0099-2240/11/$12.00 doi:10.1128/AEM.06069-11
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Identification and Characterization of a Novel Class of Extracellular
Poly(3-Hydroxybutyrate) Depolymerase from
Bacillus
sp.
Strain NRRL B-14911
䌤
Wan-Ting Ma, Ju-Hui Lin, Hui-Ju Chen, Syuan-Yi Chen, and Gwo-Chyuan Shaw*
Institute of Biochemistry and Molecular Biology, School of Life Science, National Yang-Ming University, Taipei, Taiwan, Republic of China
Received 3 July 2011/Accepted 14 September 2011
The catalytic, linker, and denatured poly(3-hydroxybutyrate) (dPHB)-binding domains of bacterial extracellular PHB depolymerases (PhaZs) are classified into several different types. We now report a novel class of extracellular PHB depolymerase from Bacillus sp. strain NRRL B-14911. Its catalytic domain belongs to type 1, whereas its putative linker region neither possesses the sequence features of the three known types of linker domains nor exhibits significant amino acid sequence similarity to them. Instead, this putative linker region can be divided into two distinct linker domains of novel types: LD1 and LD2. LD1 shows significant amino acid sequence similarity to certain regions of a large group of PHB depolymerase-unrelated proteins. LD2 and its homologs are present in a small group of PhaZs. The remaining C-terminal portion of this PhaZ can be further divided into two distinct domains: SBD1 and SBD2. Each domain showed strong binding to dPHB, and there is no significant sequence similarity between them. Each domain neither possesses the sequence features of the two known types of dPHB-binding domains nor shows significant amino acid sequence similarity to them. These unique features indicate the presence of two novel and distinct types of dPHB-binding domains. Homologs of these novel domains also are present in the extracellular PhaZ ofBacillus megateriumand the putative extracellular PhaZs ofBacillus pseudofirmusandBacillussp. strain SG-1. TheBacillussp. NRRL B-14911 PhaZ appears to be a representative of a novel class of extracellular PHB depolymerases.
Poly(3-hydroxybutyrate) (PHB) is synthesized and accumu-lated intracellularly as a carbon and energy storage material by a wide variety of bacteria (14, 28, 29). When nutrient availabil-ity is restricted or unbalanced, the accumulated PHB is subject to degradation by the intracellular PHB depolymerase(s) (PhaZ) to produce 3-hydroxybutyrate (3HB) oligomers and/or monomers. These products subsequently are metabolized by the intracellular 3HB-oligomer hydrolase and/or the 3HB de-hydrogenase as carbon and energy sources (21, 22, 30, 36, 38). The intracellular PHB exists in an amorphous state. When the amorphous PHB is subjected to solvent extraction or exposed to the extracellular environment due to cell death or lysis, the amorphous PHB is transformed into denatured semicrystalline PHB (dPHB) (13, 16).
A wide variety of bacteria can produce and secrete extra-cellular PHB depolymerases for the specific degradation of dPHB (12, 16, 40), with some exceptions. One exception is
the extracellular PHB depolymerase PhaZ7 ofPaucimonas
lemoignei, which can only degrade amorphous PHB (7). Another known exception is the periplasm-located PHB
de-polymerase of Rhodospirillum rubrum, which shows
sub-strate specificity for amorphous PHB (8). Like intracellular PHB depolymerases, extracellular PHB depolymerases can degrade PHB into 3HB oligomers and/or monomers. The
produced 3HB oligomers can be further degraded by extra-cellular 3HB-oligomer hydrolases into 3HB monomers or degraded by intracellular 3HB-oligomer hydrolases after the uptake of 3HB oligomers into bacterial cells (36, 43).
The mature forms of extracellular PHB depolymerases usu-ally are composed of three domains: a catalytic domain, a linker domain, and one or two dPHB-binding domains (16, 35). The catalytic domain is classified into type 1 and type 2 sub-groups, depending on the relative order of the histidine residue for the oxyanion hole and the catalytic triad residues. The linker domain is classified into three subgroups according to their sequence features or their similarities to other proteins: the cadherin-like (19, 25), the fibronectin type III-like (18, 20, 24, 34), and the threonine-rich linker domains (3, 15). The dPHB-binding domain is classified into two subgroups: type 1 and type 2 (16). There also are single-domain extracellular
PHB depolymerases: the PhaZ7 ofP. lemoignei(26) and the
PhaZ ofPenicillium funiculosum(10). These PhaZs contain
only a unique catalytic domain. In this study, we have iden-tified a novel class of extracellular PHB depolymerase from
Bacillussp. strain NRRL B-14911. This PhaZ contains two novel and distinct types of linker domains (LD1 and LD2), as well as two novel and distinct types of dPHB-binding domains (SBD1 and SBD2). Homologs of these novel
do-mains also are present in the extracellular PhaZ ofBacillus
megateriumand the putative extracellular PhaZs ofBacillus pseudofirmusandBacillussp. SG-1. TheBacillussp. NRRL B-14911 PhaZ appears to be a representative of a novel class of extracellular PHB depolymerases.
* Corresponding author. Mailing address: Institute of Biochemistry and Molecular Biology, School of Life Science, National Yang-Ming University, Taipei 112, Taiwan. Phone: 2826-7127. Fax: 886-2-2826-4843. E-mail: [email protected].
䌤Published ahead of print on 23 September 2011.
7924
on January 24, 2021 by guest
http://aem.asm.org/
MATERIALS AND METHODS
Bacterial strains and growth conditions.The bacterial strains and plasmids used in this study are listed in Table 1. The oligonucleotide primers are listed in Table 2. Escherichia colicells were grown at 37°C in Luria-Bertani (LB) medium (31).E. coli strain DH5␣was used as a host for cloning purposes;E. colistrain BL21(DE3) and strain JM109 were used as hosts for the heterologous expression of proteins.Bacillus
sp. NRRL B-14911 cells were grown at 37°C in LB medium for the isolation of the genomic DNA. Overlay agar plates that were made with M9 mineral salts medium (31) and supplemented with 0.6% dPHB in the top agar or overlay agar plates that were made with LB medium and supplemented with 0.6% dPHB in the top agar were prepared as described previously (17) and were used for the observation of halo formation by dPHB-degrading bacteria. Antibiotics were used at the following con-centrations (g/ml): ampicillin, 100 (forE. coli); tetracycline, 10 (forB. subtilis).
Construction of plasmids.To construct plasmid pGS2067, the insert DNA was amplified by PCR using the primer pair A098 plus A099, restriction digested, and inserted between the EcoRI and HindIII sites of pHY300PLK (Takara).
To construct plasmids pGS2164, pGS2183, and pGS2188, primer pairs A397 plus A174, A398 plus A174, and A397 plus A399, respectively, were used in PCR. The DNA fragments were individually cloned between the NdeI and XhoI sites of pET22b (Novagen).
