0095-1137/10/$12.00
doi:10.1128/JCM.01620-09
Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Identification of Clinically Important Anaerobic Bacteria
by an Oligonucleotide Array
䌤
†
Yu Tzu Lin,
1Mario Vaneechoutte,
2Ay Huey Huang,
3Lee Jene Teng,
4Hung-Mo Chen,
3Shu-Li Su,
1and Tsung Chain Chang
1*
Department of Medical Laboratory Science and Biotechnology, College of Medicine, National Cheng Kung University, Tainan,
Taiwan
1; Laboratory Bacteriology Research, Department of Clinical Chemistry, Microbiology, and Immunology, University of
Ghent, Ghent, Belgium
2; Division of Clinical Microbiology, Department of Pathology, National Cheng Kung University Hospital,
Tainan, Taiwan
3; and School of Medical Technology, National Taiwan University College of Medicine, Taipei, Taiwan
4Received 20 August 2009/Returned for modification 11 October 2009/Accepted 15 January 2010
Anaerobic bacteria can cause a wide variety of infections, and some of these infections can be serious.
Conventional identification methods based on biochemical tests are often lengthy and can produce inconclusive
results. An oligonucleotide array based on the 16S-23S rRNA intergenic spacer (ITS) sequences was developed
to identify 28 species of anaerobic bacteria and
Veillonella. The method consisted of PCR amplification of the
ITS regions with universal primers, followed by hybridization of the digoxigenin-labeled PCR products to a
panel of 35 oligonucleotide probes (17- to 30-mers) immobilized on a nylon membrane. The performance of the
array was determined by testing 310 target strains (strains which we aimed to identify), including 122 reference
strains and 188 clinical isolates. In addition, 98 nontarget strains were used for specificity testing. The
sensitivity and the specificity of the array for the identification of pure cultures were 99.7 and 97.1%,
respectively. The array was further assessed for its ability to detect anaerobic bacteria in 49 clinical specimens.
Two species (Finegoldia magna
and
Bacteroides vulgatus) were detected in two specimens by the array, and the
results were in accordance with those obtained by culture. The whole procedure of array hybridization took
about 8 h, starting with the isolated colonies. The array can be used as an accurate alternative to conventional
methods for the identification of clinically important anaerobes.
Anaerobic bacteria are important human pathogens, and
infections caused by these bacteria can be serious and
life-threatening (6). A recent report from the Mayo Clinic
(Roch-ester, MN) revealed an overall increase in the incidence of
anaerobic bacteremias of 74% from 2001 to 2004 compared to
that from 1993 to 1996 (20), although the same trend was not
found in community hospitals or in an European countries (2,
11). The commonly isolated anaerobic bacteria are the
mem-bers of the
Bacteroides fragilis
group and
Peptostreptococcus
,
Clostridium
, and
Fusobacterium
species (3, 6, 20).
Most clinical laboratories use differential biochemical tests for
the identification of anaerobic microorganisms (35). However,
Simmon et al. (31) found that 24% of the isolates of anaerobic
bacteria recovered from blood cultures were misidentified and
that 10% isolates were not identified to the species level by
phe-notypic characteristics. A rapid commercial kit, the Rapid ID 32A
kit (bioMe
´rieux, Marcy l’Etoile, France), was evaluated for its
ability to identify strains in the
Bacteroides fragilis
group. The
results showed that only 78.4% of the strains were correctly
iden-tified to the species level without supplemental tests (15). The
success of the Rapid ID 32A system for species identification
varied with different taxa (10), and a low identification rate (50%)
was observed for fusobacteria (16).
Veillonella
isolates are
rela-tively easily identified to the genus level, but the differentiation of
Veillonella
isolates at the species level remains difficult and
incon-clusive due to the lack of discriminatory tests (14). In recent years,
increasing antimicrobial resistance for some anaerobic bacteria
(1, 13, 33) were noted, especially for species in the
B. fragilis
group
(40). The rapid identification of anaerobic bacteria and the
ad-ministration of appropriate antimicrobials play crucial roles in
preventing mortality and morbidity in patients (6).
Molecular methods have emerged as accurate alternatives for
the identification of anaerobic bacteria (21, 22, 34, 36).
Approx-imately 9% isolates of bacteremic anaerobes could not be
iden-tified to the species level by 16S rRNA gene sequencing, although
all isolates were correctly assigned to the genus level (31). Other
molecular identification methods targeting the rRNA operon
in-clude PCR (32), real-time PCR (26), PCR-restriction fragment
length polymorphism analysis (39), and matrix-assisted laser
de-sorption ionization–time-of-flight mass spectrometry (37).
The intergenic spacer (ITS) region separating the 16S and 23S
rRNA genes has been suggested to be a good candidate for use
for the identification of aerobic and anaerobic bacteria (8, 19, 42).
Moreover, the DNA array technology has been applied to the
identification of a variety of microorganisms (12, 17, 41). The aim
of the study described here was to develop an oligonucleotide
array based on the ITS sequences to identify 28 clinically
impor-tant species of anaerobes and
Veillonella
.
MATERIALS AND METHODS
Bacterial strains.A collection of 310 target strains (122 reference strains and 188 clinical isolates) representing 28 species of anaerobic bacteria andVeillonella
spp. was analyzed (Table 1). Reference strains were obtained from the American
* Corresponding author. Mailing address: Department of Medical
Laboratory Science and Biotechnology, College of Medicine, National
Cheng Kung University, 1 University Road, Tainan 701, Taiwan,
Re-public of China. Phone: 886-6-2353535, ext. 5790. Fax: 886-6-2363956.
E-mail: [email protected].
† Supplemental material for this article may be found at http://jcm
.asm.org/.
䌤
Published ahead of print on 3 February 2010.
