in the cognitive control network itself. When the representation system processes novel or unfamiliar information, the cognitive control network is strongly engaged to direct attentional signaling toward task- and goal-relevant representations (Miller & Cohen, 2001). That is, when a task is not well-learned, the cognitive control network makes sure the brain stays focused on the right infor-mation. Processing in the cognitive control network is thus essential for scaffolding the early performance and learning of almost any task. However, as specific associations in the rep-resentation system become well-learned, they can evoke fur-ther processing on the basis of the information they carry without having to wait for the attentional “go” signal from the cognitive control network. Accordingly, the role of the cogni-tive control network is reduced as the representation system learns to manage information on its own.
Evidence of the broad but diminishing role of the cognitive control network during the transition from controlled to auto-matic processing has come from numerous neuroimaging studies comparing inexperienced to skilled performance. Studies using widely varying tasks and materials have shown a consistent pattern of strong activation in cognitive-control-network regions during early performance and comparatively weak activation in these brain regions once a task has become well-learned (Chein & Schneider, 2005).
Although the experience-dependent changes witnessed in the transition from controlled to automatic processing do not reflect plasticity in the cognitive control network, this system itself undergoes change. For example, developmental-imaging
studies (Luna, Padmanabhan, & O’Hearn, 2010) have sug-gested that brain regions in the cognitive control network expe-rience maturation that (a) causes them to become progressively more segregated from surrounding brain areas, (b) improves their coordinated action, and (c) affords a greater ability to gov-ern activity in more distal brain regions.
Recent studies have similarly indicated that training, not just maturation, can play an important role in shaping and enhancing the capacity for cognitive control by increasing the functional efficacy of the regions in the cognitive control net-work (Hill & Schneider, 2006). Moreover, different types of experience may enhance the adeptness of different processes associated with cognitive control. For example, repetition of appropriately designed mental exercises can increase the abil-ity to update active mental representations (Dahlin, Neely, Larsson, Backman, & Nyberg, 2008), to multitask (Dux et al., 2009), and to resist interference from distracting sources of information (Chein & Morrison, 2010; Richmond, Morrison, Chein, & Olson, 2011). These effects of behavioral training correspond with a strengthened engagement of specific brain regions within the cognitive control network (Dahlin, Backman, Neely, & Nyberg, 2009; Dux et al., 2009) and impart different advantages to other aspects of cognitive per-formance (for reviews, see Klingberg, 2010; Morrison & Chein, 2011; Tang & Posner, 2009).
The Metacognitive System
Recent research has provided early evidence indicating that a third system is important to human learning: the metacognitive system. In our framework, this system sits above the cognitive control network (Fig. 1a) and plays an important role in both reconfiguring the brain as it prepares to execute existing behav-ioral routines and monitoring behavior during the acquisition of new routines. The metacognitive system can thus be thought of as a control system that regulates the behavior of the cognitive control network. The metacognitive system might, for example, initiate or terminate a cognitive-control-network routine, or it might modify the settings of a cognitive-control-network rou-tine in order to improve learning and performance.
The region most clearly implicated as a component of the metacognitive system is found at the very front of the brain in the anterior prefrontal cortex (see Fig. 2), but this system may also include areas in the anterior half of the temporal lobes and regions closer to the midline of the brain, including the poste-rior cingulate cortex and claustrum. We speculate that indi-viduals who possess a strong ability to perform in novel contexts may have an especially well-developed metacogni-tive system, which allows them to rapidly acquire new behav-ioral routines and to consider the likely effectiveness of alternative learning strategies (e.g., rote rehearsal vs. generat-ing explanations to oneself; Chi, 2000).
We differentiate between the metacognitive system and the cognitive control network on the basis of three key sources of evidence. First, recent work has suggested that the
anterior-posterior (front to back) extent of the prefrontal cortex is organized in a hierarchical gradient (Badre & D’Esposito, 2009). More posterior prefrontal regions appear to enact spe-cific behavioral routines by influencing the flow of information in distal representational areas, a function that is consistent with a role in the cognitive control network. In contrast, more anterior prefrontal regions are implicated in more complex and abstract behaviors and primarily modulate the activity of nearby prefrontal areas (Christoff, Keramatian, Gordon, Smith, & Madler, 2009). The behavior of the anterior prefrontal cortex is thus consistent with a role in controlling a key cognitive-control-network node located further back in the prefrontal cor-tex: the dorsolateral prefrontal cortex (dlPFC; see Fig. 2).
Second, recent neuroimaging work (Cole, Bagic, Kass, & Schneider, 2010; Moss, Schunn, Schneider, McNamara, & VanLehn, 2011; Ruge & Wolfensteller, 2010; Sakai & Passingham, 2006; Strange, Henson, Friston, & Dolan, 2001) has indicated that the anterior prefrontal cortex is especially important in strategy choice when people first begin learning a new rule and in the recalibration of the brain when a previ-ously learned behavior needs to be carried out. Unlike more posterior cognitive-control-network regions that remain active well into learning, activity in the anterior prefrontal cortex declines to baseline after just a few trials in a rule-learning paradigm (Cole et al., 2010).
