'STUD¥ :DF MULTIPLE CHARACTERISTICS OF CRASSULACEAN ACID METABOLISM IN A PLANT
D.O. 'MATHUR'IFAS,AG1lICULTURAL RESEARCH CeNTmt UNIVERSITY OF ;FLORIDA ApOPKA, FLORIDA 32703 USA
AND
H.M. VINES HORTICULTIJRE DEPAltTMl!NT UNIVERSITY OF GEoRGIA ATHENS, GEORGIA 30603 USA
Revised Aug. 1,1981·
SUMMARY
Environmental effects on six characteristics of Cr8Ssulacean acid metabolism (CAM) were studied at constant soil moisture potential. Kalanchoe tubiflora when &,£own'inenvironments inducing C AM had more stomata, a higher K/Ca ratio, "'lJSimi~ Jakd'nmte:C~;PCfgiven leaf area and more total growth than plants grown in an .nvironoientinduoing out of CAM plants. The plants grown out of CAM with lower ..stomatal.density ,utilized less water per gram dry weight.
INTRODUCTION
Many desert plants belong to·theCrassulaceaefamHy have distinct morphologi;. caLfcaturesthat aid in water conservation resulting in low transpiration rates and high
,water use'efficienoy (Joshi et 01., 1965). Most plants in this group 'open their-stomata
during dark and close during the day (Loftfield, 1921; Mathur, 1975, Nishida. 1963, Thompson-Cloudley, 1954). In these plants organic acids accmulate during the night and decrease during the day. This process is referred to as Crassulacean acid metabolism (CAM)andjsa,.;sumedtohave adaptive significance '{or conservation of water CO2 in certain types ofsuQculent plants.8ucculence·inaplanthas :beendefimd in terms of K/Caandin'CAMpJants varies in specific environments (James, 1958; Kunitllke and ,Saltman, 1955; La~tsch, 1974; Mathur, 1980; Mathur, et al., 1978; Mathur.and Vines, .1979; 'Ransom and Thomas, 1960: Walker, 1966).
Varied:envjronmental conditions can cause ~some CAM plants to be either in
D.O. MATHUR AND H.M. VINES
CAM (when plant shows diurnal fluctuations of as much as 25 meq/IOO g FW of titratable acidity) or out of CAM (when the concentrations of titratable acidity remain low with no, diurnal fluctuations). When the plants are grown in CAM versus out of I CAM environments, the rates of transpiration and dry matter accumulation should be different. Therefore, a compardtive study wa') made on stomatat numbers, photo synthetic rates, transpiration. KJCa ratio and dry matter accumulation of CAM plants maintained continuously under in CAM or out of CAM environments at a constant soil moisture tension.
MATERIALS AND METHODS
,
.
Individual stock plants of Kalanchoe tubiflora L. were selected and propagated to give populations of isogenetic plants. The plants were grown in pots in a (1:1) peat: perlite mixture in the growth chamber either in CAM (12 hours of 50 K lux incandes cent and fluorescent light at 30°C and 12 hours of dark at 15°C) or out of CAM (24 hours of 50 K lux incandescent and fluorescent light at 30°C) for 9 weeks. Relative humidity was 70-75%. Plants grown continuously for more than a year under these environments were used. Roots were washed and tolat fresh weight of 30 plants of equal height was determined. Ten of these plants were used to determine dry weight and 10 other plants were grown either in CAM or out of CAM environments. Growth was determined by increase in dry weight of plant. Since the measurement of the dry. 'weight showed .009% variance the resulting mean of 0.32 was used to calculate the dry weight of plants at the end of the experiment. The plants were transferred to plastic containers and grown as described earlier (Mathur etal.• 1978). Stomatal density and net photosynthesis was measured on top 7 to 8 centimeter marked same age leaves. The stomatal density, growth, transpiration ratio, K/Ca ratio and net photosynthesis with minor alteration in the shape of leaf chamber were determined according to procedures alreldy described (Mathur and Vines, 1976; Mathur et al.• 1978; Mathur ,and VineS, 1979; Mathur, 1980; Meinzer and R~ndel, 1973; Richard, 1951; Wolf et 01.
1969)..
