0095-1137/09/$08.00
⫹
0
doi:10.1128/JCM.00544-09
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Single Nucleotide Polymorphisms in the IS
900
Sequence of
Mycobacterium avium
subsp.
paratuberculosis
Are
Strain Type Specific
䌤
Elena Castellanos,
1,2Alicia Aranaz,
1,2* Lucia de Juan,
1,2Julio A
´ lvarez,
1Sabrina Rodríguez,
1,2Beatriz Romero,
1,2Javier Bezos,
1,2Karen Stevenson,
3Ana Mateos,
1,2and Lucas Domínguez
1,2Centro de Vigilancia Sanitaria Veterinaria (VISAVET), Universidad Complutense de Madrid, Avda. Puerta de Hierro s/n, 28040 Madrid,
Spain
1; Departamento de Sanidad Animal, Facultad de Veterinaria, Universidad Complutense de Madrid,
Madrid, Spain
2; and Moredun Research Institute, Division of Control of Bacterial Diseases,
Pentlands Science Park, Bush Loan, Penicuik, EH26 OPZ Scotland, United Kingdom
3Received 17 March 2009/Accepted 27 April 2009
Insertion sequence IS
900
is used as a target for the identification of
Mycobacterium avium
subsp.
paratu-berculosis.
Previous reports have revealed single nucleotide polymorphisms within IS
900
. This study, which
analyzed the IS
900
sequences of a panel of isolates representing
M. avium
subsp.
paratuberculosis
strain types
I, II, and III, revealed conserved type-specific polymorphisms that could be utilized as a tool for diagnostic and
epidemiological purposes.
The insertion sequence IS
900
(13) is one of the 20 members
of the IS
110
family and is considered to be unique to
Myco-bacterium avium
subsp.
paratuberculosis
, although IS
900
-like
sequences have been found rarely in other environmental
my-cobacteria (9, 12). IS
900
is present in multiple copies (14–18)
within the
M. avium
subsp.
paratuberculosis
genome (13) and
therefore is an ideal target for identification (16, 31). The
heterogeneity of
M. avium
subsp.
paratuberculosis
isolates
based on the number of IS
900
copies and their positions within
the genome has been exploited for epidemiological purposes.
The IS
900
sequence has been used in different molecular
techniques such as multiplex PCR and restriction fragment
length polymorphism analysis with hybridization to IS
900
,
leading to the identification of individual strains and the
classification of
M. avium
subsp.
paratuberculosis
into strain
types (3, 17, 19, 30).
M. avium
subsp.
paratuberculosis
strains have been divided
into three clusters named type I (also called sheep [S]
strains), type II (also called cattle [C] strains), and type III
(also called intermediate strains), although some of the
studies include types I and III in the S type (8, 35). This
classification into three clusters is based on results from
molecular characterization techniques such as restriction
fragment length polymorphism analysis with hybridization
to IS
900
(19), pulsed-field gel electrophoresis (PFGE) (11,
29), PCR-restriction enzyme analysis (PCR-REA) of
gyrB
(5), PCR-REA of
inhA
(6), PCR and denaturing gradient
gel electrophoresis analyses of MAP1506 (14), PCR
se-quencing of
recF
(31), and comparative genomic
hybridiza-tion analyses (4). The genotypic and phenotypic
dissimilar-ities among types of
M. avium
subsp.
paratuberculosis
may be
reflected in the differences detected in the progression of
paratuberculosis or Johne’s disease among infected herds
(33, 34).
Comparisons of IS
900
sequences from different
M. avium
subsp.
paratuberculosis
isolates with the published genome
se-quence of
M. avium
subsp.
paratuberculosis
strain K-10 have
revealed single nucleotide polymorphisms (SNPs) (2, 20, 21,
23, 24, 27, 28, 35, 36) (Table 1).
Of special interest are the results reported by Semret et al.
(24), who observed major variance in the IS
900
sequences in
M. avium
subsp.
paratuberculosis
S strains (classification into
type I or III was not indicated in the study) compared to the
IS
900
sequences in C strains.