To construct plasmid pGS2218, primer pairs A397 plus A467 and A468 plus A174 were used in PCR. After restriction digestion with NdeI plus EcoRI and EcoRI plus XhoI, respectively, these two DNA fragments were ligated together into the NdeI and XhoI sites of pET22b. Plasmid pGS2281 was constructed in a similar manner by using the primer pairs A397 plus A467 and A556 plus A174. To construct plasmids pGS2230, pGS2247, pGS2248, and pGS2249, primer pairs A468 plus A099, A468 plus A555, A556 plus A557, and A558 plus A099, respectively, were used in PCR. The DNA inserts were individually cloned between the EcoRI and HindIII sites of pET32a (Novagen). To construct plas-mid pGS2254, the primer pair A551 plus A568 was used in PCR. The insert DNA was cloned between the BamHI and HindIII sites of pET32a.
TABLE 1. Bacterial strains and plasmids
Strain or plasmid Descriptiona Reference or
sourceb
E. coli
DH5␣ F⫺80dlacZ⌬M15⌬(lacZYA-argF)recA1 gyrA endA1 relA1 supE44 hsdR17 Laboratory stock
JM109 recA1 supE44 endA1 hsdR17 gyrA96 relA1 thi⌬(lac-proAB)关F⬘traD36 proAB⫹lacIqZ⌬M15兴 Takara
BL21(DE3) F⫺ompT hsdS gal dcm(DE3) Novagen
B. subtilis
168 trpC2 Laboratory stock
BM1269 trpC2(pGS2067) This study
BM548 trpC2(pHY300PLK) This study
Bacillussp.
NRRL B-14911 ARSCC
B. megaterium
ATCC 11561 ATCC
Plasmids
pHY300PLK Expression vector; Apr, Tcr Takara
pQE30 Expression vector for producing His-tagged proteins inE. coliJM109; Apr Qiagen pET22b Expression vector for producing His-tagged proteins inE. coliBL21(DE3); Apr Novagen pET32a Expression vector for producing His-tagged thioredoxin fusion proteins inE. coliBL21(DE3); Apr Novagen pGS1243 pQE30 carrying the NAD⫹-dependent (R)-3HB dehydrogenase gene ofB. thuringiensis 40 pGS2067 pHY300PLK carrying DNA encoding the full-length PhaZ(BspNRRL) This study pGS2164 pET22b carrying DNA encoding the mature form of PhaZ(BspNRRL) This study pGS2183 pET22b carrying DNA encoding the C domain of PhaZ(BspNRRL) This study pGS2188 pET22b carrying DNA encoding the N domain of PhaZ(BspNRRL) This study pGS2218 pET22b producing the mature form of PhaZ(BspNRRL) with deletion of the linker domain LD1 This study pGS2230 pET32a producing the fusion protein of TrxA and the C domain of PhaZ(BspNRRL) with
deletion of the linker domain LD1
This study pGS2247 pET32a producing the fusion protein of TrxA and the linker domain LD2 of PhaZ(BspNRRL) This study pGS2248 pET32a producing the fusion protein of TrxA and the dPHB-binding domain SBD1 of
PhaZ(BspNRRL)
This study pGS2249 pET32a producing the fusion protein of TrxA and the dPHB-binding domain SBD2 of
PhaZ(BspNRRL)
This study pGS2254 pET32a producing the fusion protein of TrxA and the linker domain LD1 of PhaZ(BspNRRL) This study pGS2281 pET22b producing the mature form of PhaZ(BspNRRL) with deletion of the linker domains LD1
and LD2 This study a Apr , ampicillin resistant; Tcr , tetracycline resistant. b
ARSCC, Agricultural Research Service Culture Collection; ATCC, American Type Culture Collection.
TABLE 2. Oligonucleotide primers used in this study Name Sequence (5⬘–3⬘)a Location
A098 GCGGAATTCGAATGAAAAGGAGGAATGG PhaZ 5⬘end A099 GCCAAAGCTTCAAAGGGAAAAAAGAGAG PhaZ 3⬘end A174 GCCACTCGAGGTTGGCGCGATTTACTAT SBD2 3⬘end A397 GCCAACACATATGGCAGGACAATTTATTACAG N 5⬘end A398 GCCAACACATATGGATGCCCCTGCTACATC LD1 5⬘end A399 GCCACTCGAGGCCGGCATCTCCTCCA N 3⬘end A467 GCGGAATTCGCCGGCATCTCCTCCA N 3⬘end A468 GCGGAATTCAGCGCCATCTCAGG LD2 5⬘end A551 GCGGGATCCGATGCCCCTGCTACATC LD1 5⬘end A555 GCCAAAGCTTTCACGCTGTCTGTCCGAGC LD2 3⬘end A556 GCGGAATTCATTGGGTCAGCAAAATTGC SBD1 5⬘end A557 GCCAAAGCTTTCAAACGGCGGCCGGCTG SBD1 3⬘end A558 GCGGAATTCTCTAATTACATTGATATAGATA SBD2 5⬘end A568 GCCAAAGCTTTCATGTCAGCGAGTCCCCG LD1 3⬘end
a
Underlined sequences represent restriction sites inserted.
on January 24, 2021 by guest
http://aem.asm.org/
All sequences of PCR-amplified DNA fragments were confirmed by DNA sequencing.
Overproduction and purification of His-tagged proteins.For the purification of His-tagged proteins overproduced fromE. colistrain BL21(DE3) bearing plasmid pGS2164, pGS2183, pGS2188, pGS2230, pGS2249, or pGS2254, cells were grown in LB medium at 37°C to an absorbance at 600 nm of 0.5 and then treated with 0.3 mM IPTG (isopropyl--D-thiogalactopyranoside) at 37°C for 3 h. Cells were collected, disrupted by sonication, and centrifuged at 15,000⫻gfor 10 min. The resulting supernatant was used for the purification of His-tagged proteins by affinity chromatography on an Ni-nitrilotriacetic acid (NTA) agarose column according to the instructions of the matrix manufacturer (Qiagen Inc.). A similar procedure was followed to overproduce and purify His-tagged thiore-doxin itself fromE. colistrain BL21(DE3) bearing the expression vector pET32a (Novagen).
For the purification of His-tagged proteins overproduced fromE. colistrain BL21(DE3) bearing plasmid pGS2218, pGS2247, or pGS2248, cells were grown in LB medium at 37°C to an absorbance at 600 nm of 0.5 and then treated with 0.02 mM IPTG at 16°C for 16 h. The purification of His-tagged NAD⫹-dependent (R)-3HB dehydrogenase ofB. thuringiensisfrom E. coli strain JM109 bearing the plasmid pGS1243 was carried out exactly as de-scribed previously (41).