1283
on May 16, 2020 by guest
http://jcm.asm.org/
TABLE 1. Anaerobic bacteria used for identification by the array
Species Reference strain(s) Clinical isolate(s)
Total no. of strains
Anaerococcus prevotii CCUG 41932T
, CCUG 44020A, CCUG 44289 A776-1, C270, C400-3 6
Anaerococcus tetradius CCUG 44198, CCUG 46590T
2
Bacteroides fragilis ATCC 29762, CCUG 4856T
, CCUG 25931 6121N, L778-1, 5830 N2, 6552N, M393, M631, M850, N146, N227-2, M926-1, N925, Q61750, 2066N, W507, 464, 562-1, Z118, Z353-1, 846-1, 646, Z875, 25, 81-2, 828, A404, 7071N, I626-4, 273N, K445, 304, J304, 1193N, L553-1, 3134N, 4073N, 911-1, 911-2, P771-3, 341-3, R664, 133, 247-1
45
Bacteroides ovatus ATCC 8483T, CCUG 35192, CCUG 48900,
CCUG 700292
4
Bacteroides thetaiotaomicron ATCC 29741, CCUG 10774T, CCUG 34419,
CCUG 38386A, CCUG 38792
Z362-2, B478, E51-1, J951, J416, L304, L100-1 12
Bacteroides uniformis ATCC 8492T
, CCUG 35501, CCUG 39989 6895N3 4
Bacteroides ureolyticus CCUG 7319T
, CCUG 44020B, CCUG 45874, CCUG 48545, CCUG 49390, CCUG 52055
N951-1, U194 8
Bacteroides vulgatus ATCC 8482T, CCUG 12546, CCUG 36807 5915 N1, 6895N1, P746, P771-1 7
Bilophila wadsworthia CCUG 32349T, CCUG 43589, CCUG 43686,
CCUG 45035
4
Clostridium bifermentans CCUG 9279, CCUG 32113, CCUG 33082, CCUG 35297, CCUG 35556A, CCUG 36626T
, CCUG 43592
1710N1, J46 9
Clostridium difficile CCUG 4938T
, CCUG 19126, CCUG 37766, CCUG 37779, CCUG 37780
5278 N3, A193, B442, C350-1, D686, I191, 83, U418, #626, #4346 15
Clostridium perfringens CCUG 1795T, CCUG 18370, CCUG 33957,
CCUG 35750, CCUG 44727
L717-1, L277, M194-2, M631-3, 1710 N2, I789, I860, 6456, #1428, #2887, #8104, #8603
17
Clostridium septicum CCUG 4855T, CCUG 30432, CCUG 35171,
CCUG 38619, CCUG 39580, CCUG 39581, CCUG 49629, CCUG 50972
N140 9
Finegoldia magna CCUG 17636T
, CCUG 12548, CCUG 4947 L943-3, O99, O55-2, 335-4, 869, Z351-1, A653, D752-3, D794-1, J737-1, L558-2, R490-2, 347, U440
17
Fusobacterium mortiferum CCUG 14475T
2377N 2
Fusobacterium necrophorum
subsp. necrophorum
BCRC 10679T, JCM 3718 2
Fusobacterium nucleatum CCUG 45873 23, 365, 507, Q338, 5084N2 6
Fusobacterium nucleatum
subsp. animalis
JCM 11025T
1
Fusobacterium nucleatum
subsp. fusiforme
JCM 11024T
1
Fusobacterium nucleatum
subsp.nucleatum
CCUG 32989T 1
Fusobacterium nucleatum
subsp.polymorphum
CCUG 9126T, CCUG 37995, CCUG 51781 3
Fusobacterium nucleatum
subsp. vincentii
JCM 11023T
1
Fusobacterium varium CCUG 4858T
, CCUG 35114, CCUG 49763 166 4
Peptoniphilus asaccharolyticus CCUG 9988T, CCUG 12549, CCUG 42643 Z354-2, 101, A658, B619-2, B678-2, C302-2, D752-2, G14-1, J578,
3480N, 247-2
14
Peptostreptococcus anaerobius CCUG 7835T, CCUG 37992, CCUG 49327,
CCUG 46594
562-2, Z354-1, B226-3, K789-2, 374, S973-2 10
Parvimonas micra
(Peptostreptococcus micros)
CCUG 46357T
, CCUG 48626 A, CCUG 51889
K775-1, K709-2, K520-3, N185, N366, N458-2, N785-2, 1840N, 530-2, 843, 848-2, 481-1, A735, B619-1, G747, 8462N, I503, iI404, 8594 M, 38N, J997, J764-2, 2376, 2415N, 3760N, U201, U193
30
Porphyromonas asaccharolytica
ATCC 27067, CCUG 7834T
2
Porphyromonas gingivalis CCUG 14449, CCUG 25211, CCUG 25226, CCUG 25893T, CCUG 27724
5
Prevotella intermedia BCRC 14416T, CCUG 27725, CCUG 27726 895-2, E325, I626-1, K497, K86-1 8
Prevotella melaninogenica CCUG 4944BT
, CCUG 51439, JCM 6321 3
Propionibacterium acnes BCRC 16144, BCRC 16145, CCUG 1794T
, CCUG 4945, CCUG 6369
4563N, 5037N, 5232N, 5244, 6105N, 6414, 7231, M848, 8201N, 8216N, O340, O62, 1835N, Z987, A975, G279, 6998N, 8757N, J191, 1311N, 1172N, L340, L440, 2132N
29
Propionibacterium granulosum
CCUG 32987T, CCUG 14831, CCUG 43704,
CCUG 52208
N467-2 5
Propionibacterium propionicus
ATCC 29325, ATCC 29326, CCUG 4939T,
LMG 16717
4
Veillonellaspp. L515-1, 369, 729, B462-3, 3237N, J825, 749, S546-2, S973-1 9
Veillonella atypica ATCC 17744T
1
Veillonella dentocariosi CCUG 54361, CCUG 54362T 2
Veillonella dispar ATCC 17748T 1
Veillonella montpellierensis CCUG 48299T
1
Veillonella parvula CCUG 35361, CCUG 44569, CCUG 45896,
CCUG 47017, CCUG 48555, CCUG 5123T 6
Total 122 188 310
on May 16, 2020 by guest
http://jcm.asm.org/
Type Culture Collection (ATCC; Manassas, VA); the Belgian Coordinated Col-lections (BCCM/LMG; Ghent, Belgium); the Bioresources Collection and Re-search Center (BCRC; Hsinchu, Taiwan); the Culture Collection, University of Go¨teborg (CCUG; Go¨teborg, Sweden); and the Japan Collection of Microor-ganisms, Riken BioResource Center (JCM; Saitama, Japan). Clinical isolates, identified by use of the Rapid ID 32A system, were obtained from the National Cheng Kung University Hospital (Tainan, Taiwan) and the National Taiwan University Hospital (Taipei, Taiwan). In addition, 98 nontarget strains (51 spe-cies) were used for specificity testing of the oligonucleotide array (see Table S1 in the supplemental material). All anaerobic bacteria were cultured on CDC anaerobe 5% sheep blood agar (BBL, Becton Dickinson Microbiology Systems, Cockeysville, MD) and incubated in an anaerobic chamber at 35°C, while aerobic and facultative anaerobic bacteria were cultured on blood agar or chocolate agar plates and incubated in ambient air at 35°C.
DNA preparation and ITS sequencing.The boiling method was used to extract DNA from the bacteria (24). The ITS sequences of some anaerobes were de-termined in this study and were submitted to GenBank (Table 2). The bacterium-specific universal primers 2F TTGTACACACCGCCCGTC-3⬘) and 10R (5⬘-TTCGCCTTTCCCTCACGGTA-3⬘) were used to amplify the ITS regions, as described previously (41). The TOPO TA cloning kit (Invitrogen, Carlsbad, CA)
was used for cloning of the ITS region for species that possessed multiple ITS fragments with different lengths and sequences, according to the manufacturer’s instructions. The ITS fragments of positive clones were amplified by PCR and sequenced (41).