The aim of this research was to follow up these findings and
to analyze the IS
900
sequences from a panel of type I, II, and
III
M. avium
subsp.
paratuberculosis
isolates to establish the
degree of nucleotide identity among them and the relationship
between
M. avium
subsp.
paratuberculosis
types and IS
900
se-quence profiles.
For this purpose, we selected 33 isolates of
M. avium
subsp.
paratuberculosis
types I, II, and III from separate
geographical locations throughout Spain, Scotland, and
Denmark (Table 2). Every isolate was cultured and
identi-fied as
M. avium
subsp.
paratuberculosis
by PCRs directed to
the gene F57 (22) (using primers F57-F, 5
⬘
CCCGATAGC
TTTCCTCTCCT 3
⬘
, and F57-R, 5
⬘
GATCTCAGACAGTG
GCAGGTG 3
⬘
[7]). Additionally, the isolates were typed
using a set of PCR-based assays, REA targeting IS
1311
(15),
PCR analysis of the genomic region described by Collins et
al. (DMC) (8), analysis of large sequence polymorphism
A20 (LSP
A20) (25), and
hsp65
sequencing (32) (Table 3).
Then the isolates were classified as type I, II, or III by
PFGE, PCR-REA of
gyrB
, or PCR-REA of
inhA
as
de-scribed previously (5, 6, 10, 11).
* Corresponding author. Mailing address: Departamento de Sanidad
Animal, Facultad de Veterinaria, Universidad Complutense de Madrid,
Avda. Puerta de Hierro s/n, 28040 Madrid, Spain. Phone: (34) 913943721.
Fax: (34) 913943795. E-mail: [email protected].
䌤
Published ahead of print on 13 May 2009.
2260
on May 16, 2020 by guest
http://jcm.asm.org/
A fragment of 662 bp of the IS
900
sequence from each of the
isolates was amplified by PCR directed to the 5
⬘
end of the
insertion sequence, considered to be specific to
M. avium
subsp.
paratuberculosis
(9, 18) and found previously to be
[image:2.585.44.542.88.385.2]poly-morphic (24). Primers IS
900
-F (5
⬘
CCTTTCTTGAAGGGTG
TTCG 3
⬘
[24]) and IS
900
-R (5
⬘
CCACCAGATCGGAA
CGTC 3
⬘
) were used in the amplification reaction, and then
amplicons were purified with a QIAquick PCR purification kit
TABLE 1. Previously described SNPs in the 5
⬘
fragments of IS900
sequences relative to the
M. avium
subsp.
paratuberculosis
K-10 genome sequence
aReference Strain
type
Geographic
origin Host(s) or source
Molecular
technique SNP
Position (bp)
GenBank accession no. Bhide et al.,
2006 (2)
⫺ ⫺ Cattle crossbreedBos
indicus
SSCPb
analysis No nucleotide (instead of A) 284 AY974345
⫺ ⫺ Cattle SSCP analysis No nucleotide (instead of A) 284 AY974346
CG (instead of GC) 367–368
⫺ Slovakia Slovak Valachian sheep SSCP analysis C (instead of G) 232 AY974348
T (instead of A) 243
CG (instead of GC) 367–368
Pickup et al., ⫺ United Kingdom River water Sequencing G (instead of A) 214c ⫺
2005 (20) A/G 214c
Pickup et al., ⫺ United Kingdom River water Sequencing G (instead of A) 216 ⫺
2006 (21) A/G 216
Semret et al., 2006 (24)
S ⫺ Sheep Sequencing T/C (with a C small peak) 169 ⫺
S ⫺ Sheep Sequencing G/A (with an A small peak) 216 ⫺
Scanu et al.,
2007 (23) ⫺
Italy Humans, Sardinian sheep Sequencing C/T (with a T small peak) 247d ⫺
Sivakumar et al.,
2005 (27) ⫺
India Buffalo Sequencing No nucleotide (instead of G) 688 AY660657
Extra C 722
T (instead of A) 899
⫺ India Goats Sequencing G (instead of A) 422 AY660658
No nucleotide (instead of G) 688
Extra C 722
Sohal et al., 2009 (28)
Bison India Jamunapari goats Sequencing Extra A 421e
EF514832
Willemsen et al., 1999 (36)
Bovine ⫺ 316F vaccine Sequencing Extra GC 35–36 AF416985
GC (instead of CG) 119–120
Whittington et al., 2001 (35)
S Australia Sheep Sequencing G (instead of A) 216 ⫺
a⫺
, no information was provided in the reference.