Preparation of dPHB.dPHB was isolated fromB. megateriumcells that were grown at 37°C in LB medium supplemented with 2% sodium acetate for 8 h by a procedure involving sodium hypochlorite digestion and subsequent solvent extraction with acetone-ether (2:1, vol/vol) as described previously (6).
Determination of the enzyme activities of PHB depolymerase and its deriva-tives.The enzyme activity of PHB depolymerase or its derivative was determined by monitoring the decrease in the turbidity of a suspension made from dPHB due to the hydrolysis of dPHB as described previously (9). The turbidity of the reaction mixture was assayed spectrophotometrically at 650 nm by using a Spec-traMax 190 microplate reader (Molecular Devices Corp.). The reaction mixture (200l) contained 100 mM Tris-HCl (pH 8.0), 0.7 mg of dPHB, and 0.24M purified PHB depolymerase or its derivative, and it was incubated at 37°C for 30 min. One unit of activity is defined as a decrease of 1 U in the optical density at 650 nm (OD650) in 1 min.
To investigate the effects of various chemical reagents on the dPHB-hydro-lyzing activity of PhaZ, the chemical reagent was preincubated with PhaZ in the reaction mixture for 10 min prior to the addition of dPHB.
The spectrophotometric measurement of yellow p-nitrophenol generated from the hydrolysis of p-nitrophenylbutyrate by PhaZ or its derivative was performed exactly as described previously (41).
Quantitation of dPHB and 3HB monomers.The determination of the amount of dPHB was carried out as described previously (5). The amount of 3HB monomers produced from the hydrolysis of dPHB by PHB depolymerase was quantified by the enzymatic method using NAD⫹-dependent (R)-3HB dehydro-genase according to the description in a previous report (42) and as reported previously by us (41).
Western blot analysis of the dPHB- or chitin-binding ability.PHB depoly-merase or its derivative was pretreated with 1 mM PMSF (phenylmethylsul-fonyl fluoride) for 10 min to inhibit the degrading activity. The dPHB-or chitin-binding assay mixture (13l) contained 10 mM Tris-HCl (pH 7.4), 200g of dPHB or chitin (Sigma), and 1 to 3g of PMSF-treated PHB depolymerase or its derivative. After incubation at 30°C for 15 min, the assay mixture was separated into the supernatant and the dPHB- or chitin-contain-ing pellet by centrifugation. The pellet was washed three times with 40l of 10 mM Tris-HCl (pH 7.4) at room temperature. Proteins in the washed pellet and in the supernatant from the assay mixture were analyzed by sodium dodecyl sulfate-12% polyacrylamide gel electrophoresis (SDS-12% PAGE) and Western blotting with anti-His tag polyclonal antibody (Santa Cruz Biotechnology, Inc.) as the probe. Western blotting was carried out exactly as described previously (5).
Protein sequence analysis. Amino acid identities and similarities between proteins were calculated with the CLUSTAL W software (39). Signal peptide sequences were analyzed by the SignalP software (23). The isoelectric point (pI) was calculated by using the ExPASy proteomics server of the Swiss Institute of Bioinformatics.
Other methods.The genomic DNA ofBacillussp. NRRL B-14911 was isolated by the method described previously (27). The transformation ofB. subtiliscells by the protoplast method was performed as described previously (4). Protein concentrations were determined by the bicinchoninic acid protein assay method according to the instructions of the assay kit manufacturer (Pierce Biotechnol-ogy, Inc.) with bovine serum albumin as the standard.
Amino acid sequence accession numbers.Amino acid sequences for the following proteins have been submitted to GenBank (accession numbers are in parentheses): PHB depolymerases of Bacillus sp. NRRL B-14911 (ZP_01169502), Bacillus sp. SG-1 (ZP_021858765),B. pseudofirmus OF4 (YP_003425754), andB. megateriumQM B1551 (YP_003560966); NHL re-peat-containing protein ofGeobactersp. FRC-32 (YP_002537077); FG-GAP repeat-containing protein ofArthrobacter aurescensTC1 (YP_946834); and Ig domain protein group 2 domain protein of Thermincola sp. JR (YP_003641592).
RESULTS
Identification of a putative extracellular PHB depolymerase
from Bacillus sp. NRRL B-14911. The genomic sequence of
Bacillussp. NRRL B-14911 has been partially determined and is available from GenBank. It encodes a putative PHB depoly-merase (GenBank accession number ZP_01169502) of 592 amino acids (see Discussion) that shows 53% identity and 68%
similarity to the extracellular PHB depolymerase ofB.
mega-teriumN-18-25-9 (590 amino acids; BAF35850) (37). The
anal-ysis of the putative PhaZ ofBacillussp. NRRL B-14911 by the
SignalP 3.0 software revealed the presence of a typical signal peptide (100% probability) from amino acid position 1 to 28. The most likely cleavage site is predicted to be located between Ala28 and Ala29 (99.7% probability). These features suggest that this putative PhaZ is an extracellular enzyme. When grow-ing on an opaque overlay agar plate that was prepared with M9 mineral salts medium and supplemented with 0.6% dPHB in
the top agar, Bacillus sp. NRRL B-14911 cells produced a
clearing zone (Fig. 1A). This observation indicates the
pres-ence of dPHB depolymerase activity in Bacillus sp. NRRL
B-14911. When the pHY300PLK-based plasmid pGS2067,
which constitutively expressed the full-length PhaZ ofBacillus
sp. NRRL B-14911 from a promoter present in pHY300PLK,
was introduced into B. subtilis cells, the transformants
pro-duced a clearing zone on an opaque LB agar plate supple-mented with 0.6% dPHB in the top agar (Fig. 1B). This is in
contrast to that observed in theB. subtilistransformants
har-boring the control vector pHY300PLK (Fig. 1B). These
obser-vations indicate that this putative enzyme ofBacillussp. NRRL
B-14911 has dPHB-degrading activity.
