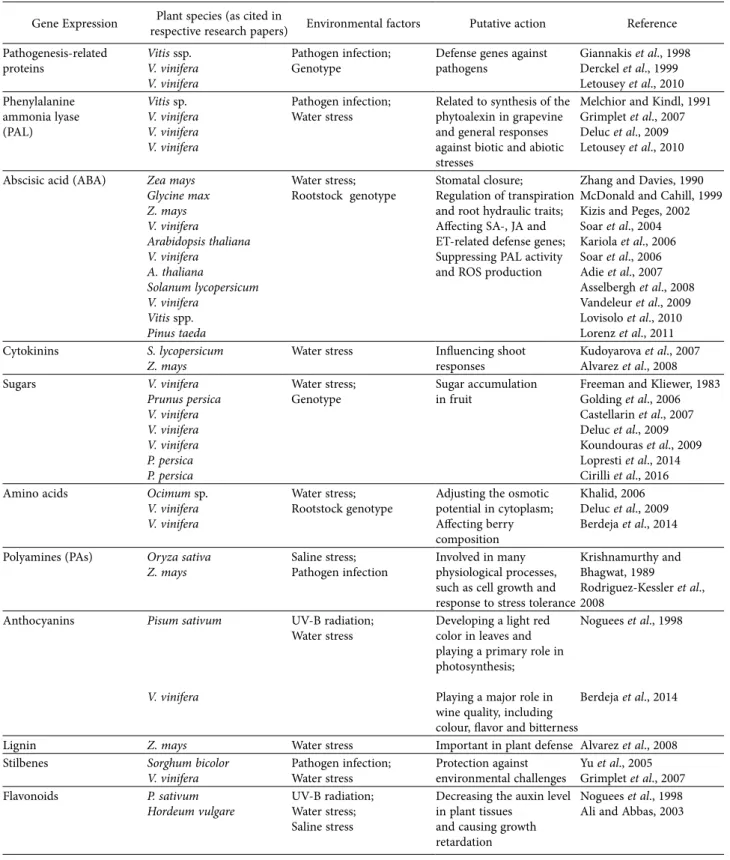
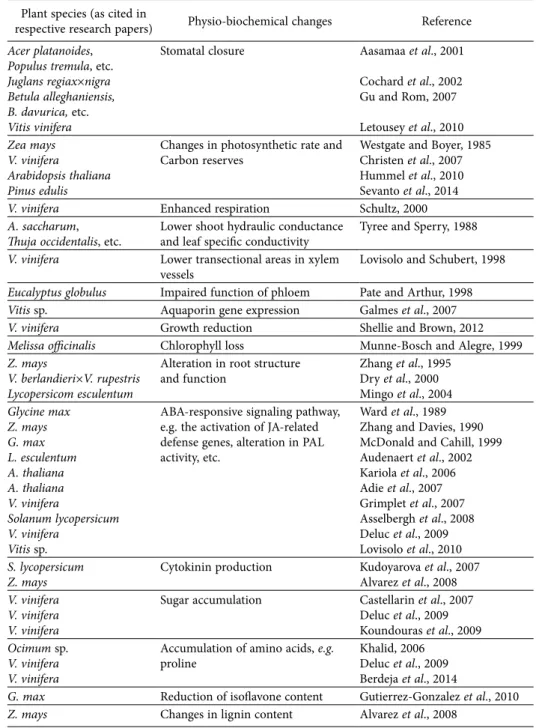
Grapevine, esca complex, and environment: the disease triangle
Full text
Figure
Related documents
been applied in the interdisciplinary socio-ecological systems literature and U.S. environmental law analysis, I suggest that climate change can be understood as an emergent
The ingestion of data is simply the process of introducing data from proprietary industry formats and/or standard data transfer protocols into an eDiscovery platform to allow
By comparing these models, we obtained estimated economical abatement costs to reduce emissions, social cost of carbon, and impact of climate change and global
11 With an endowment economy and separable preferences this indeterminacy has no effect on consumption but does affect utility (through real cash balances), so we classify this as
The mechanism that generates the negative relationship between monetary and real growth in the in fi nitely- lived, representative agent economy has an intuitive appeal: a higher
The Nash solution satis fi es all but Expansion Independence and Weak Expansion Independence, the egalitar- ian solution satis fi es all but E ffi ciency and Scale Invariance, and
The property tax was modeled as a triple of time-invariant tax rates, the 5rst applied to pre-development land value, the second to post-development (residual) site value (de5ned