b
SSCP, single-strand conformational polymorphism.
c
The SNP corresponds to position 216 in the IS900sequence.
d
The SNP was reported to occur at bp 247, but according to the published IS900sequence ofM. aviumsubsp.paratuberculosisK-10 (GenBank accession no. AE
016958), corresponds to bp 244.
e
The observed SNP is relative to the IS900sequence at locus 17 ofM. aviumsubsp.paratuberculosisK-10.
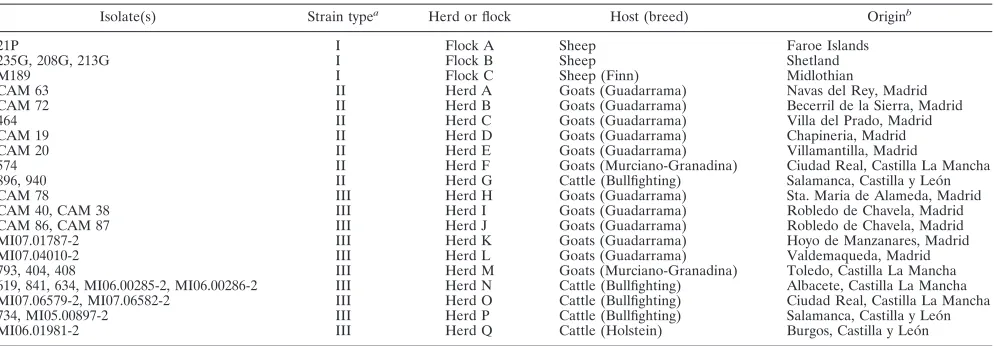
TABLE 2. Panel of
M. avium
subsp.
paratuberculosis
isolates used in this study
Isolate(s) Strain typea
Herd or flock Host (breed) Originb
21P I Flock A Sheep Faroe Islands
235G, 208G, 213G I Flock B Sheep Shetland
M189 I Flock C Sheep (Finn) Midlothian
CAM 63 II Herd A Goats (Guadarrama) Navas del Rey, Madrid
CAM 72 II Herd B Goats (Guadarrama) Becerril de la Sierra, Madrid
464 II Herd C Goats (Guadarrama) Villa del Prado, Madrid
CAM 19 II Herd D Goats (Guadarrama) Chapineria, Madrid
CAM 20 II Herd E Goats (Guadarrama) Villamantilla, Madrid
574 II Herd F Goats (Murciano-Granadina) Ciudad Real, Castilla La Mancha
896, 940 II Herd G Cattle (Bullfighting) Salamanca, Castilla y Leo´n
CAM 78 III Herd H Goats (Guadarrama) Sta. Maria de Alameda, Madrid
CAM 40, CAM 38 III Herd I Goats (Guadarrama) Robledo de Chavela, Madrid
CAM 86, CAM 87 III Herd J Goats (Guadarrama) Robledo de Chavela, Madrid
MI07.01787-2 III Herd K Goats (Guadarrama) Hoyo de Manzanares, Madrid
MI07.04010-2 III Herd L Goats (Guadarrama) Valdemaqueda, Madrid
793, 404, 408 III Herd M Goats (Murciano-Granadina) Toledo, Castilla La Mancha
619, 841, 634, MI06.00285-2, MI06.00286-2 III Herd N Cattle (Bullfighting) Albacete, Castilla La Mancha
MI07.06579-2, MI07.06582-2 III Herd O Cattle (Bullfighting) Ciudad Real, Castilla La Mancha
734, MI05.00897-2 III Herd P Cattle (Bullfighting) Salamanca, Castilla y Leo´n
MI06.01981-2 III Herd Q Cattle (Holstein) Burgos, Castilla y Leo´n
a
Every isolate was classified as type I, II, or III by PFGE (10, 11), PCR-REA ofgyrB(5), or PCR-REA ofinhA(6).
