Photocontrol of the Accumulation of Plastid Polypeptides during Greening of Tomato Cotyledons'
Potentiation by a Pulse of Red Light
Yael Pauncz, Shimon Gepstein, and BenjaminA. Horwitz*
Department ofBiology, Technion-Israel Institute ofTechnology, Haifa 32000, Israel
ABSTRACT
Apulse of redlight actingthrough phytochrome acceleratesthe formationofchlorophyll uponsubsequenttransferofdark-grown seedlingstocontinuouswhitelight. Specific antibodieswereused tofollow theaccumulationofrepresentative subunits of the major photosyntheticcomplexesduring greeningofseedlingsof tomato (Lycopersicon esculentum). Thetime coursefor accumulation of the varioussubunitswascomparedinseedlings that receivedared light pulse 4 h prior to transfer to continuous white light and parallelcontrols thatdid notreceivea redlight pulse. The light- harvestingchlorophyll-bindingproteins ofphotosystem11(LHC II), the 33-kD extrinsic polypeptide oftheoxygen-evolving complex (OEC1), and subunit11of photosystemI (psaD gene product) all increased in the light, and did so much faster in seedlings that received theinductive red light pulse. The red lightpulse hadno significant effectontheabundanceof thesmall subunit ofribulose 1,5-bisphosphatecarboxylase/oxygenase (Rubisco),noronseveral plastid-encodedpolypeptides: the large subunit of Rubisco,the,B subunitof the CF1complexofplastidATPase, and the43-and47- kDsubunitsofphotosystem11(CP43, CP47). SubunitsI(cytochrome b6f) and IlIl (Rieske Fe-Sprotein) ofthecytochromeb6f complex showed a small or no increase as a resultofthe red pulse. The potentiationofgreeningby apulseofredlight, therefore,isnot expressed uniformly in the abundance of all the photosynthetic complexes andtheir subunits.
Theleaves ofangiosperm seedlingsgrownintotaldarkness contain no Chl, and their plastids differ in structure and composition from mature chloroplasts. The transition from such etioplasts tophotosyntheticallycompetentchloroplasts involves thesynthesis of Chl andmanychloroplastpolypep- tides, and the reorganization of plastidstructure, directed by thenuclearandplastid geneticcompartments(9, 13, 18, 19, 27-29). Light regulates severalaspectsof this greeningproc- ess. In particular, continuous light is required for photore- ductionofPchlide, andapulse of lightactsthrough phyto- chrome to increase the levels of nuclear mRNAs encoding chloroplast proteins (1, 9, 16, 18, 20, 21, 23, 24, 29, 30). Blue light receptors have also been implicated in the control of plastid (5) and nuclear-encoded mRNAs (20, 30).
'Thisworkwassupportedbygrants toB.A.H. from theBat-Sheva deRothschildFoundation,theIsraelMinistry ofAbsorption, and the J.S.FrankfordFoundation(Technion V.P.R.Fund).
Thepolypeptides of thephotosyntheticapparatus,though, are notregulated entirelyatthemRNA level. Plastid-encoded mRNAs are often present in thedark, andcontinuouswhite light regulates their translationorstability,orthe accumula- tion of the protein products (3, 4, 12, 17). Chl stabilizes its apoproteins,whether nuclear orplastid encoded(7, 17). Even forgeneswhose transcription hasclearly beenshown tobe light regulated, additional levelsof control have beenimpli- cated. Transcription rates, mRNA levels, and the rates of polypeptideorChlaccumulationoftendifferintheir kinetics and fluencedependence(16, 24, 29).
A pulse ofR2 reduces or eliminates the lag in Chl accu- mulation that is normally observed when dark-grown an- giosperm seedlings, including those oftomato(Lycopersicon esculentum) (11), are transferred to continuous white light.
This potentiatingeffectofR is ausefulphotobiological tool, because theinductiveactionoflightviaphytochromecanbe separated from thoseprocesses requiring continuousillumi- nation (10). Nevertheless, littleattention has been given to the question of how phytochrome modulation of mRNA levels correspondsto theaccumulation of thepolypeptides.