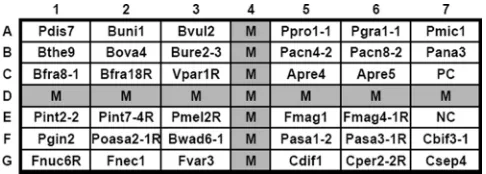
[image:3.585.46.540.82.458.2]Design of oligonucleotide probes and array fabrication.Thirty-five oligonu-cleotide probes (18- to 30-mers) (Table 2) were designed to identify the anaer-obic bacteria listed in Table 1. These probes included 33 species- and group-specific probes and 2 positive control probes (designed from the 3⬘ends of the 16S rRNA genes). Each probe except the positive control was spotted on the array as a single dot; the positive control dot contained a mixture of two probes at equal concentra-tions (Table 2). Ten or 15 additional bases of thymine were added to the 3⬘end of each probe to increase the hybridization signal (7). An irrelevant probe (code M) (5⬘-digoxigenin-GCATATCAATAAGCGGAGGA-3⬘) labeled with digoxigenin at the 5⬘end was used as a position marker on the array (Fig. 1). The oligonucleotide probes were diluted with a tracking dye, drawn into wells of a 96-well microtiter plate, and spotted onto positively charged nylon membranes (Roche, Mannheim, Germany), as described previously (41). The arrays (0.7 by 0.7 cm, 7 by 7 dots) were fabricated with an automatic arrayer (model SR-A300; EZlife Technology Co., Taipei, Taiwan) by use of a solid pin (diameter, 400m). The layout of the different probes on the array is shown in Fig. 1.
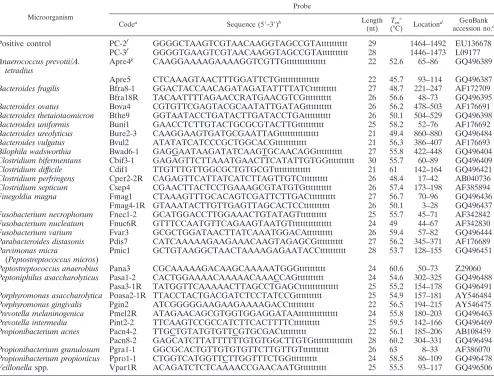
TABLE 2. Oligonucleotide probes used to identify anaerobic bacteria
Microorganism
Probe
Codea Sequence (5⬘–3⬘)b Length
(nt)
Tmc
(°C) Location
d GenBank accession no.e
Positive control
PC-2
fGGGGCTAAGTCGTAACAAGGTAGCCGTAtttttttttt
29
1464–1492
EU136678
PC-3
fGGGGTGAAGTCGTAACAAGGTAGCCGTAtttttttttt
28
1446–1473
L09177
Anaerococcus prevotii/A.
tetradius
Apre4
gCAAGGAAAAGAAAAGGTCGTTGttttttttttttttt
22
52.6
65–86
GQ496389
Apre5
CTCAAAGTAACTTTGGATTCTGttttttttttttttt
22
45.7
93–114
GQ496387
Bacteroides fragilis
Bfra8-1
GGACTACCAACAGATAGATATTTTATCtttttttttt
27
48.7
221–247
AF172709
Bfra18R
TACAATTTTAGAACCRATGAACGTCGtttttttttt
26
56.6
48–73
GQ496395
Bacteroides ovatus
Bova4
CGTGTTCGAGTACGCAATATTGATAGtttttttttt
26
56.2
478–503
AF176691
Bacteroides thetaiotaomicron
Bthe9
GGTAATACCTGATACTTGATACCTGAtttttttttt
26
50.1
504–529
GQ496398
Bacteroides uniformis
Buni1
GAACCTCTTGTACTGCGCGTACTTGtttttttttt
25
58.2
52–76
AF176692
Bacteroides ureolyticus
Bure2-3
CAAGGAAGTGATGCGAATTAGttttttttttttttt
21
49.4
860–880
GQ496484
Bacteroides vulgatus
Bvul2
ATATATCATCCCGCTGGCACGtttttttttttt
21
56.3
386–407
AF176693
Bilophila wadsworthia
Bwad6-1
GAGGAATAAGATATCAAGTGCAACAGGtttttttttt
27
55.8
422–448
GQ496404
Clostridium bifermentans
Cbif3-1
GAGAGTTCTTAAATGAACTTCATATTGTGGtttttttttt
30
55.7
60–89
GQ496409
Clostridium difficile
Cdif1
TTGTTTGTTGGCGCTGTGCGTtttttttttttttt
21
61
142–164
GQ496421
Clostridium perfringens
Cper2-2R
CAGAGTTCATTATCATCTTAGTTGTCtttttttttt
26
48.4
17–42
AB040736
Clostridium septicum
Csep4
CGAACTTACTCCTGAAAGCGTATGTGtttttttttt
26
57.4
173–198
AF385894
Finegoldia magna
Fmag1
CTAAAGTTTGCACAGTCGATTCTTGACtttttttttt
27
56.7
70–96
GQ496436
Fmag4-1R GTAAATACTTGTTGAGTTAGCACTCCtttttttttt
26
50.1
3–28
GQ496437
Fusobacterium necrophorum
Fnec1-2
GCATGGACCTTGGAAACTGTATAGTtttttttttt
25
55.7
45–71
AF342842
Fusobacterium nucleatum
Fnuc6R
GTTTCCAATGTTCAGAAGTAATGTttttttttttttttt
24
49
44–67
AF342830
Fusobacterium varium
Fvar3
GCGCTGGATAACTTATCAAATGGACAtttttttttt
26
59.4
57–82
GQ496444
Parabacteroides distasonis
Pdis7
CATCAAAAAGAAGAAACAAGTAGAGCGtttttttttt
27
56.2
345–371
AF176689
Parvimonas micra
(
Peptostreptococcus micros
)
Pmic1
GCTGTAAGGCTAACTAAAAGAGAATACCtttttttttt
28
53.7
128–155
GQ496451
Peptostreptococcus anaerobius
Pana3
CGCAAAAAGACAAGCAAAAATGGGtttttttttt
24
60.6
50–73
Z29060
Peptoniphilus asaccharolyticus
Pasa1-2
CACTGGAAAACAAAAACAAACCAGtttttttttt
24
54.6
302–325
GQ496488
Pasa3-1R
TATGGTTCAAAAACTTAGCCTGAGCttttttttttttttt
25
55.2
154–178
GQ496491
Porphyromonas asaccharolytica
Poasa2-1R TTACCTACTGACGATCTCCTATCCGtttttttttt
25
54.9
157–181
AY546484
Porphyromonas gingivalis
Pgin2
ATCGGGGGAAGAAGAAAAGACCtttttttttt
22
56.5
194–215
AY546475
Prevotella melaninogenica
Pmel2R
ATAGAACAGCGTGGTGGAGGATAAttttttttttttttt
24
55.8
180–203
GQ496463
Prevotella intermedia
Pint2-2
TTCAAGTCCGCCATCTTCACTTTTCtttttttttt
25
59.5
142–166
GQ496469
Propionibacterium acnes
Pacn4-2
TTGCTGTATGTGTTCGTGCGACtttttttttt
22
56.1
185–206
AB108459
Pacn8-2
GAGCATCTTATTTTTTGTGTGGCTTGTGttttttttttttttt
28
60.2
304–331
GQ496494
Propionibacterium granulosum
Pgra1-1
GGCGCACTGTTGTGTGTTCTTGTTGTtttttttttt
26
63
8–33
AF386070
Propionibacterium propionicus
Ppro1-1
CTGGTCATGGTTCTTGGTTTCTGGtttttttttt
24
58.5
86–109
GQ496478
Veillonella
spp.