b
Geographic distribution: Faroe Islands, Denmark; Shetland, northern Scotland; Midlothian, southeast Scotland; Madrid, central Spain; Castilla La Mancha, south
central Spain; and Castilla y Leo´n, north central Spain.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:2.585.45.542.526.699.2](Qiagen, GmbH) and both strands were sequenced by using an
ABI Prism 3730 DNA sequencer (Applied Biosystems; CIB
Sequencing Facilities, Madrid, Spain). Forward and reverse
sequences were aligned with the IS
900
elements of the
pub-lished genome sequence of
M. avium
subsp.
paratuberculosis
K-10 (type II strain; GenBank accession no. AE016958;
refer-ence sequrefer-ence no. NC_002944), as suggested by Semret et al.
(24). The analysis of the chromatograms showed IS
900
SNPs
that were homogeneous, conserved, and dependent on the
M.
avium
subsp.
paratuberculosis
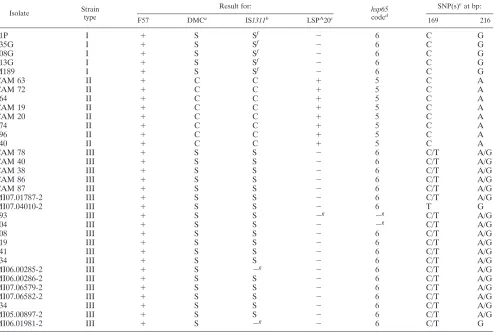
type (Table 3). Every type I ovine
isolate showed an SNP at position 216, with a G instead of an
A, and no other sequence modifications. This SNP was
ob-served previously in a sample extracted directly from river
water (20, 21), in an isolate from a sheep in Australia (35), and
in some ovine isolates analyzed by Semret et al. (24). The eight
isolates of type II subjected to analysis displayed complete
homology to
M. avium
subsp.
paratuberculosis
K-10. However,
for the type III isolates tested, double peaks of the same size at
bp 169 (T/C) and bp 216 (G/A) or a single T peak and a G
were noticed. For four of these isolates with ambiguities,
another DNA extraction from a single colony was
per-formed with an inoculating needle, but the results were
corroborated, ruling out the possibility of contamination by
two different strain types. The results of this analysis match
previous observations of these SNPs in some of the S
iso-lates tested by Semret et al. (24). This pattern is probably
indicative of the presence of a point mutation in some of the
copies of IS
900
(Table 3) and also confirms the presence of
type III isolates outside Spain (10). To our knowledge, this
is the first report to establish a correlation between IS
900
polymorphisms and strain types I, II, and III. Interestingly,
in two previous studies, an irresolvable A/G polymorphism
at position 216 (reported initially to be at bp 214) was
observed in two samples obtained from a river flow (20, 21),
due maybe to the presence of both types I and II in the
samples collected. On the other hand, none of our isolates
revealed any other SNPs described previously in the
litera-ture (2, 23, 27, 28, 36).
The type III strains exhibited polymorphisms identical to
those of type I strains, as determined by PCR analysis of DMC,
IS
1311
REA, and LSP
A20 and
hsp65
sequencing (Table 3),
[image:3.585.48.542.82.414.2]confirming previous observations (1) and showing the genetic
TABLE 3. Summary of PCR results and IS900
sequencing data obtained for isolates of
M. avium
subsp.