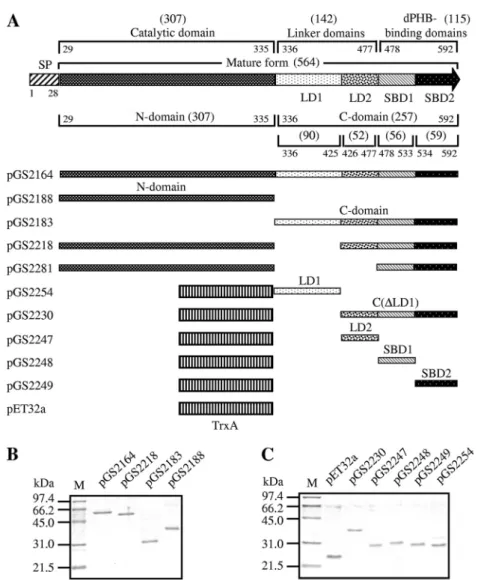
Characterization of the mature form of PhaZ ofBacillussp. NRRL B-14911.To examine the dPHB-hydrolyzing activity of
the mature form of Bacillus sp. NRRL B-14911 PhaZ, we
constructed plasmid pGS2164 (Fig. 2A) that could
overpro-duce the His-tagged mature form of PhaZ inE. coliand then
FIG. 1. (A) Growth ofBacillussp. NRRL B-14911 on a dPHB-containing M9 agar plate for 48 h. (B) Growth ofB. subtilisharboring the PhaZ(BspNRRL)-producing plasmid pGS2067 or the control plas-mid pHY300PLK on a dPHB-containing LB agar plate for 24 h.
on January 24, 2021 by guest
http://aem.asm.org/
purified this His-tagged protein by affinity chromatography on an Ni-NTA agarose column. A turbidimetric method that monitored the decrease in the turbidity of a suspension made from dPHB was used to examine the degradation of dPHB by the purified PhaZ. The result showed that this PhaZ could rapidly hydrolyze dPHB (Fig. 3). We also analyzed the
prod-ucts of hydrolysis of dPHB by this PhaZ with purified NAD⫹
-dependent (R)-3HB dehydrogenase as described in Materials
and Methods. The yield of 3HB monomers produced from dPHB degradation by this PhaZ corresponded to approxi-mately 99% of total 3HB equivalents present in dPHB. This
result suggests that the Bacillus sp. NRRL B-14911 PhaZ
works, at least in part, as an exotype enzyme.
The dPHB-degrading activity of PhaZ was strongly inhibited by the freshly prepared serine esterase inhibitor PMSF (96 and 99% inhibition at 1 and 10 mM, respectively), suggesting the presence of a nucleophilic serine residue at the active center of this PhaZ. The reducing agent DTT (dithiothreitol) also sig-nificantly inhibited the dPHB-degrading activity of PhaZ (50
FIG. 2. (A) Domain structures of theBacillussp. NRRL B-14911 PhaZ and plasmid constructs producing the mature form of PhaZ, its deletion derivatives, or various TrxA fusion proteins. The starting and ending positions of each domain within full-length PhaZ are numbered relative to the translational start site of PhaZ. The amino acid length of each domain is indicated in parentheses. SP, signal peptide. (B) SDS-12% PAGE of purified His-tagged proteins. (C) SDS-12% PAGE of purified His-tagged TrxA fusion proteins. M, molecular mass standards.
FIG. 3. Turbidimetric determination of the dPHB-degrading activ-ity of PHB depolymerase or its derivative. A decrease in the turbidactiv-ity of a reaction mixture (200l) containing dPHB (0.7 mg) and purified His-tagged protein (0.24M) was detected spectrophotometrically at 650 nm. Symbols: solid circles, the mature form of PhaZ; solid squares, the mature form of PhaZ with the deletion of the linker domain LD1; solid triangles, the N-domain of PhaZ; hollow circles, no protein.
on January 24, 2021 by guest
http://aem.asm.org/
and 75% inhibition at 1 and 10 mM, respectively), suggesting the requirement of a disulfide bond for the enzyme activity. PhaZ showed maximum activity toward dPHB at pH 9.0 and 70°C. The specific dPHB-degrading activity of PhaZ is approx-imately 23 U/mg of protein at pH 8.0 and 37°C and 220 U/mg of protein at pH 9.0 and 70°C. Its enzyme activity was not
increased by the addition of 1 mM CaCl2 to the reaction
mixture and was not inhibited by the presence of 10 mM EDTA, suggesting that the calcium ion is not required as a cofactor for the enzyme activity. This is in striking contrast to theB. megateriumN-18-25-9 PhaZ (37).
To analyze the dPHB-binding ability of PhaZ, the mature form of PhaZ was pretreated with 1 mM PMSF to inhibit the dPHB-degrading activity. After the incubation of PMSF-treated PhaZ with dPHB, the assay mixture was separated into the supernatant and the dPHB-containing pellet by centrifu-gation. The pellet was washed with the incubation buffer three times. The washed pellet and the supernatant from the assay mixture then were subjected to SDS-PAGE followed by West-ern blotting with anti-His tag antibody as the probe. The result showed that PhaZ was present in the pellet (Fig. 4A), suggest-ing that PhaZ could bind to dPHB. In a control experiment, PhaZ could not bind to chitin (Fig. 4B), indicating that the binding is specific.
Catalytic domain of PhaZ.The N-terminal catalytic domain
(from position 29 to 335; 307 amino acids) of Bacillus sp.
NRRL B-14911 PhaZ (Fig. 2A) showed homology with those
of the PhaZs of B. megaterium N-18-25-9 (level of identity,
61%),Bacillussp. SG-1 (ZP_021858765) (80%),B.
pseudofir-mus OF4 (YP_003425754) (61%), and B. megaterium QM
B1551 (YP_003560966) (61%) (Fig. 5). Interestingly, it also displayed homology with the intracellular PhaZ1 (323 amino
acids; GenBank accession no. FJ175152) of B. megaterium
ATCC 11561 (49%) (Fig. 5). PhaZ1 has been demonstrated to
be capable of degrading dPHB (5). The analysis of the amino acid sequence alignment of the catalytic domains of these
PhaZs suggests that Ser146, Asp222, and His295 of theBacillus
sp. NRRL B-14911 PhaZ constitute the putative catalytic triad, and that His63 is the putative oxyanion hole (Fig. 5). The sequential order of these residues indicates that the catalytic
domain of the Bacillus sp. NRRL B-14911 PhaZ belongs to
type 1. A pentapeptide sequence (G-L-S146-A-G) present in its
catalytic domain is similar to the lipase box motif (G-X-S-X-G) (where X represents any amino acid residue) (11) identified in many serine hydrolases.
The purified His-tagged catalytic domain (Fig. 2B) of the
Bacillus sp. NRRL B-14911 PhaZ had very weak dPHB-de-grading activity (Fig. 3), whereas its ability to hydrolyze the
synthetic substrate p-nitrophenylbutyrate (ca. 0.085 mol/
min/mg of protein) was comparable to that of the mature form
of PhaZ (ca. 0.088mol/min/mg of protein). Western blotting
showed that this catalytic domain had no dPHB-binding ability (Fig. 4C), whereas the remaining C domain could bind to dPHB (Fig. 4D). These observations are in agreement with the idea that the catalytic domain of this PhaZ possesses a full
structural requirement for the esterase activity againstp
-nitro-phenylbutyrate, but it still requires the presence of the remain-ing C domain for dPHB degradation.