Vpar1R
ACAGATCTCTCAAAACCGAACAATGtttttttttt
25
55.5
93–117
GQ496506
aOligonucleotide probes were arranged on the array as indicated in Fig. 1. Antisense probes are indicated by an R at the end of the probe code.
bTen or 15 additional bases of thymine (t) were added to the 3⬘end of each probe. The underlined nucleotides indicate mismatch nucleotides intentionally introduced into the probe to avoid nonspecific hybridization. The nucleotide R in probe Bfra18R, used to identifyBacteroides fragilis, represents a mixture of A and G.
cT
m, melting temperature.
dFor all probes except the positive control probe, the location of the probe in the ITS region is indicated by the nucleotide number of the ITS region. eGQ, accession numbers were determined in this study and submitted to GenBank.
fPositive control probes were designed from the 16S rRNA gene. gThe probe was specific for bothAnaerococcus prevotiiandA. tetradius.
on May 16, 2020 by guest
http://jcm.asm.org/
Species identification by array hybridization. The ITS region of the test bacterium was amplified by PCR with primer pair 2F and 10R, with each primer being labeled with a digoxigenin molecule at the 5⬘end. The reagents and procedures used for prehybridization, hybridization (50°C for 90 min), and color development with enzyme-conjugated antidigoxigenin antibodies were described previously (41). The hybridized spots (diameter, 400m) could be read by the naked eye. A strain was identified as one of the species listed in Table 1 when both the positive control probe and the species-specific probe (or at least one of the two probes designed to be specific for a species) were hybridized (Table 2). Identification was determined to the species level; subspecies-level identification was not considered.
Discrepancy analysis.In cases in which the result of array identification did not correspond with the original species name of a strain, the test with the Rapid ID 32A system was repeated to check the species name of the strain. If the result of one of the two tests with the Rapid ID 32A system agreed with that provided by the array, a concordant identification was considered for the strain. If the discrepancy continued to exist, the identity of the strain was determined by sequencing nearly the complete length of the 16S rRNA gene (27). The se-quences determined were used for a BLAST search of the sese-quences in public databases. The following criteria were used for identification: (i) when the com-parison of the sequence determined with a best-scoring reference sequence of a classified species yielded an identity ofⱖ99%, the isolate was assigned to that species; and (ii) when the identity was⬍99% andⱖ95%, the isolate was assigned to the corresponding genus (4). When discrepant identification occurred, the result of 16S rRNA gene sequencing was considered the final identification.
Definition of sensitivity, specificity, and detection limit.Sensitivity was defined as the number of target strains correctly identified (true positives) by the array divided by the total number of target strains tested. Specificity was defined as the number of nontarget strains producing negative hybridization reactions (true negatives) divided by the total number of nontarget strains tested (23). The detection limit was the smallest amount of bacterial DNA that could be detected by the array. Serial 10-fold dilutions of DNAs ofPeptostreptococcus anaerobius
CCUG 7835,Fusobacterium nucleatumsubsp. fusiformeJCM 11024, and Bacte-roides fragilisCCUG 4856 were used to determine the detection limit.
Direct detection of anaerobic bacteria in clinical specimens.A total of 49 clinical specimens were analyzed by use of the array, and the results were compared to those obtained by the conventional methods. The specimens in-cluded cerebrospinal fluid (7 samples), pleural effusion (10 samples), ascitic fluid (10 samples), synovial fluid (7 samples), bile (9 samples), pus (5 samples), and tissue (1 samples). All specimens were obtained from National Cheng Kung University Hospital. DNA was extracted from the clinical specimens with DNeasy blood and tissue kit (Qiagen, Hilden, Germany), and the ITS regions were amplified by seminested PCR. The first amplification was conducted with primers 11F (5⬘-GTTTGATCCTGGCTCAG-3⬘) and 10R (5⬘-TTCGCCTTTCC CTCACGGTA-3⬘). PCR was performed in a reaction volume of 25l consisting of 10 mM Tris-HCl (pH 8.8), 50 mM KCl, 1.5 mM MgCl2, 0.6 U GoTaq HotStart
polymerase (Promega, Madison, WI), 0.8 mM deoxyribonucleoside triphos-phates (0.2 mM each), and 1M (each) primer. The thermocycling conditions were as follows: initial denaturation at 95°C for 3 min; 25 cycles of denaturation (94°C, 1 min), annealing (55°C, 1 min), and extension (72°C, 1.5 min); and a final extension step at 72°C for 7 min. One microliter from the first reaction was then used for the second run of the PCR by use of the same thermocycling conditions, except that primer pair 2F (5⬘-TTCTACACACCGCCCGTC-3⬘) and 10R was used, each primer was labeled with a digoxigenin molecule at the 5⬘end, and 35 cycles of amplification were performed.
Nucleotide sequence accession numbers.The ITS sequences of some anaer-obes determined in this study were submitted to GenBank, and the accession numbers are reported in Table 2.
RESULTS
Probe design.
A total of 35 probes with high degrees of
sensitivity and specificity were spotted onto the array (Table 2).
For most species, a single probe was designed to identify an
individual species, but two probes were used to identify each of
the following species due to intraspecies ITS sequence
varia-tions in these species:
Anaerococcus prevotii Anaerococcus
tet-radius
,
Bacteroides fragilis
,
Finegoldia magna
,
Peptoniphilus
asaccharolyticus
, and
Propionibacterium acnes
(Table 2). It
should be noted that 1- or 2-base mismatches were
intention-ally incorporated into some probes to eliminate the nonspecific
hybridization caused by some nontarget bacteria (Table 2).