paratuberculosis
types I, II, and III
Isolate Strain
type
Result for: hsp65
coded
SNP(s)eat bp:
F57 DMCa
IS1311b
LSPA
20c
169 216
21P
I
⫹
S
S
f⫺
6
C
G
235G
I
⫹
S
S
f⫺
6
C
G
208G
I
⫹
S
S
f⫺
6
C
G
213G
I
⫹
S
S
f⫺
6
C
G
M189
I
⫹
S
S
f⫺
6
C
G
CAM 63
II
⫹
C
C
⫹
5
C
A
CAM 72
II
⫹
C
C
⫹
5
C
A
464
II
⫹
C
C
⫹
5
C
A
CAM 19
II
⫹
C
C
⫹
5
C
A
CAM 20
II
⫹
C
C
⫹
5
C
A
574
II
⫹
C
C
⫹
5
C
A
896
II
⫹
C
C
⫹
5
C
A
940
II
⫹
C
C
⫹
5
C
A
CAM 78
III
⫹
S
S
⫺
6
C/T
A/G
CAM 40
III
⫹
S
S
⫺
6
C/T
A/G
CAM 38
III
⫹
S
S
⫺
6
C/T
A/G
CAM 86
III
⫹
S
S
⫺
6
C/T
A/G
CAM 87
III
⫹
S
S
⫺
6
C/T
A/G
MI07.01787-2
III
⫹
S
S
⫺
6
C/T
A/G
MI07.04010-2
III
⫹
S
S
⫺
6
T
G
793
III
⫹
S
S
⫺
g⫺
gC/T
A/G
404
III
⫹
S
S
⫺
⫺
gC/T
A/G
408
III
⫹
S
S
⫺
6
C/T
A/G
619
III
⫹
S
S
⫺
6
C/T
A/G
841
III
⫹
S
S
⫺
6
C/T
A/G
634
III
⫹
S
S
⫺
6
C/T
A/G
MI06.00285-2
III
⫹
S
⫺
g⫺
6
C/T
A/G
MI06.00286-2
III
⫹
S
S
⫺
6
C/T
A/G
MI07.06579-2
III
⫹
S
S
⫺
6
C/T
A/G
MI07.06582-2
III
⫹
S
S
⫺
6
C/T
A/G
734
III
⫹
S
S
⫺
6
C/T
A/G
MI05.00897-2
III
⫹
S
S
⫺
6
C/T
A/G
MI06.01981-2
III
⫹
S
⫺
g⫺
6
C/T
G
aPCR analysis of DMC was performed as described by Collins et al. (8). Isolates were identified as S or C strains.
bPCR-REA was performed as described by Marsh et al. (15). Isolates were identified as S or C strains.
cThe PCR method used was developed by Semret et al. (25).
dhsp65sequevar.
eSNPs found in the IS900sequences.
fIS1311data were also confirmed in previous work (29).
gThe DNA template was insufficient and did not yield a visible product in the PCR.
on May 16, 2020 by guest
http://jcm.asm.org/
similarity of these two strain types. However, this report
pro-vides further evidence of additional polymorphic loci that can
be used to distinguish between these two strain types. Also, it
supports the theory that less genomic divergence exists among
type II (bovine) isolates than among type I and III (S) isolates
(4, 26, 31).
The stability and conservation of the IS
900
sequence drift
is reflected at positions 169 and 216 in some copies of IS
900
in type I and III strains from herds in different locations.
These results are consistent with suggestions in previous
studies that
M. avium
subsp.
paratuberculosis
strains tend to
be clonal (34). The IS
900
sequence analysis could be used as
a complementary diagnostic tool for epidemiological
pur-poses to study the geographical distribution patterns of the
three clusters within the
M. avium
subsp.
paratuberculosis
group. On the other hand, from this evidence, it is still not
possible to obtain a correlation between pathogen
adapta-tion to environmental factors or virulence pathways and the
divergences in IS
900
sequences.
Notwithstanding these results,
M. avium
subsp.
paratubercu-losis
nomenclature and the subdivision of strains into groups
are still controversial. Current classifications have been based
on the data gathered by several research groups. It would be
necessary to perform whole-genome sequencing of
M. avium
subsp.
paratuberculosis
types I and III to further clarify the
taxonomy of the lineages.
Nucleotide sequence accession numbers.
The GenBank
ac-cession numbers for IS
900
partial sequences from
M. avium
subsp.
paratuberculosis
type I and type III are FJ775181 and
FJ775182, respectively.
This research was funded by the European Union project
Para-TBTools FP6-2004-FOOD-3B-023106 and the project
AGL2005-07792 of the Spanish Ministry of Science and Technology and by the
Spanish Ministry of the Environment and Rural and Marine Affairs.
E.C. is the recipient of a grant (AP2005-0696 GAN) from the Science
and Innovation Ministry.
We thank the staff of the SADNA (C.I.B. Madrid) for sequencing.
We are also grateful to Devin J. Morey for careful revision of the
manuscript.