Third, recent structural imaging of the human connectome (i.e., patterns of neural connections) has shown that the regions of the metacognitive system have a different wiring pattern than do the regions of the cognitive control network. Using diffusion-weighted imaging to examine neural connectivity, we recently found that the anterior prefrontal cortex connects to the anterior portion of the temporal lobe (see Fig. 2) but contains few projections to anterior cingulate or parietal areas (regions of the cognitive control network that are tightly con-nected to the posterior prefrontal cortex).
Learning Systems Working Together
The relative involvement of the three learning systems varies as people transition through stages of learning and experience (Figs. 1 and 2). During the formation stage, the metacognitive system plays an especially prominent role in initiating and calibrating the brain so that it is ready to carry out a new rou-tine. During controlled execution, the cognitive control net-work becomes foremost in importance by directing internal attention in order to control the flow of information process-ing. Finally, when automaticity is attained, activity shifts to the representation system, which has completed its coding of stimulus–response associations, allowing the cognitive control network to loosen its grip. Despite these transitions, the three interlinked systems remain tightly orchestrated, with the metacognitive system monitoring activity in the cognitive control network and upper-level representation system, and the cognitive control network tracking and periodically rerout-ing representation-system activity.
The impressive efficiency of human learning, relative to that of other species, illustrates the fundamental advantage of having evolved these three systems that possess different prop-erties but are capable of operating in a coordinated fashion. With only the associative learning mechanisms (i.e., the mech-anisms comprising the representation system), it would take people thousands of trials to learn even very simple rules. The cognitive control network’s regulation of processing in the associative system accelerates learning by several orders of magnitude, and metacognitive control over these routines fur-ther accelerates initial learning and increases adaptability to changing contextual information. With this triarchic learning-and-control architecture, humans can learn to perform a new task in just a few trials and to perform that task more or less automatically after a few hundred. When needed, humans can alter overlearned behaviors (e.g., reading) to readily accom-modate novel constraints (e.g., by switching from reading for-ward to reading backfor-ward). The fast, flexible, and transferable learning that is supported by this triarchic learning architecture thus provides enormous advantages to the human species. Recommended Readings
Chein, J. M., & Morrison A. B. (2010). Expanding the mind’s work-space: Training and transfer effects with a complex working memory span task. Psychonomic Bulletin & Review, 17, 193–199. A representative study that demonstrates how cognitive-control network training can lead to increased working memory capacity with generalization to other cognitive abilities.
Craik, F., & Bialystock, E. (2006). Cognition through the lifespan: Mechanisms of change. Trends in Cognitive Sciences, 10, 131– 138. A complementary review of the pattern of the changes that occur in brain systems responsible for representation (i.e., the rep-resentation system) and control (i.e., the cognitive control network) over the course of maturation and age-related decline.
Duncan, J. (2010). The multiple-demand (MD) system of the primate brain: Mental programs for intelligent behavior. Trends in Cogni-tive Sciences, 14, 172–179. A comprehensive review of the neu-roscientific evidence implicating the brain’s control networks in the assembly and execution of structured mental programs. Schneider, W., & Chein, J. M. (2003). Controlled and automatic
processing: From mechanisms to biology. Cognitive Science, 27, 525–559. Review of the biological and behavioral evidence distinguishing between controlled processing in the cognitive control network and automatic processing in the representation system.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by the Defense Advanced Research Projects Agency (DARPA) under contract NBCHZ090439. The views, opinions, and findings contained in this article are those of the authors and should not be interpreted as representing the official
views or policies, either expressed or implied, of DARPA or the United States Department of Defense.
References
Anderson, J. R., Fincham, J. M., Qin, Y., & Stocco, A. (2008). A central circuit of the mind. Trends in Cognitive Sciences, 12, 136–143.
Baddeley, A. D. (1996). Exploring the central executive. Quarterly Journal of Experimental Psychology: Human Experimental Psy-chology, 49A, 5–28.
Badre, D., & D’Esposito, M. (2009). Is the rostro-caudal axis of the frontal lobe hierarchical? Nature Reviews Neuroscience, 10, 659–669.
Cabeza, R., & Nyberg, L. (2000). Imaging cognition II: An empiri-cal review of 275 PET and fMRI studies. Journal of Cognitive Neuroscience, 12, 1–47.
Chein, J. M., & Morrison, A. B. (2010). Expanding the mind’s work-space: Training and transfer effects with a complex working memory span task. Psychonomic Bulletin & Review, 17, 193–199. Chein, J. M., & Schneider, W. (2005). Neuroimaging studies of
practice-related change: FMRI and meta-analytic evidence of a domain-general control network for learning. Brain Research: Cognitive Brain Research, 25, 607–623.
Chi, M. (2000). Self-explaining expository texts: The dual pro-cesses of generating inferences and repairing mental models. In R. Glaser (Ed.), Advances in instructional psychology (Vol. 5, pp. 161–238). Mahwah, NJ: Erlbaum.