Anaerococcus prevotii
and
A. tetradius
had almost identical
ITS sequences (data not shown), and two probes (codes Apre4
and Apre5) were used to identify the two species as a group
(Table 2 and Fig. 2).
Fusobacterium varium
produced weak
cross-hybridization with the probe (code Fnec1-2) targeting
Fusobacterium necrophorum
(Fig. 2, chip 23). Therefore, a
strain was identified as
F. varium
if the species-specific probe
(code Fvar3) was hybridized, regardless of whether the
F.
necrophorum
probe (code Fnec1-2) was hybridized. Different
Veillonella
species had very high ITS sequence similarities
(data not shown), and hence, only a genus-specific probe (code
Vpar1R) was designed to identify the genus. All five species of
Veillonella
(
Veillonella atypica
,
V. dentocariosi
,
V. dispar
,
V.
montpellierensis
, and
V. parvula
) hybridized to the
genus-spe-cific probe (Fig. 2). Since subspecies-level identification was
not considered, all subspecies of
Fusobacterium nucleatum
(
F.
nucleatum
subsp.
animalis
,
F. nucleatum
subsp.
fusiforme
,
F.
nucleatum
subsp.
nucleatum
,
F. nucleatum
subsp.
polymor-phum
, and
F. nucleatum
subsp.
vincentii
) hybridized to a single
probe (code Fnuc6R) (Fig. 2).
Identification of reference strains by the array.
The
hybrid-ization patterns of 28 anaerobic bacterial species and
Veil-lonella
species are shown alphabetically in Fig. 2. Of 122
ref-erence strains analyzed, 118 hybridized to the respective
probes and were correctly identified, but 4 strains produced
discrepant identifications by use of the array (Table 3). The
array identified
Bacteroides ovatus
CCUG 35192 as
Bacteroides
thetaiotaomicron
,
Propionibacterium acnes
CCUG 4945 as
Pro-pionibacterium granulosum
, and
Propionibacterium propionicus
LMG 16717 as
Propionibacterium acnes
(Table 3). Sequence
analyses of the 16S rRNA genes clearly confirmed the accurate
identifications made by the array. Another discrepant
refer-ence strain (
Peptostreptococcus anaerobius
CCUG 49327) was
not identified by the array; however, 16S rRNA gene
sequenc-ing revealed that the strain was
Peptostreptococcus stomatis
, a
nontarget species in this study. In brief, 121 of the 122
refer-ence strains were correctly identified to the species level by
the array, with the remaining strain (CCUG 49327) being a
nontarget species (
P. stomatis
). Therefore, the sensitivity of
the array for the identification of reference strains was 100%
(121/121).
Identification of clinical isolates by the array.
Of 188 target
[image:4.585.42.285.68.155.2]clinical isolates, the array yielded 20 discrepant identifications.
FIG. 1. Layout of oligonucleotide probes on the array (0.7 by 0.7 cm,
7 by 7 dots). Probe PC (C7) was a positive control, and probe NC was a
negative control (tracking dye only). Probe M, a position marker, was an
irrelevant probe labeled with digoxigenin at the 5
⬘
end. The corresponding
species names and sequences of all probes are listed in Table 2.
on May 16, 2020 by guest
http://jcm.asm.org/
Repeat testing of these discordant strains with the Rapid ID
32A system reduced the number of discordant strains to 11
(Table 3). Of these 11 strains, four (
Anaerococcus prevotii
A776-1,
Anaerococcus prevotii
C270,
Fusobacterium nucleatum
365, and
Fusobacterium nucleatum
5084N2) were correctly
identified by the array, as evidenced by the results of 16S
rRNA gene sequencing. One target isolate (
Finegoldia magna
D752-3) was not identified. The remaining six strains (
Anaero-coccus prevotii
C400-3,
Fusobacterium nucleatum
K789-2,
Pep-toniphilus asaccharolyticus
B619-2,
Peptoniphilus
asaccharolyti-cus
C302-2,
Peptoniphilus
asaccharolyticus
3480N,
and
Parvimonas micra
I503) were actually nontarget isolates.
Among these six strains, strains C400-3 and K789-2 were
mis-identified as
Finegoldia magna
and
Fusobacterium
necropho-rum
, respectively, and the remaining four strains were not
identified by the array. In addition, one nontarget isolate (
Bac-teroides caccae
483) was identified as
B. fragilis
by the array,
and the identification was confirmed by 16S rRNA gene
se-quencing (Table 3). In summary, the total number of target
clinical isolates was 183 (188
⫺
6
⫹
1) and 182 isolates were
correctly identified, resulting in a test sensitivity of 99.5% (182/
183). If reference strains and clinical isolates were taken
together, the sensitivity of the array for the identification of
anaerobic bacteria was 99.7% [(182
⫹
121)/(183
⫹
121)].
Specificity and detection limit of the array.
A collection of
98 nontarget strains (51 species), including anaerobic and
aerobic bacteria, were used for specificity testing (see Table
S1 in the supplemental material).
Bacteroides caccae
L117
was misidentified as
B. vulgatus
by the array; however, the
strain was determined to be
B. dorei
by its 16S rRNA gene
sequence (Table 3). In addition to the 98 clinical nontarget
isolates, another 7 strains (
Peptostreptococcus anaerobius
CCUG 49327;
Anaerococcus prevotii
C400-3;
Fusobacterium
nucleatum
K789-2;
Peptoniphilus asaccharolyticus
B619-2,
C302-2, and 3480N; and
Parvimonas micra
I503), initially
included as target strains, were found to be nontarget
mi-FIG. 2. Hybridization patterns of 28 species of anaerobic bacteria and
Veillonella
spp. All strains except
Anaerococcus prevotii
CCUG 44020A
and
Peptoniphilus asaccharolyticus
CCUG 12549 were type strains. The corresponding probes hybridized on the arrays are indicated in Fig. 1, and
the corresponding sequences of the hybridized probes are shown in Table 2.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:5.585.46.542.68.473.2]croorganisms through discrepant analysis (Table 3). Among
the seven nontarget strains, two (C400-3 and K789-2) were
misidentified as target species. On the contrary, one
non-target isolate (
Bacteroides caccae
483) was found to be
B.
fragilis
, a target isolate. Therefore, a total of 104 (98
⫹
7
⫺
1) nontarget strains were analyzed by the array and three
strains (C400-3, K789-2, and L117) were misidentified,
re-sulting in an identification specificity of 97.1% (101/104).
The detection limits of the array ranged from 10 fg (
Pep-tostreptococcus anaerobius
CCUG 7835 and
Fusobacterium
nucleatum
subsp
. fusiforme
JCM 11024) to 100 fg (
Bacte-roides fragilis
CCUG 4856) per assay. If a bacterial cell has
about 4 fg of DNA (18), the detection limits of the array
were from 3 to 30 cells per assay.