REFERENCES
1.Alvarez, J., E. Castellanos, L. de Juan, B. Romero, J. Bezos, S. Rodriguez, A. Aranaz, A. Mateos, and L. Dominguez.2008. Characterization of Mycobac-terium aviumsubsp.paratuberculosisisolates by large sequence
polymor-phisms (LSP) detection andhsp65 sequencing, p. 3–5.InS. S. Nielsen (ed.),
Proceedings of the 9th International Colloquium on Paratuberculosis. Inter-national Association for Paratuberculosis, Madison, WI.
2.Bhide, M., E. Chakurkar, L. Tkacikova, S. Barbuddhe, M. Novak, and I. Mikula.2006. IS900-PCR-based detection and characterization of Mycobac-terium aviumsubsp.paratuberculosisfrom buffy coat of cattle and sheep. Vet.
Microbiol.112:33–41.
3.Bull, T. J., J. Hermon-Taylor, I. Pavlik, F. El-Zaatari, and M. Tizard.2000.
Characterization of IS900loci inMycobacterium aviumsubsp.
paratubercu-losisand development of multiplex PCR typing. Microbiology146:3285. 4.Castellanos, E., A. Aranaz, K. A. Gould, R. Linedale, K. Stevenson, J.
Alvarez, L. Dominguez, L. de Juan, J. Hinds, and T. J. Bull.2009. Discovery
of stable and variable differences in theMycobacterium aviumsubsp.
para-tuberculosistype I, II, and III genomes by pan-genome microarray analysis.
Appl. Environ. Microbiol.75:676–686.
5.Castellanos, E., A. Aranaz, B. Romero, L. de Juan, J. Alvarez, J. Bezos, S. Rodriguez, K. Stevenson, A. Mateos, and L. Dominguez.2007.
Polymor-phisms ingyrAandgyrBgenes amongMycobacterium aviumsubsp.
paratu-berculosistype I, II, and III isolates. J. Clin. Microbiol.45:3439–3442. 6.Castellanos, E., J. Alvarez, A. Aranaz, B. Romero, L. de Juan, J. Bezos, S.
Rodríguez, K. Stevenson, A. Mateos, and L. Domínguez.2008. Use of single
nucleotide polymorphisms in inhA gene to characterize Mycobacterium
aviumsubspeciesparatuberculosisinto types I, II and III, p. 6–8.InS. S.
Nielsen (ed.), Proceedings of the 9th International Colloquium on Paratu-berculosis. International Association for Paratuberculosis, Madison, WI. 7.Coetsier, C., P. Vannuffel, N. Blondeel, J. F. Denef, C. Cocito, and J. L. Gala.
2000. Duplex PCR for differential identification ofMycobacterium bovis,M.
avium, andM. aviumsubsp.paratuberculosisin formalin-fixed
paraffin-em-bedded tissues from cattle. J. Clin. Microbiol.38:3048–3054.
8.Collins, D. M., M. De Zoete, and S. M. Cavaignac.2002.Mycobacterium aviumsubsp.paratuberculosisstrains from cattle and sheep can be distin-guished by a PCR test based on a novel DNA sequence difference. J. Clin.
Microbiol.40:4760–4762.
9.Cousins, D. V., R. Whittington, I. Marsh, A. Masters, R. J. Evans, and P. Kluver.1999. Mycobacteria distinct fromMycobacterium aviumsubsp. para-tuberculosisisolated from the faeces of ruminants possess IS900-like
se-quences detectable by IS900polymerase chain reaction: implications for
diagnosis. Mol. Cell. Probes13:431–442.
10.de Juan, L., J. Alvarez, A. Aranaz, A. Rodriguez, B. Romero, J. Bezos, A. Mateos, and L. Dominguez.2006. Molecular epidemiology of Types I/III
strains of Mycobacterium aviumsubspeciesparatuberculosisisolated from
goats and cattle. Vet. Microbiol.115:102–110.
11.de Juan, L., A. Mateos, L. Dominguez, J. M. Sharp, and K. Stevenson.2005.
Genetic diversity ofMycobacterium aviumsubspeciesparatuberculosis
iso-lates from goats detected by pulsed-field gel electrophoresis. Vet. Microbiol.