Christoff, K., Keramatian, K., Gordon, A. M., Smith, R., & Madler, B. (2009). Prefrontal organization of cognitive control according to levels of abstraction. Brain Research, 1286, 94–105.
Cole, M. W., Bagic, A., Kass, R., & Schneider, W. (2010). Prefrontal dynamics underlying rapid instructed task learning reverse with practice. Journal of Neuroscience, 30, 14245–14254.
Cole, M. W., & Schneider, W. (2007). The cognitive control network: Integrated cortical regions with dissociable functions. Neuro- Image, 37, 343–360.
Dahlin, E., Backman, L., Neely, A. S., & Nyberg, L. (2009). Train-ing of the executive component of workTrain-ing memory: Subcortical areas mediate transfer effects. Restorative Neurology and Neuro-science, 27, 405–419.
Dahlin, E., Neely, A. S., Larsson, A., Backman, L., & Nyberg, L. (2008). Transfer of learning after updating training mediated by the striatum. Science, 320, 1510–1512.
Dux, P. E., Tombu, M. N., Harrison, S., Rogers, B. P., Tong, F., & Marois, R. (2009). Training improves multitasking performance by increasing the speed of information processing in human pre-frontal cortex. Neuron, 63, 127–138.
Hill, N. M., & Schneider, W. (2006). Brain changes in the develop-ment of expertise: Neurological evidence on skill-based adapta-tions. In K. A. Ericsson, N. Charness, P. Feltovich, & R. Hoffman (Eds.), Cambridge handbook of expertise and expert performance
(pp. 653–683). New York, NY: Cambridge University.
Klingberg, T. (2010). Training and plasticity of working memory.
Trends in Cognitive Sciences, 14, 317–324.
Kounios, J., Fleck, J. I., Green, D. L., Payne, L., Stevenson, J. L., Bowden, E. M., & Jung-Beeman, M. (2008). The origins of
insight in resting-state brain activity. Neuropsychologia, 46, 281–291.
Luna, B., Padmanabhan, A., & O’Hearn, K. (2010). What has fMRI told us about the development of cognitive control through ado-lescence? Brain and Cognition, 72, 101–113.
McClelland, J. L., McNaughton, B. L., & O’Reilly, R. C. (1995). Why there are complementary learning systems in the hippo-campus and neocortex: Insights from the successes and failures of connectionist models of learning and memory. Psychological Review, 102, 419–457.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefron-tal cortex function. Annual Review of Neuroscience, 24, 167–202. Morrison, A. B., & Chein, J. M. (2011). Does working memory train-ing work? The promise and challenges of enhanctrain-ing cognition by training working memory. Psychonomic Bulletin & Review, 18, 46–60.
Moscovitch, M. (1992). Memory and working-with-memory: A com-ponent process model based on modules and central systems.
Journal of Cognitive Neuroscience, 4, 257–267.
Moss, J., Schunn, C. D., Schneider, W., McNamara, D. S., & Van-Lehn, K. (2011). An fMRI study of strategic reading com-prehension: Cognitive control and discourse comprehension.
NeuroImage, 58, 675–686.
Pascual-Leone, A., Grafman, J., & Hallett, M. (1995). Procedural learning and prefrontal cortex. Annals of the New York Academy of Sciences, 769, 61–70.
Richmond, L. L., Morrison, A. B., Chein, J. M., & Olson, I. R. (2011). Working memory training and transfer in older adults. Psychol-ogy and Aging, 26, 813–822.
Rougier, N. P., Noelle, D. C., Braver, T. S., Cohen, J. D., & O’Reilly, R. C. (2005). Prefrontal cortex and flexible cognitive control: Rules without symbols. Proceedings of the National Academy of Sciences, USA, 102, 7338–7343.
Ruge, H., & Wolfensteller, U. (2010). Rapid formation of pragmatic rule representations in the human brain during instruction-based learning. Cerebral Cortex, 20, 1656–1667.
Sakai, K., & Passingham, R. E. (2006). Prefrontal set activity predicts rule-specific neural processing during subsequent cognitive per-formance. Journal of Neuroscience, 26, 1211–1218.
Schneider, W., & Chein, J. M. (2003). Controlled and automatic pro-cessing: Behavior, theory, and biological mechanisms. Cognitive Science, 27, 525–559.
Shiffrin, R. M., & Schneider, W. (1977). Controlled and automatic human information processing: II. Perceptual learning, automatic attending and a general theory. Psychological Review, 84, 127–190. Strange, B. A., Henson, R. N., Friston, K. J., & Dolan, R. J. (2001).
Anterior prefrontal cortex mediates rule learning in humans.
Cerebral Cortex, 11, 1040–1046.
Tang, Y. Y., & Posner, M. I. (2009). Attention training and attention state training. Trends in Cognitive Sciences, 13, 222–227. Ungerleider, L. G., Doyon, J., & Karni, A. (2002). Imaging brain
plasticity during motor skill learning. Neurobiology of Learning and Memory, 78, 553–564.
Wood, J. N., & Grafman, J. (2003). Human prefrontal cortex: Pro-cessing and representational perspectives. Nature Reviews Neuro- science, 4, 139–147.