Detection of anaerobic bacteria in clinical specimens.
A
total of 49 clinical specimens were analyzed by the array. The
array detected two species (
Finegoldia magna
and
Bacteroides
vulgatus
) in two pus samples, and the results corresponded to
those obtained by culture.
DISCUSSION
In this study, an oligonucleotide array was developed to
identify 28 species of clinically important anaerobic bacteria
and
Veillonella
spp. The sensitivity and the specificity of the
array were 99.7 and 97.1%, respectively.
Four strains (
Bacteroides ovatus
CCUG 35192,
Peptostrepto-coccus anaerobius
CCUG 49327,
Propionibacterium acnes
CCUG 4945, and
Propionibacterium propionicus
LMG 16717)
from two strain collection centers were given the wrong species
names, as evidenced by the results obtained with the array and
by 16S rRNA gene sequencing (Table 3). Two of the four
strains belonged to the genus
Propionibacterium
, which
con-firms that the phenotypic identification of the propionibacteria
is still problematic and that alternative identification
tech-niques are required for this genus (28). Three
Propionibacte-rium
species, i.e.,
P. acnes
,
P. granulosum
, and
P. propionicus
,
were included in this study and were well differentiated from
one another by the array (Fig. 2).
Bacteria in the
Bacteroides fragilis
group are important
pathogens in polymicrobial infections. The group includes
B.
fragilis
,
B. thetaiotaomicron
,
B. vulgatus
,
B. ovatus
,
B. distasoni
s,
[image:6.585.44.540.80.424.2]B. uniformis
, and other species, with
B. thetaiotaomicron
being
much more resistant to many antimicrobials (5, 9). The group
accounted for as many as 61% of the anaerobic isolates
recov-ered from blood cultures (3). The members of the group are
phenotypically very similar and are frequently misidentified by
biochemical tests. Since different species in the
B. fragilis
group
TABLE 3. Analysis of strains that produced discrepant identifications by the array
Strain no. Species identity upon receipt
Species identification (% of strainsa) by:
Best match Oligonucleotide array analysis 16S rRNA gene sequencing
Target strain
CCUG 35192
Bacteroides ovatus
Bacteroides thetaiotaomicron
B. thetaiotaomicron
(99.3)
B. thetaiotaomicron
B. ovatus
(95.6)
CCUG 49327
Peptostreptococcus anaerobius
Not identified
P. stomatis
(99.8)
P. stomatis
P. anaerobius
(98.8)
CCUG 4945
Propionibacterium acnes
Propionibacterium
P. granulosum
(99.3)
P. granulosum
granulosum
P. acnes
(94.0)
LMG 16717
Propionibacteriun propionicus
Propionibacteriun acnes
P. acnes
(99.7)
P. acnes
P. propionicus
(91.4)
A776-1
Anaerococcus prevotii
Peptoniphilus asaccharolyticus
P. asaccharolyticus
(99.3)
P. asaccharolyticus
A. prevotii
(85.0)
C270
Anaerococcus prevotii
Finegoldia magna
F. magna
(99.2)
F. magna
A. prevotii
(82.5)
C400-3
Anaerococcus prevotii
Finegoldia magna
F. magna
(98.9)
Finegoldia
sp.
A. prevotii
(82.6)
D752-3
Finegoldia magna
Not identified
F. magna
(99.3)
F. magna
365
Fusobacterium nucleatum
Fusobacterium necrophorum
F. necrophorum
(99.3)
F. necrophorum
F. nucleatum
(90.0)
5084N2
Fusobacterium nucleatum
Fusobacterium necrophorum
F. necrophorum
(99.4),
F. necrophorum
F. nucleatum
(91.3)
K789-2
Fusobacterium nucleatum
Fusobacterium necrophorum
F. gonidiaformans
(98.3)
Fusobacterium
sp.
F. necrophorum
(96.9)
B619-2
Peptoniphilus asaccharolyticus
Not identified
Peptoniphilus
sp. (98.4)
Peptoniphilus
sp.
P. asaccharolyticus
(95.9)
C302-2
Peptoniphilus asaccharolyticus
Not identified
Peptoniphilus
sp. (88.3)
An unknown species
P. asaccharolyticus
(83.6)
3480N
Peptoniphilus asaccharolyticus
Not identified
Peptoniphilus
sp. (98.2)
Peptoniphilus
sp.
P. asaccharolyticus
(96.8)
I503
Parvimonas micra
Not identified
Solobacterium moorei
(99.6)
S. moorei
P. micra
(52.7)
Nontarget strain
483
Bacteroides caccae
Bacteroides fragilis
B. fragilis
(99.6)
B. fragilis
L117
Bacteroides caccae
Bacteroides vulgatus
B. dorei
(99.9)
B. dorei
a
Values in parentheses are percentages of 16S rRNA gene sequence similarity of the isolates with the best-scoring sequences in GenBank.
on May 16, 2020 by guest
http://jcm.asm.org/
vary in their resistance to antimicrobial agents (40), it is
im-portant to differentiate species in the group, especially for
severe infections (44). The taxonomy of
Bacteroides
has
under-gone major revisions in the last few decades, with more than 20
species now being included in the genus (29, 43). In this study,
oligonucleotide probes were successfully applied to
differenti-ate the five important members (
B. fragilis
,
B. ovatus
,
B.
the-taiotaomicron
,
B. uniformis
, and
B. vulgatus
) of the
B. fragilis
group (Fig. 2).
Gram-positive anaerobic cocci are a heterogeneous group of
organisms, with the different species displaying major
differ-ences in antimicrobial susceptibility patterns (25). In this study,
three isolates of
Anaerococcus prevotii
(isolates A776-1, C270,
and C400-3) were found to be misidentifications of
Peptoniphi-lus asaccharolyticus
or
Finegoldia
species (Table 3). In addition,
three clinical isolates (isolates B619-2, C302-2, and 3480N) of
Peptoniphilus asaccharolyticus
were not identified to the species
level by the array or by 16S rRNA gene sequencing, suggesting
that the three isolates were misidentified by conventional
bio-chemical tests (Table 3). Since the three
Peptoniphilus
isolates
were recovered from blood cultures, their significance and real
identities warrant further investigation.
Fusobacterium nucleatum
and
F. necrophorum
can cause
manifold infections, such as periodontitis, organ abscesses, and
bacteremia (30). Two clinical
F. nucleatum
isolates (isolates
365 and 5084N2) were misidentifications of
F. necrophorum
, as
revealed by the array and 16S rRNA gene sequencing (Table
3). The identification of clinically relevant
Fusobacterium
spp.
is hampered by their slow growth and the low levels of
reli-ability of biochemical tests (30, 38). In the present study,
F.
necrophorum
was well differentiated from
F. nucleatum
,
includ-ing several subspecies (Table 1 and Fig. 2).