106:249–257.
12.Englund, S., G. Bolske, and K. E. Johansson.2002. An IS900-like sequence
found in aMycobacteriumsp. other thanMycobacterium aviumsubsp.
para-tuberculosis.FEMS Microbiol. Lett.209:267–271.
13.Green, E. P., M. L. Tizard, M. T. Moss, J. Thompson, D. J. Winterbourne, J. J. McFadden, and J. Hermon-Taylor.1989. Sequence and characteristics
of IS900, an insertion element identified in a human Crohn’s disease isolate
ofMycobacterium paratuberculosis. Nucleic Acids Res.17:9063–9073. 14.Griffiths, T. A., K. Rioux, and J. De Buck.2008. Sequence polymorphisms in
a surface PPE protein distinguish types I, II, and III ofMycobacterium avium
subsp.paratuberculosis.J. Clin. Microbiol.46:1207–1212.
15.Marsh, I., R. Whittington, and D. Cousins.1999. PCR-restriction
endonu-clease analysis for identification and strain typing ofMycobacterium avium
subsp.paratuberculosisandMycobacterium avium subsp.avium based on
polymorphisms in IS1311.Mol. Cell. Probes13:115–126.
16.Mobius, P., H. Hotzel, A. Rassbach, and H. Kohler.2008. Comparison of 13
single-round and nested PCR assays targeting IS900, ISMav2, f57 and locus
255 for detection ofMycobacterium aviumsubsp.paratuberculosis.Vet.
Mi-crobiol.126:324–333.
17.Mobius, P., G. Luyven, H. Hotzel, and H. Kohler.2008. High genetic
diver-sity amongMycobacterium aviumsubsp.paratuberculosisstrains of German
cattle herds shown by combination of IS900restriction fragment length
polymorphism analysis and mycobacterial interspersed repetitive
unit-vari-able-number tandem-repeat typing. J. Clin. Microbiol.46:972–981.
18.Moss, M. T., E. P. Green, M. L. Tizard, Z. P. Malik, and J. Hermon-Taylor.
1991. Specific detection ofMycobacterium paratuberculosisby DNA
hybridi-sation with a fragment of the insertion element IS900. Gut32:395–398.
19.Pavlik, I., A. Horvathova, L. Dvorska, J. Bartl, P. Svastova, R. du Maine, and I. Rychlik.1999. Standardisation of restriction fragment length
polymor-phism analysis forMycobacterium aviumsubspeciesparatuberculosis. J.
Mi-crobiol. Methods38:155–167.
20.Pickup, R. W., G. Rhodes, S. Arnott, K. Sidi-Boumedine, T. J. Bull, A. Weightman, M. Hurley, and J. Hermon-Taylor.2005.Mycobacterium avium
subsp.paratuberculosisin the catchment area and water of the River Taff in
South Wales, United Kingdom, and its potential relationship to clustering of
Crohn’s disease cases in the city of Cardiff. Appl. Environ. Microbiol.71:
2130–2139.
21.Pickup, R. W., G. Rhodes, T. J. Bull, S. Arnott, K. Sidi-Boumedine, M. Hurley, and J. Hermon-Taylor.2006.Mycobacterium aviumsubsp. paratu-berculosisin lake catchments, in river water abstracted for domestic use, and in effluent from domestic sewage treatment works: diverse opportunities for
environmental cycling and human exposure. Appl. Environ. Microbiol.72:
4067–4077.
22.Poupart, P., M. Coene, H. Van Heuverswyn, and C. Cocito.1993. Preparation
of a specific RNA probe for detection ofMycobacterium paratuberculosisand
diagnosis of Johne’s disease. J. Clin. Microbiol.31:1601–1605.
23.Scanu, A. M., T. J. Bull, S. Cannas, J. D. Sanderson, L. A. Sechi, G. Dettori, S. Zanetti, and J. Hermon-Taylor.2007.Mycobacterium aviumsubspecies
paratuberculosisinfection in cases of irritable bowel syndrome and compar-ison with Crohn’s disease and Johne’s disease: common neural and immune
pathogenicities. J. Clin. Microbiol.45:3883–3890.