Linker domains LD1 and LD2.We next attempted to define the linker domain of PhaZ. A BLAST search with the amino
acid sequence of theBacillus sp. NRRL B-14911 PhaZ from
position 301 to 592 as a probe against the nonredundant pro-tein databases of NCBI revealed an interesting result: a region
of theBacillussp. NRRL B-14911 PhaZ (roughly from position
336 to 425) is significantly similar in amino acid sequence to certain regions (ca. 90 amino acids long) of a small group of PhaZs and a large group of PHB depolymerase-unrelated pro-teins. These PHB depolymerase-unrelated proteins include the
FG-GAP repeat-containing protein of Arthrobacter aurescens
TC1 (YP_946834), the NHL repeat-containing protein of
Geo-bactersp. FRC-32 (YP_002537077), and the Ig domain protein
group 2 domain protein ofThermincolasp. JR (YP_003641592) (Fig.
6A). This putative linker domain of theBacillus sp. NRRL
B-14911 PhaZ, designated LD1 (Fig. 2A and 6A), neither possesses the sequence features of the three known types of linker domains of PhaZs (cadherin-like, fibronectin-like, and threonine-rich linker domains) nor shows significant amino acid sequence similarity with them. These features
indicate that the putative linker domain LD1 of theBacillus
sp. NRRL B-14911 PhaZ is a representative of a novel type of linker domain.
To test the dPHB-binding ability of the putative linker do-main LD1, we constructed a fusion protein of thioredoxin (TrxA) with LD1 as shown in the plasmid pGS2254 (Fig. 2A). It turned out that the purified His-tagged fusion protein TrxA-LD1 showed weak binding to dPHB in a reproducible manner (Fig. 4E). In a control experiment, the purified His-tagged TrxA protein itself was found to be unable to bind to dPHB (Fig. 4J). We also constructed a fusion protein of TrxA with the remaining C-terminal portion of PhaZ as shown in the plasmid pGS2230 (Fig. 2A). This purified His-tagged fusion protein exhibited a strong dPHB-binding ability (Fig. 4F).
Extracellular PHB depolymerases usually contain a dPHB-binding domain of 40 to 60 amino acids in length at their C
FIG. 4. Western blot analysis of results of dPHB- and chitin-bind-ing assays. dPHB or chitin (200g) was incubated with PMSF-treated PHB depolymerase (the mature form) or its derivative (1 to 3g) at 30°C for 15 min. The assay mixture then was separated into the su-pernatant and the dPHB- or chitin-containing pellet by centrifugation. The pellet was washed three times. Proteins in the washed pellet and in the supernatant from the assay mixture were analyzed by SDS-12% PAGE and Western blotting with anti-His tag polyclonal antibody as the probe. The experiments were performed at least twice. A repre-sentative experiment is shown. In, input His-tagged protein; T, total proteins before separation into fractions by centrifugation; S, super-natant; P, pellet.
on January 24, 2021 by guest
http://aem.asm.org/
termini (1, 16, 25, 35). Exceptionally, the extracellular PHB
depolymerases ofPseudomonas stutzeri(25) andMarinobacter
sp. strain NK-1 (19) contain two similar dPHB-binding do-mains at their C termini. To identify the possible dPHB-bind-ing domain of PhaZ, we divided the remaindPHB-bind-ing C domain of PhaZ into three parts (LD2, SBD1, and SBD2, of 52, 56, and 59 amino acids in length, respectively) (Fig. 2A) roughly on the basis of the amino acid alignment (Fig. 6B, C, and D). We then constructed fusion proteins of TrxA with each of them (Fig. 2A) to test their dPHB-binding abilities. It turned out that, like the fusion protein TrxA-LD1, the purified His-tagged fusion protein TrxA-LD2 also showed reproducibly weak binding to dPHB (Fig. 4G). There is no significant amino acid sequence similarity between LD1 and LD2. LD2 neither possesses the sequence features of the three known types of linker domains of PhaZs nor shows significant amino acid sequence similarity with them (Fig. 6B). However, homologs of LD2 are present in the
extracellular PhaZs ofB. megaterium,B. pseudofirmus, and
Bacil-lus sp. strain SG-1 (Fig. 6B). These features indicate that the
putative linker domain LD2 of theBacillussp. NRRL B-14911
PhaZ is also a representative of a novel type of linker domain. It is generally believed that the linker domain plays a
struc-tural role by providing suitable spacing between the catalytic domain and the substrate-binding domain for dPHB degrada-tion. It has been demonstrated previously that the deletion of
the fibronectin-like linker domain of theRalstonia pickettiiT1
PhaZ abolished its degrading ability, whereas its dPHB-binding ability still was maintained (24). This result indicates the importance of the presence of the linker domain for dPHB degradation by PhaZ. In this study, we found that the deletion of the putative linker domain LD1 from the mature form of PhaZ as shown in the plasmid pGS2218 (Fig. 2A) only partially reduced its dPHB-degrading ability (Fig. 3). Its dPHB-binding ability still was maintained (Fig. 4K). Therefore, we next at-tempted to delete LD1 and LD2 simultaneously from the mature form of PhaZ, as shown in the plasmid pGS2281 (Fig. 2A). Un-expectedly, the resulting PhaZ was insoluble and found only in inclusion bodies (data not shown). This prompted us to construct a fusion protein of the soluble protein TrxA with this deletion derivative. It was found that this fusion protein still was insoluble, even when the induction of protein production was carried out with 0.04 mM IPTG and at 16°C (data not shown). This problem hindered our analysis of the effect of the deletion of both LD1 and LD2 on the dPHB-degrading ability of this PhaZ.
FIG. 5. Alignment of amino acid sequences of the putative catalytic domains of extracellular PhaZs from severalBacillusspecies and the full-length intracellular PhaZ1 fromB. megateriumATCC 11561. The sequences of the putative catalytic domains of extracellular PhaZs from
Bacillussp. NRRL B-14911 [PhaZ(BspNRRL)],Bacillussp. SG-1 [PhaZ(BspSG)],B. pseudofirmusOF4 [PhaZ(Bps)],B. megateriumQM B1551 [PhaZ(BmeQM)], andB. megateriumN-18-25-9 [PhaZ(BmeN)], and the sequence of the full-length intracellular PhaZ1 fromB. megateriumATCC 11561 [PhaZ1(BmeATCC)] are shown. All sequences are numbered relative to the translational start sites of the proteins. The conserved histidine residues for the predicted oxyanion holes, the putative catalytic triads, and two conserved cysteine residues are shown in white on black. The lipase box-like sequences are shaded in gray. Identical residues are marked below with asterisks. GenBank accession numbers are the following: PhaZ(BspNRRL), ZP_01169502; PhaZ(BspSG), ZP_01858765; PhaZ(Bps), YP_003425754; PhaZ(BmeQM), YP_003560966; PhaZ(BmeN), BAF35850; and PhaZ1(BmeATCC), FJ175152.