RESULTS
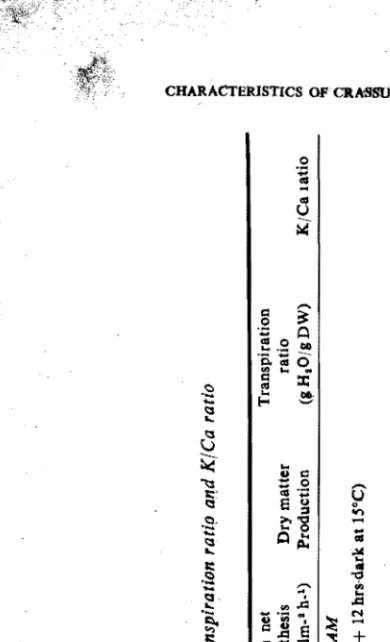
The data in Table I show ~tomatal numher, net photosynthesis, growth; transpi ration ratio and K/Ca ratio of Kalanchoe tubiflora when in CA M and the values Were
found to be 2450,6.80, 1.81, 75.03 and 1.86 respectively. When Kalanchoe tubiflorawas
•
grown out of CAM the stomatal uumber, net photosynthesis, dry ~atter production,transpiration ratio and K/Ca ratio was 1600, 0.25, 1;31, 35,.93 and 0.87. respectively. Water utilization in a short period shows that plants which have CAM type metabolism grow successfully in regions where environment'll conditions such as daily
(") ~ Table I. Stomatal number, net photosynthesis, growth, transpirl;"tion r(ltip and KfCa ratio
'lG > ~ ~
Plant
Soil Moisture. Potential Stomatal number (dm-I )
Maximum
net
Photosynthesis (mc
CO.
dm-I h_i)
Dry matter Production Transpiration ratio (JHIO/gDW) K/Ca latio
.... r:n >-l (5 r:n ~
IN
CAM
~
Kaltlllcftoe tubilJora
L.
Field capacity
(12 hrs light, SOK lux at 30°C
+
12 hrs·dark at IS°C) 24S0±8 6.8±0.20 1.IJI ± 0.14 7U3±4.1? 1.86 ± 0.20I
I'"..
~
Kalqnch(}e tubi/lora
L.
:field capacity
1600± S OUT OF CAM (24 hrs licht, SOK lux at JOoC) O.2S±0.IS 1.3l ±O.1S IS.5I! ± 3.S1 0.87± O.IS
:IIi :ill
;;
5
Pl~nta were grown at constant soil moisture tension for 9 weeks at light intcIlSity of 50 K lux with 12 hours of light at
~
30"C and 12 hours of dark at 15°C, in CAM and 24 hours of light at 30°C, out of CAM. Relative pumidity 70.75%.~
....
r:na::
/
V> ...
~:..
lation during 12 hours of dark period caused differences in transpiration rates and dry matter accumulation when in CAM versus out of CAM (Table I).
Similar maximum dark CO2 assimilation has been reported in other CAM plants (Bannet and Clark~ 1931; Gregory et al., 1954; Mathur et al., 1978; Mathur, 1980; Zabka
et
al., 1958; Zabka and Chaturvedi 1975; Zabka and McMahen. 1964), In '::Chrysanthemum plants, photoperiod alone has been shown to regulate stomatal behaviour (Schwabe, 1952). The night time opening c# stomates in the present study on Kalanchoe tubiflora is attributed to either the formation of organic acids which caused lowering of the pH and subsequently increased the osmotic value or due to swelling of ,amphoteric colloids as proposed by Nishida (1963). The decarboxylation of organic acids in the light then reversed the process controlling\
stomatal size and closure of stomates during the daytime. This reverse stomatal response was found when'Kalanchoe tubijlQra was growl) out of CAM thus confirming that major COli assimilation in these test plants occurred in the dark. Plants which do not grow well in continuous light for idefinite period lack capacity to adapt under adverse conditions in a short period as do CAM plants.
The growth at the end of 9 w~ks was' also hi$her when Kalanchoe tubiflora was in CAM, water utilization was more than
two
times higher when in CAM versus out of CAM. This is attributed to higher number of stomata i.e. 2450 per dmll, higher CO2 assimilation during 12 hours of dark at 15D
C and high endogenous CO. fixation during 12 hours of light at 30D
C. Plants under out
of
CAM environment had less growth and lower w~ter utilization (Table I). This is attributed to lower number of stomata ie. 1600 p~r dmZ, lower CO2 assimilation during 24 hours of light at 30°C and high endogenous COt fixation for 24 hours at 30DC. The out or CAM environment can be similar to situatio,n where' succulent plants did not show COli assimilation under 24 hours of light at 30°C (Nishida, 1963; Mathur,ei aI., 1978; Ransom and Thomas, 1960). Short-term environmental effects on laC shift in the leaves of CAM plant have
already been reported (Mathur aud Vines, 1978). Therefore, in CAM plant growth is t limited by photoperiod, ,thermoperiod, CO. assimilation and endogenous COt fixation
as compared to transpiration in a specific environment.
, D.O. MATHUR AND H.M. VINES
DISCUSSION
In Kalanchoe tubijlora, change in stomatal density and maximum COli assimi
•
CHARACTERISl'ICS OF CRASSULACEAN . ACID METABOLISM $3
t.SOfJames;J958; Mathut eial.,'1918: Mathur and Vines, 1979). ',The observed K/Ca
ratio of Kalanchoe tubiflora is in agreement. It would be appropriate to mention that
succuleai:le-may be associated witli • CAM in some plants but succulence does riot
necessarily assure the presence of CAM.