The array was also assessed for its ability to directly detect
anaerobic bacteria in 49 clinical specimens. Two species
(
Finegoldia magna
and
Bacteroides vulgatus
) were detected in two
specimens by the array, and the results were in accordance with
those obtained by culture. However, instead of PCR, nested
PCR was required to produce enough amplicons for
hybrid-ization. These results indicate that the array may have the
potential to detect anaerobic bacteria in clinical specimens.
However, the number and types of clinical specimens tested in
this study were limited, and further comprehensive evaluation
is needed to validate this potential. The low rate of detection
(4.1%) of anaerobic bacteria in these samples might be due to
the use of a high percentage of sterile body specimens.
In conclusion, identification of the species of clinically
rele-vant anaerobes by use of the array described here is highly
reliable. The method could be used as an accurate alternative
to the conventional methods if adequate species identification
is of concern. The whole procedure of array hybridization took
about 8 h, starting with the isolated colonies.
ACKNOWLEDGMENTS
This study was supported by grants from the National Science
Coun-cil (grant 96-2320-B-006-024-MY3), the Center for Frontier Materials
and Micro/Nano Science and Technology (grant D97-2720), National
Cheng Kung University, and the Department of Health (grant
DOH-99-TD-B-111-002), Taiwan, Republic of China.
REFERENCES
1.Aldridge, K. E., D. Ashcraft, K. Cambre, C. L. Pierson, S. G. Jenkins, and J. E. Rosenblatt.2001. Multicenter survey of the changing in vitro antimi-crobial susceptibilities of clinical isolates ofBacteroides fragilisgroup, Pre-votella,Fusobacterium,Porphyromonas, andPeptostreptococcusspecies. An-timicrob. Agents Chemother.45:1238–1243.
2.Bengualid, V., H. Singh, V. Singh, and J. Berger.2008. An increase in the incidence of anaerobic bacteremia: true for tertiary care referral centers but not for community hospitals? Clin. Infect. Dis.46:323–324.
3.Blairon, L., Y. De Gheldre, B. Delaere, A. Sonet, A. Bosly, and Y. Glupczyn-ski.2006. A 62-month retrospective epidemiological survey of anaerobic bacteraemia in a university hospital. Clin. Microbiol. Infect.12:527–532. 4.Bosshard, P. P., R. Zbinden, S. Abels, B. Bo¨ddinghaus, M. Altwegg, and
E. C. Bo¨ttger.2006. 16S rRNA gene sequencing versus the API 20 NE system and the VITEK 2 ID-GNB card for identification of nonfermenting Gram-negative bacteria in the clinical laboratory. J. Clin. Microbiol.44:1359–1366. 5.Brook, I.1989. Pathogenicity of theBacteroides fragilisgroup. Ann. Clin. Lab.
Sci.19:360–376.
6.Brook, I.2002. Clinical review: bacteremia caused by anaerobic bacteria in children. Crit. Care6:205–211.
7.Brown, T. J., and R. M. Anthony.2000. The addition of low numbers of 3⬘
thymine bases can be used to improve the hybridization signal of oligonu-cleotides for use within arrays on nylon supports. J. Microbiol. Methods
42:203–207.
8.Chang, H. C., Y. F. Wei, L. Dijkshoorn, M. Vaneechoutte, C. T. Tang, and T. C. Chang.2005. Species-level identification of isolates of theAcinetobacter calcoaceticus-Acinetobacter baumanniicomplex by sequence analysis of the 16S-23S rRNA gene spacer region. J. Clin. Microbiol.43:1632–1639. 9.Citron, D. M., I. R. Poxton, and E. J. Baron.2007.Bacteroides,
Porphyromo-nas,Prevotella,Fusobacterium, and other anaerobic Gram-negative rods, p. 911–932.InP. R. Murray, E. J. Baron, J. H. Jorgensen, M. L. Landry, and M. A. Pfaller (ed.), Manual of clinical microbiology, 9th ed. American Society for Microbiology, Washington, DC.
10.Downes, J., A. King, J. Hardie, and I. Phillips.1999. Evaluation of the Rapid ID 32A system for identification of anaerobic Gram-negative bacilli, exclud-ing theBacteroides fragilisgroup. Clin. Microbiol. Infect.5:319–326. 11.Fenner, L., A. F. Widmer, C. Straub, and R. Frei.2008. Is the incidence of
anaerobic bacteremia decreasing? Analysis of 114,000 blood cultures over a ten-year period. J. Clin. Microbiol.46:2432–2434.
12.Fukushima, M., K. Kakinuma, H. Hayashi, H. Nagai, K. Ito, and R. Kawaguchi.2003. Detection and identification ofMycobacteriumspecies isolates by DNA microarray. J. Clin. Microbiol.41:2605–2615.
13.Hecht, D. W.2004. Prevalence of antibiotic resistance in anaerobic bacteria: worrisome developments. Clin. Infect. Dis.39:92–97.
14.Igarashi, E., A. Kamaguchi, M. Fujita, H. Miyakawa, and F. Nakazawa.
2009. Identification of oral species of the genusVeillonellaby polymerase chain reaction. Oral Microbiol. Immunol.24:310–313.
15.Jenkins, S. A., D. B. Drucker, M. G. Keaney, and L. A. Ganguli.1991. Evaluation of the RAPID ID 32A system for the identification ofBacteroides fragilisand related organisms. J. Appl. Bacteriol.71:360–365.
16.Kitch, T. T., and P. C. Appelbaum.1989. Accuracy and reproducibility of the 4-hour ATB 32A method for anaerobe identification. J. Clin. Microbiol.
27:2509–2513.
17.Ko, W.-C., N.-Y. Lee, S. C. Su, L. Dijkshoorn, M. Vaneechoutte, L.-R. Wang, J.-J. Yan, and T. C. Chang.2008. Oligonucleotide array-based identification of species in theAcinetobacter calcoaceticus-A. baumanniicomplex isolated from blood cultures and antimicrobial susceptibility testing of the isolates. J. Clin. Microbiol.46:2052–2059.
18.Kubitschek, H. E., and M. C. Friedman.1971. Chromosome replication and the division cycle ofEscherichia coliB/r. J. Bacteriol.107:95–99. 19.Kuwahara, T., I. Norimatsu, H. Nakayama, S. Akimoto, K. Kataoka, H.
Arimochi, and Y. Ohnishi.2001. Genetic variation in 16S-23S rDNA internal transcribed spacer regions and the possible use of this genetic variation for molecular diagnosis ofBacteroidesspecies. Microbiol. Immunol.45:191–199. 20.Lassmann, B., D. R. Gustafson, C. M. Wood, and J. E. Rosenblatt.2007.