on January 24, 2021 by guest
http://aem.asm.org/
dPHB-binding domains SBD1 and SBD2. We next tested the dPHB-binding abilities of purified His-tagged TrxA-SBD1 and SBD2. Like the mature form of PhaZ, both TrxA-SBD1 and TrxA-SBD2 showed strong binding to dPHB (Fig. 4H and I). There is no significant amino acid sequence simi-larity between SBD1 and SBD2 (Table 3). Neither SBD1 nor SBD2 shows significant amino acid sequence similarity to the two known types of dPHB-binding domains of PhaZs (Table 3). There is no cysteine residue in SBD1 and SBD2 (Fig. 6C and D). SBD1 and SBD2 do not contain the sequence motif HXXAGR (where X represents any amino acid residue) (Fig. 6C and D), which is conserved in the two known types of dPHB-binding domains (16). Nevertheless, homologs of SBD1
and SBD2 are present in the extracellular PhaZs ofB.
mega-terium,B. pseudofirmus, andBacillussp. SG-1 (Fig. 6C and D).
These features indicate that SBD1 and SBD2 of theBacillus
sp. NRRL B-14911 PhaZ are representatives of two novel and distinct types of dPHB-binding domains. The presence of two
dPHB-binding domains in the Bacillus sp. NRRL B-14911
PhaZ may provide an advantage for PhaZ to more easily gain access to dPHB in the environment.
DISCUSSION
The annotated size of the extracellular PhaZ ofBacillussp.
NRRL B-14911 (ZP_01169502) was 595 amino acids.
How-FIG. 6. (A) Alignment of amino acid sequences of the putative linker domains LD1 of extracellular PhaZs from severalBacillusspecies plus the homologous regions of several PHB-unrelated proteins. The sequences of the putative linker domains LD1 of the extracellular PhaZs from
Bacillussp. NRRL B-14911 [PhaZ(BspNRRL)],Bacillussp. SG-1 [PhaZ(BspSG)],B. pseudofirmusOF4 [PhaZ(Bps)],B. megateriumQM B1551 [PhaZ(BmeQM)], andB. megateriumN-18-25-9 [PhaZ(BmeN)] and the sequences of the homologous regions of Ig domain protein group 2 domain protein fromThermincolasp. JR [Ig(Tsp)], NHL repeat-containing protein fromGeobactersp. FRC-32 [NHL(Gsp)], and FG-GAP repeat-containing protein fromArthrobacter aurescensTC1 [FG-GAP(Aau)] are shown. (B) Alignment of amino acid sequences of the putative linker domains LD2 of extracellular PhaZs from severalBacillusspecies. (C) Alignment of amino acid sequences of the putative dPHB-binding domain SBD1 of extracellular PhaZs from severalBacillusspecies. (D) Alignment of amino acid sequences of the putative dPHB-binding domain SBD2 of extracellular PhaZs from severalBacillusspecies. All sequences are numbered relative to the translational start sites of the proteins. Amino acid residues that are fully conserved (B, C, and D) or conserved in at least six out of the eight sequences (A) are shown in white on black. GenBank accession numbers are the following: NHL(Gsp), YP_002537077; FG-GAP(Aau), YP_946834; and Ig(Tsp), YP_003641592. Other accession numbers are indicated in the legend to Fig. 5.
on January 24, 2021 by guest
http://aem.asm.org/
ever, there is no typical Shine-Dalgarno (SD) sequence (33) present with an appropriate spacing upstream from the pre-dicted translational start site ATG (5-CCGAAGAATGAAA
AGGAGGAATG-3⬘). Therefore, the translation of this PhaZ
is more likely to be initiated from a second ATG located 6 nucleotides downstream of the first one, because the second
ATG is preceded by a typical SD sequence (5⬘-AGGAGG-3⬘)
with a spacing of 10 nucleotides. Thus, this second ATG is predicted to work as the translational start site of PhaZ. If so, this PhaZ should comprise 592 amino acids, which is what was used throughout this paper.
Bacillussp. NRRL B-14911 is a marine bacterium that was isolated from seawater at a depth of 10 m. Tools for the genetic
manipulation of Bacillus sp. NRRL B-14911 have not been
successfully developed so far. Therefore, it is not feasible now
to disrupt thephaZ gene in the chromosome to examine its
influence on the ability ofBacillussp. NRRL B-14911 to form
a clearing zone on an opaque plate. It has been noted that the
PhaZs ofAlcaligenes faecalisAE122,Ralstonia pickettiiT1, and
Pseudomonas stutzeri, which contain type 1 catalytic domains, could degrade dPHB to produce a mixture of 3HB oligomers
and monomers, whereas those PhaZs from Acidovorax sp.,
Comamonas sp.,C. acidovorans,C. testosteroni, and Streptomy-ces exfoliates, which contain type 2 catalytic domains, could yield 3HB monomer as a main product (32). However, the
PhaZs fromBacillus sp. NRRL B-14911 and P. stutzeri (25)
contain type 1 catalytic domains, and almost all of the end products from the degradation of dPHB by these PhaZs are 3HB monomers. Therefore, there seems to be no absolute correlation between the types of the catalytic domains of PhaZs and the composition of end products of dPHB degra-dation by PhaZs.
It has been proposed that the fibronectin type III-like linker domains present in several bacterial water-insoluble polymer hydrolases (including extracellular PHB depolymerases) were acquired initially from an animal source and spread later by horizontal transfers between distantly related bacteria (2). It is
of interest that the putative linker domain LD1 of theBacillus
sp. NRRL B-14911 PhaZ is significantly similar in amino acid sequence to certain regions of a large group of PHB depoly-merase-unrelated proteins in the databases. It is unclear whether the regions that are present in many bacterial proteins
and homologous to the linker domain LD1 of theBacillussp.
NRRL B-14911 PhaZ also were acquired initially from an animal source and spread by horizontal transfers among dis-tantly related bacteria.
In this study, dPHB-binding assays revealed that each of the two linker domains, LD1 and LD2, showed weak binding to dPHB. This is somewhat different from the cadherin-like linker
domains of the extracellular PhaZs ofP. stutzeriand
Marino-bactersp. NK-1, which showed no binding to dPHB (19, 25). The significance of the presence of two distinct linker domains
in the Bacillus sp. NRRL B-14911 PhaZ and the biological
implication of the very weak binding of the linker domains LD1 and LD2 to dPHB are unclear.