24.Semret, M., C. Y. Turenne, and M. A. Behr.2006. Insertion sequence IS900
revisited. J. Clin. Microbiol.44:1081–1083.
25.Semret, M., C. Y. Turenne, P. de Haas, D. M. Collins, and M. A. Behr.2006.
Differentiating host-associated variants ofMycobacterium aviumby PCR for
detection of large sequence polymorphisms. J. Clin. Microbiol.44:881–887.
26.Sevilla, I., J. M. Garrido, M. Geijo, and R. A. Juste.2007. Pulsed-field gel
electrophoresis profile homogeneity ofMycobacterium avium subsp.
paratu-berculosisisolates from cattle and heterogeneity of those from sheep and
goats. BMC Microbiol.7:18.
on May 16, 2020 by guest
http://jcm.asm.org/
27.Sivakumar, P., B. N. Tripathi, and N. Singh.2005. Detection of Mycobac-terium avium subsp. paratuberculosisin intestinal and lymph node tissues of
water buffaloes (Bubalus bubalis) by PCR and bacterial culture. Vet.
Micro-biol.108:263–270.
28.Sohal, J. S., N. Sheoran, K. Narayanasamy, V. Brahmachari, S. Singh, and S. Subodh.2009. Genomic analysis of local isolate ofMycobacterium avium
subspeciesparatuberculosis. Vet. Microbiol.134:375–382.
29.Stevenson, K., V. M. Hughes, L. de Juan, N. F. Inglis, F. Wright, and J. M. Sharp.2002. Molecular characterization of pigmented and nonpigmented
isolates ofMycobacterium aviumsubsp.paratuberculosis.J. Clin. Microbiol.
40:1798–1804.
30.Thibault, V. C., M. Grayon, M. L. Boschiroli, C. Hubbans, P. Overduin, K. Stevenson, M. C. Gutierrez, P. Supply, and F. Biet.2007. New
variable-number tandem-repeat markers for typingMycobacterium aviumsubsp.
para-tuberculosisandM. aviumstrains: comparison with IS900and IS1245
restric-tion fragment length polymorphism typing. J. Clin. Microbiol.45:2404–2410.
31.Turenne, C. Y., D. M. Collins, D. C. Alexander, and M. A. Behr.2008.
Mycobacterium aviumsubsp.paratuberculosisandM. aviumsubsp.aviumare
independently evolved pathogenic clones of a much broader group ofM.
aviumorganisms. J. Bacteriol.190:2479–2487.
32.Turenne, C. Y., M. Semret, D. V. Cousins, D. M. Collins, and M. A. Behr.
2006. Sequencing ofhsp65distinguishes among subsets of theMycobacterium
aviumcomplex. J. Clin. Microbiol.44:433–440.
33.Whipple, D., P. Kapke, and C. Vary.1990. Identification of restriction
frag-ment length polymorphisms in DNA fromMycobacterium paratuberculosis.
J. Clin. Microbiol.28:2561–2564.
34.Whittington, R., I. Marsh, E. Choy, and D. Cousins.1998. Polymorphisms in
IS1311, an insertion sequence common toMycobacterium aviumandM.
avium subsp. paratuberculosis, can be used to distinguish between and within
these species. Mol. Cell. Probes12:349–358.
35.Whittington, R. J., C. A. Taragel, S. Ottaway, I. Marsh, J. Seaman, and V. Fridriksdottir.2001. Molecular epidemiological confirmation and
circum-stances of occurrence of sheep (S) strains ofMycobacterium avium subsp.
paratuberculosisin cases of paratuberculosis in cattle in Australia and sheep
and cattle in Iceland. Vet. Microbiol.79:311–322.
36.Willemsen, P. T. J., D. Bakker, and F. G. van Zijderveld.1999. An error in
the reported IS900nucleotide sequence affects the proposed expression of
ORF2 (hed) and the detection ofM. paratuberculosisusing the polymerase
chain reaction, p. 466–471.InE. J. B. Manning and M. T. Collins (ed.),
Proceedings of the 6th International Colloquium on Paratuberculosis. Inter-national Association for Paratuberculosis, Madison, WI.