Therefore, on the basis of the present results, it is concluded that in CAM and
'Don C:::AM'plantsstomata play an important-role in COil assimilation:. transpiration
and
. dry matter production in CAM plants grown in a specific environment. SinCe crassul
'acean acid "metabolism' changes' iIi: response to a specific environment, 'studies of
changes of more than four of these characteristics are desirable.
". -"
REFERENCES
Bannet-Clark, T. A. (1933). The role of organic acids in plant metabolism. New phytol., 32: 37-n, 128-61, 197-230.
Bunnil\l. E. (1960). Circadian rhythms and the time ~surement in photoperiodism. Cold Spring H!lr\>OrSym. QlU1nt•. Biol., 25: 249-56.
• - . • <" • -~ • .
GteBQry, F. G., Spear, J. and Thiman, K. V. (1954). The interrelationship between CO, metabolism
, '.'
; .. an4
photOpeti~ism hl Kalanchoe plants. plant Physiol., 29: ~20-29.Ii. " .
James. W. O. (1958). SuccuieJlt Plants .. Endeavour.•. ,
n:
90-5•.Joshi, M. C., Boyer, J. S. and Kramer, P. J. (1956). Growth, carbon dioxide exchange, transpiration and transpiration ratio of pineapple. Bot. Gaz., 126: 174-179.
Kunitake, G. M. and SaItman, P. (1958). Dark fixation of CO. by succulent leaves. Conservation of dark fixed COs under d;urnal conditions. Plant. Physiol., 33; 40-403.
Laetsch, W. M. (1974). lhe C, syndrome; A structural analysis. Ann. Rev. Plant Physiol., 25: 27-52. Loftfield, J. V. G. (1921). The behavior of stomata. Carnegie Inst., Wash. Publ. No. 314.'
Mathur, D. D. (1975). Environmentally induced changes in Crassulacean Acid Metabolism. Doctoral Dissertation, University of Oeorg:a, Athens.
- - - 8 l I d Vines., H. M. (1976). Photoperiod and Thermoperiod Effects on Crassulacean Acid Metabolism. Ind. J. Plant Physiol., 19 (I) : 47-52.
_______• (1978). Environmental effects on 3laC shift in the leaves of Sedum Rubrotinctum.
Comm. Soil and Plant Analysis, 9 (9), 843-850.
_ - - - - , Hendershott, C. H. and Vines, H. M. (1978). Efficiency of Water Utilization in Crassu !acean Acid Metabolism Plants When IN CAM versus OUT OF CAM. Ind. J. Plant
Physiol., 21 (I) : 7-11.
:...:..-___- - - . Natarella, N. J. and Vines, H. M. (1978). Elemental Analyses of Crassulacean Acid .. Metabolism Plant Tissue. Comm. Soil and Plant Analysis, 9 (2) : 121-39.
______, and Vines, H. M. (1979). Environmental Effects on Ihe Element!ll Leaf Composition of Crassulacean Acid Metabolism Plants •.. J. Plant Nutrition, 1 (4) ; 407-16.
Nishida. ~
lism. Physiol.
Ransom, S. L. and Thomas, M. (1960). 11 : 81-100.
R.idlard, L. A. (1951). A
51: 377-86. Schwabe,. W. W. (1952).
169 : 1053-55.
Tbompson-Cloudley. (1954).
Walker, D. A. (1966).
D.:D.. MATlIUR. AND H.M. VINES
, MciDzar, F. C. awl Riuadel, P; W. (1973). Crassulaceanaeid metabolism and
.\ft_
use eliciency,iaEche'Hria pumila •. PlwlosYlllhuica, 7 : 258-64.
(1969). Studies.on stomatal movement ofcrassulaeeanplantl!. in. relanoll to acUi metabo
Plafft., 16: 281-98.
Crassulacean acid metabolism. Ann. Rev •. Plant Phys;ol.,
pressure membrane extraction apparatus for soil solutions. &Iii Sd.,
Effects of photoperiodic treatment on stomatal movement. Natllre,
Biology of the desert. Heffner and Sons, New York.
Carboxylation in plants. Endeavour, 15: 21-26.
Wolf, D. D., Pearce, R B., Carl&ow, G. E. and Lee. D. R.. (1969).
..
Measuring pbotosynthesis of attached~ with air sealed chambers. Crop Sci., I) : 26-27 Zebka,G. G., Gregory, F. G. and Edelman, 1. 0_). Dark fixation of carbon dioxide in Xalanchoe
in relation to photoperiodism. Nature 183; 1375-77.
- - - a n d McMahen. E. (1964). R.elationship among COt dark fixation succulence, flowering and organic acid formation in Xalanchae blossfeldiana variety Tom Thumb•. Can. J Bot.,
42: 44'7-52.
- - - a n d Chaturvedi, S. N. (1975). Water conservation in l\.alanchoe biossfeldiana in relation