It is worth mentioning that the calculated pI is 4.02 and 4.86
for the linker domains LD1 and LD2 of theBacillussp. NRRL
B-14911 PhaZ, respectively, and is 6.20 and 9.52 for its binding domains SBD1 and SBD2, respectively. Its dPHB-binding domains appear to be more basic than its linker do-mains. This phenomenon also is manifested in the extracellular PHB depolymerases of other types: the calculated pI is 3.73 for
the linker domain ofP. stutzeri PhaZ (ACG63776), 5.55 for
SDB2, and 8.95 for SDB1; 3.54 for the linker domain of
Marinobactersp. NK-1 PhaZ (BAC15574), 4.14 for SDB2, and
4.75 for SDB1; 4.78 for the linker domain ofR. pickettiiPhaZ
(BAA04986) and 5.98 for its SDB; 4.58 for the linker domain ofS. exfoliatesPhaZ (AAB02914) and 8.19 for its SDB; 8.76 for
the linker domain ofP. lemoigneiPhaZ2 (AAB17150) and 9.25
for its SDB; 5.08 for the linker domain ofP. lemoigneiPhaZ3
(AAB48166) and 9.36 for its SDB; 4.36 for the linker domain ofComamonassp. PhaZ (AAA87070) and 8.89 for its SDB;
and 6.08 for the linker domain of C. acidovorans PhaZ
TABLE 3. Amino acid sequence identities and similarities among the dPHB-binding domains of various PhaZs
dPHB-binding domain Length of dPHB-binding domain (amino acids)
Position of dPHB-binding domain (residue) % Identity/% similarity to PhaZ(BspNRRL) domain: SBD1 SBD2 PhaZ(BspNRRL) SBD1 56 478⫺533 100/100 14/20 PhaZ(BspSG) SBD1 56 478⫺533 58/77 2.9/5.8 PhaZ(BmeQM) SBD1 57 475⫺531 36/59 1.9/4.8 PhaZ(Bps) SBD1 57 482⫺538 40/62 2.9/5.7 PhaZ(BmeN) SBD1 57 475⫺531 36/59 1.9/4.8 PhaZ(BspNRRL) SBD2 59 534⫺592 14/20 100/100 PhaZ(BspSG) SBD2 60 534⫺593 12/19 62/73 PhaZ(BmeQM) SBD2 59 532⫺590 19/31 38/57 PhaZ(Bps) SBD2 59 539⫺597 13/24 37/60 PhaZ(BmeN) SBD2 59 532⫺590 15/31 40/58 PhaZ(Pst) SBDII 60 455⫺514 16/30 12/15 PhaZ(Pst) SBDI 50 527⫺576 11/19 13/18 PhaZ(Msp) SBDII 59 450⫺508 14/27 14/19 PhaZ(Msp) SBDI 50 529⫺578 12/15 10/19 PhaZ(Rpi) 53 440⫺492 1.9/2.9 16/28 PhaZ(Sex) 50 439⫺488 2.1/4.3 0.9/1.9 PhaZ2(Ple) 53 381⫺433 14/24 4.9/5.8 PhaZ3(Ple) 53 367⫺419 11/18 16/24
on January 24, 2021 by guest
http://aem.asm.org/
Downloaded from
(BAA19791) and 8.93 for its SDB. The biological significance of this phenomenon is not yet clear.
ACKNOWLEDGMENTS
This research was supported by grant NSC 97-2311-B-010-003-MY3 from the National Science Council and a grant, Aim for the Top University Plan, from the Ministry of Education of the Republic of China.
REFERENCES
1.Behrends, A., B. Klingbeil, and D. Jendrossek.1996. Poly(3-hydroxybu-tyrate) depolymerases bind to their substrate by a C-terminal located sub-strate binding site. FEMS Microbiol. Lett.143:191–194.
2.Bork, P., and R. F. Doolittle.1992. Proposed acquisition of an animal protein domain by bacteria. Proc. Natl. Acad. Sci. U. S. A.89:8990–8994. 3.Briese, B., B. Schmidt, and D. Jendrossek.1994.Pseudomonas lemoigneihas
five poly(hydroxyalkanoic acid) (PHA) depolymerase genes: a comparative study of bacterial and eukaryotic PHA depolymerases. J. Environ. Polym. Degrad.2:75–87.
4.Chang, S., and S. N. Cohen.1979. High frequency transformation ofBacillus subtilisprotoplasts by plasmid DNA. Mol. Gen. Genet.168:111–115. 5.Chen, H. J., S. C. Pan, and G. C. Shaw.2009. Identification and
character-ization of a novel intracellular poly(3-hydroxybutyrate) depolymerase from Bacillus megaterium. Appl. Environ. Microbiol.75:5290–5299.
6.Delafield, F. P., M. Doudoroff, N. J. Palleroni, C. J. Lusty, and R. Conto-poulos.1965. Decomposition of poly-beta-hydroxybutyrate by pseudomon-ads. J. Bacteriol.90:1455–1466.
7.Handrick, R., et al.2001. A new type of thermoalkalophilic hydrolase of Paucimonas lemoigneiwith high specificity for amorphous polyesters of short chain-length hydroxyalkanoic acids. J. Biol. Chem.276:36215–36224. 8.Handrick, R., S. Reinhardt, P. Kimmig, and D. Jendrossek. 2004. The
“intracellular” poly(3-hydroxybutyrate) (PHB) depolymerase of Rhodospiril-lum rubrumis a periplasm-located protein with specificity for native PHB and with structural similarity to extracellular PHB depolymerases. J. Bacte-riol.186:7243–7253.
9.Handrick, R., et al.2004. Unraveling the function of theRhodospirillum rubrumactivator of polyhydroxybutyrate (PHB) degradation: the activator is a PHB-granule-bound protein (phasin). J. Bacteriol.186:2466–2475. 10.Hisano, T., et al.2006. The crystal structure of polyhydroxybutyrate
depoly-merase fromPenicillium funiculosumprovides insights into the recognition and degradation of biopolyesters. J. Mol. Biol.356:993–1004.
11.Jaeger, K. E., et al.1994. Bacterial lipases. FEMS Microbiol. Rev.15:29–63. 12.Jendrossek, D.2002. Extracellular polyhydroxyalkanoate depolymerases: the key enzymes of PHA degradation, p. 41–83.InY. Doi and A. Steinbu¨chel (ed.), Biopolymers, vol. 3b: polyesters II. Wiley-VCH, Weinheim, Germany. 13.Jendrossek, D.2007. Peculiarities of PHA granules preparation and PHA depolymerase activity determination. Appl. Microbiol. Biotechnol.74:1186– 1196.
14.Jendrossek, D.2009. Polyhydroxyalkanoate granules are complex subcellular organelles (carbonosomes). J. Bacteriol.191:3195–3202.
15.Jendrossek, D., et al.1995. Biochemical and molecular characterization of thePseudomonas lemoigneipolyhydroxyalkanoate depolymerase system. J. Bacteriol.177:596–607.