Reemergence of anaerobic bacteremia. Clin. Infect. Dis.44:895–900. 21.Lau, S. K., J. L. Teng, K. W. Leung, N. K. Li, K. H. Ng, K. Y. Chau, T. L. Que,
P. C. Woo, and K. Y. Yuen.2006. Bacteremia caused bySolobacterium moorei
in a patient with acute proctitis and carcinoma of the cervix. J. Clin. Micro-biol.44:3031–3034.
22.Marchandin, H., C. Teyssier, M. Sime´on De Buochberg, H. Jean-Pierre, C. Carriere, and E. Jumas-Bilak.2003. Intra-chromosomal heterogeneity be-tween the four 16S rRNA gene copies in the genusVeillonella: implications for phylogeny and taxonomy. Microbiology149:1493–1501.
23.McClure, F. D.1990. Design and analysis of quantitative collaborative stud-ies: minimum collaborative program. J. Assoc. Off. Anal. Chem.73:953–960. 24.Millar, B. C., X. Jiru, J. E. Moore, and J. A. Earle.2000. A simple and sensitive method to extract bacterial, yeast and fungal DNA from blood culture material. J. Microbiol. Methods42:139–147.
25.Murdoch, D. A.1998. Gram-positive anaerobic cocci. Clin. Microbiol. Rev.
11:81–120.
on May 16, 2020 by guest
http://jcm.asm.org/
26.Nonnenmacher, C., A. Dalpke, R. Mutters, and K. Heeg.2004. Quantitative detection of periodontopathogens by real-time PCR. J. Microbiol. Methods
59:117–125.
27.Relman, D. A.1993. Universal bacterial 16S rDNA amplification and se-quencing, p. 489–495,InD. H. Persing, F. C. Tenover, T. F. Smith, and T. J. White (ed.), Diagnostic molecular microbiology. American Society for Mi-crobiology, Washington, DC.
28.Riedel, K. H., B. D. Wingfield, and T. J. Britz.1998. Identification of classical Propionibacterium species using 16S rDNA-restriction fragment length poly-morphisms. Syst. Appl. Microbiol.21:419–428.
29.Sakamoto, M., and Y. Benno.2006. Reclassification ofBacteroides distasonis,
Bacteroides goldsteinii, andBacteroides merdaeasParabacteroides distasonis
gen. nov, comb. nov.,Parabacteroides goldsteiniicomb. nov., and Parabacte-roides merdaecomb. nov. Int. J. Syst. Bacteriol.56:1599–1605.
30.Sigge, A., A. Essig, B. Wirths, K. Fickweiler, N. Kaestner, N. Wellinghausen, and S. Poppert.2007. Rapid identification ofFusobacterium nucleatumand
Fusobacterium necrophorumby fluorescence in situ hybridization. Diagn. Microbiol. Infect. Dis.58:255–259.
31.Simmon, K. E., S. Mirrett, L. B. Reller, and C. A. Petti.2008. Genotypic diversity of anaerobic isolates from bloodstream infections. J. Clin. Micro-biol.46:1596–1601.
32.Siqueira, J. F., Jr., I. N. Rocas, A. F. Andrade, and M. de Uzeda.2003.
Peptostreptococcus microsin primary endodontic infections as detected by 16S rDNA-based polymerase chain reaction. J. Endod.29:111–113. 33.Snydman, D. R., N. V. Jacobus, L. A. McDermott, R. Ruthazer, Y. Golan,
E. J. C. Goldstein, S. M. Finegold, L. J. Harrell, D. W. Hecht, S. G. Jenkins, C. Pierson, R. Venezia, V. Yu, J. Rihs, and S. L. Gorbach.2007. National survey on the susceptibility ofBacteroides fragilisgroup: report and analysis of trends in the United States from 1997 to 2004. Antimicrob. Agents Che-mother.51:1649–1655.
34.Song, Y., C. Liu, M. Bolanos, J. Lee, M. McTeague, and S. M. Finegold.2005. Evaluation of 16S rRNA sequencing and reevaluation of a short biochemical scheme for identification of clinically significantBacteroidesspecies. J. Clin. Microbiol.43:1531–1537.
35.Song, Y., C. Liu, and S. M. Finegold.2007. Development of a flow chart for identification of gram-positive anaerobic cocci in the clinical laboratory. J. Clin. Microbiol.45:512–516.
36.Stingu, C. S., A. C. Rodloff, H. Jentsch, R. Schaumann, and K. Eschrich.
2008. Rapid identification of oral anaerobic bacteria cultivated from subgin-gival biofilm by MALDI-TOF-MS. Oral Microbiol. Immunol.23:372–376. 37.Strauss, J., A. White, C. Ambrose, J. McDonald, and E. Allen-Vercoe.2008.
Phenotypic and genotypic analyses of clinicalFusobacterium nucleatumand
Fusobacterium periodonticumisolates from the human gut. Anaerobe14:
301–309.
38.Stubbs, S. L., J. S. Brazier, P. R. Talbot, and B. I. Duerden.2000. PCR-restriction fragment length polymorphism analysis for identification of Bac-teroidesspp. and characterization of nitroimidazole resistance genes. J. Clin. Microbiol.38:3209–3213.
39.Teng, L. J., P. R. Hsueh, J. C. Tsai, S. J. Liaw, S. W. Ho, and K. T. Luh.2002. High incidence of cefoxitin and clindamycin resistance among anaerobes in Taiwan. Antimicrob. Agents Chemother.46:2908–2913.
40.Tung, S. K., L. J. Teng, M. Vaneechoutte, H. M. Chen, and T. C. Chang.
2006. Array-based identification of species of the generaAbiotrophia,
Enterococcus,Granulicatella, andStreptococcus. J. Clin. Microbiol. 44:4414– 4424.
41.Tung, S. K., L. J. Teng, M. Vaneechoutte, H. M. Chen, and T. C. Chang.
2007. Identification of species ofAbiotrophia,Enterococcus,Granulicatella
andStreptococcusby sequence analysis of the ribosomal 16S-23S intergenic spacer region. J. Med. Microbiol.56:504–513.
42.Wexler, H. M.2007.Bacteroides: the good, the bad, and the nitty-gritty. Clin. Microbiol. Rev.20:593–621.
43.Wybo, I., D. Pierard, I. Verschraegen, M. Reynders, K. Vandoorslaer, G. Claeys, M. Delmee, Y. Glupczynski, B. Gordts, M. Ieven, P. Melin, M. Struelens, J. Verhaegen, and S. Lauwers.2007. Third Belgian multicentre survey of antibiotic susceptibility of anaerobic bacteria. J. Antimicrob. Che-mother.59:132–139.