16.Jendrossek, D., and R. Handrick.2002. Microbial degradation of polyhy-droxyalkanoates. Annu. Rev. Microbiol.56:403–432.
17.Jendrossek, D., I. Knoke, R. B. Habibian, A. Steinbu¨chel, and H. G. Schlegel.
1993. Degradation of poly(3-hydroxybutyrate), PHB, by bacteria and purifi-cation of a novel PHB depolymerase from Comamonassp. J. Environ. Polym. Degrad.1:53–63.
18.Kasuya, K., et al.1997. Biochemical and molecular characterization of the polyhydroxybutyrate depolymerase of Comamonas acidovorans YM1609, isolated from freshwater. Appl. Environ. Microbiol.63:4844–4852. 19.Kasuya, K., et al.2003. Cloning, expression and characterization of a
poly(3-hydroxybutyrate) depolymerase fromMarinobactersp. NK-1. Int. J. Biol. Macromol.33:221–226.
20.Klingbeil, B., R. M. Kroppenstedt, and D. Jendrossek.1996. Taxonomic identification of Streptomyces exfoliatus K10 and characterization of its poly(3-hydroxybutyrate) depolymerase gene. FEMS Microbiol. Lett.142:
215–221.
21.Kobayashi, T., M. Shiraki, T. Abe, A. Sugiyama, and T. Saito.2003.
Purifi-cation and properties of an intracellular 3-hydroxybutyrate-oligomer hydro-lase (PhaZ2) inRalstonia eutrophaH16 and its identification as a novel intracellular poly(3-hydroxybutyrate) depolymerase. J. Bacteriol.185:3485– 3490.
22.Kobayashi, T., K. Uchino, T. Abe, Y. Yamazaki, and T. Saito.2005. Novel intracellular 3-hydroxybutyrate-oligomer hydrolase inWautersia eutropha H16. J. Bacteriol.187:5129–5135.
23.Nielsen, H., J. Engelbrecht, S. Brunak, and G. von Heijne.1997. A neural network method for identification of prokaryotic and eukaryotic signal pep-tides and prediction of their cleavage sites. Int. J. Neural Syst.8:581–599. 24.Nojiri, M., and T. Saito.1997. Structure and function of
poly(3-hydroxybu-tyrate) depolymerase fromAlcaligenes faecalisT1. J. Bacteriol.179:6965– 6970.
25.Ohura, T., K. I. Kasuya, and Y. Doi.1999. Cloning and characterization of the polyhydroxybutyrate depolymerase gene ofPseudomonas stutzeriand analysis of the function of substrate-binding domains. Appl. Environ. Micro-biol.65:189–197.
26.Papageorgiou, A. C., S. Hermawan, C. B. Singh, and D. Jendrossek.2008. Structural basis of poly(3-hydroxybutyrate) hydrolysis by PhaZ7 depoly-merase fromPaucimonas lemoignei. J. Mol. Biol.382:1184–1194. 27.Pospiech, A., and B. Neumann.1995. A versatile quick-prep of genomic
DNA from gram-positive bacteria. Trends Genet.11:217–218.
28.Po¨tter, M., and A. Steinbuchel.2005. Poly(3-hydroxybutyrate) granule-asso-ciated proteins: impacts on poly(3-hydroxybutyrate) synthesis and degrada-tion. Biomacromolecules6:552–560.
29.Rehm, B. H. 2003. Polyester synthases: natural catalysts for plastics. Biochem. J.376:15–33.
30.Saegusa, H., M. Shiraki, and T. Saito.2002. Cloning of an intracellular D(-)-3-hydroxybutyrate-oligomer hydrolase gene from Ralstonia eutropha H16 and identification of the active site serine residue by site-directed mutagenesis. J. Biosci. Bioeng.94:106–112.
31.Sambrook, J., and D. Russell.2001. Molecular cloning: a laboratory manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 32.Scho¨ber, U., C. Thiel, and D. Jendrossek.2000. Poly(3-hydroxyvalerate)
depolymerase of Pseudomonas lemoignei. Appl. Environ. Microbiol. 66:
1385–1392.
33.Shine, J., and L. Dalgarno.1974. The 3⬘-terminal sequence ofEscherichia coli16S ribosomal RNA: complementarity to nonsense triplets and ribosome binding sites. Proc. Natl. Acad. Sci. U. S. A.71:1342–1346.
34.Shinomiya, M., T. Iwata, K. Kasuya, and Y. Doi.1997. Cloning of the gene for poly(3-hydroxybutyric acid) depolymerase ofComamonas testosteroni and functional analysis of its substrate-binding domain. FEMS Microbiol. Lett.154:89–94.
35.Sudesh, K., H. Abe, and Y. Doi.2000. Synthesis, structure and properties of polyhydroxyalkanoates: biological polyesters. Prog. Polym. Sci. 25:1503– 1555.
36.Sugiyama, A., T. Kobayashi, M. Shiraki, and T. Saito.2004. Roles of poly(3-hydroxybutyrate) depolymerase and 3HB-oligomer hydrolase in bacterial PHB metabolism. Curr. Microbiol.48:424–427.
37.Takaku, H., A. Kimoto, S. Kodaira, M. Nashimoto, and M. Takagi.2006. Isolation of a Gram-positive poly(3-hydroxybutyrate) (PHB)-degrading bac-terium from compost, and cloning and characterization of a gene encoding PHB depolymerase ofBacillus megaterium N-18-25-9. FEMS Microbiol. Lett.264:152–159.
38.Takanashi, M., and T. Saito.2006. Characterization of two 3-hydroxybu-tyrate dehydrogenases in poly(3-hydroxybu3-hydroxybu-tyrate)-degradable bacterium, Ralstonia pickettii T1. J. Biosci. Bioeng.101:501–507.
39.Thompson, J. D., D. G. Higgins, and T. J. Gibson.1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res.22:4673–4680.
40.Tokiwa, Y., and B. P. Calabia.2004. Degradation of microbial polyesters. Biotechnol. Lett.26:1181–1189.
41.Tseng, C. L., H. J. Chen, and G. C. Shaw.2006. Identification and charac-terization of theBacillus thuringiensis phaZgene, encoding new intracellular poly-3-hydroxybutyrate depolymerase. J. Bacteriol.188:7592–7599. 42.Williamson, D. H., J. Mellanby, and H. A. Krebs.1962. Enzymic
determi-nation of D(-)-beta-hydroxybutyric acid and acetoacetic acid in blood. Biochem. J.82:90–96.
43.Zhang, K., M. Shiraki, and T. Saito.1997. Purification of an extracellular D-(-)-3-hydroxybutyrate oligomer hydrolase fromPseudomonassp. strain A1 and cloning and sequencing of its gene. J. Bacteriol.179:72–77.