The abundance of many plastid polypeptides strongly in- creases during greening under continuous white light, whereas othersare present atsignificant levels in etioplasts (12, 13, 19). In this study, we have used antibodies to determine which subunits of the photosynthetic complexes accumulatepreferentiallyinseedlingswhose greeningis ac- celeratedbyaninductivepulse ofR.
MATERIALS AND METHODS PlantMaterialandLightTreatments
Seeds of tomato (Lycopersicon esculentum cv Marymond Rehovot) were purchased from Hazera Seeds Ltd. (Haifa, Israel). Seedlingsweregrown onwater-saturatedvermiculite inatemperature-controlled darkroom at 24 to250Cin 10 x 10 x 10 cm plastic boxes that were placed open in large coveredplastic boxes. The large boxeswerewrappedinblack cloth.Other than the indicatedlight treatments,theseedlings 2Abbreviations: R, red light; LHC II, light-harvesting Chl a/b
binding proteins of PSII; OEC1, 33-kD subunit of the oxygen- evolving complex; FR, far red light; LF, low fluence phytochrome response;VLF,very lowfluence phytochromeresponse.
1934
RED PULSE ACCELERATION OF PLASTID POLYPEPTIDE ACCUMULATION
were grown and handled in total darkness until harvest.
Cotyledons for the 'zero time' (transfer fromdark to white light)points were harvested under dim green light; harvest was in the light for all other timepoints. For protein extrac- tion,cotyledonswerequicklyfrozenwith liquid nitrogen and storedat-800Cuntil use.After6.5 d of growth in the dark, seedlings weretransferredto agrowth chamber (LabLine) at 24 to250C, 80%RH, undercontinuous white light (100,umol m-2 s-' photosynthetically activeradiation) from fluorescent tubes(cool white). TheRandFRlightsources, neutral density and band-pass filters, and fluence rate measurement have been described in detail (11). The standard R pulse was a 100-s exposure giving a totalfluence of 6000,molm-2. FR was given as a high-fluence rate background illumination during and afterthe exposure to R(fordetails,seeFig. 2and ref. 11).
Measurementof Chl Accumulation
Cotyledonswereharvested and put intoglass vials. Total Chl wasmeasured by extractionof intact tissue (15): NN- dimethylformamide was added (1 mL per 50 mg fresh weight), the vialswere keptin thedark at40C for2d, and Chlwasassayed spectrophotometrically.
Immunoblotting
About 50cotyledonpairs(0.2gfresh weight)wereground ina glass-glass homogenizerin 1.5 mLofextraction buffer (100 mm Tris HCl,pH 8.0,5 mm/3-mercaptoethanol, 1 mM PMSF, and 1 mm benzamidine). The extract was filtered through 35-,um monofilament nylon mesh and centrifuged 10min at12,000rpm in aSorvall Microspin 24S. Thepellet (crude membrane fraction) was sonicated for 30 s (setting No.3,microtip, BransonSonifier)in dissociationbuffer(2%
[w/v]SDS,100 mmTris, pH6.8,2%[v/v]f3-mercaptoethanol,
2 mm EDTA, and 10% v/vglycerol) andcentrifuged 5 min at 12,000 rpm as above, and the solubilized proteins were
separatedon a12.5or15%SDS-PAGEinaminigelapparatus (Hoefer, San Francisco, CA). Thesupernatant (solublefrac- tion, 12kS)wasbrought to 10% TCA, kept onicefor 1 h, andcentrifuged10 minat12,000 rpm. TheTCA-precipitated proteins wereresuspended bysonication in a small volume of1 MTrisbase; aftersonication, several volumes ofsample bufferwere added, and the sampleswere centrifugedfor5 minbefore separationbySDS-PAGE.
Immunoblotting methods followed, in general, those de- scribed (6): the polypeptides were blotted to nitrocellulose (Schleicher &Schull, BA85)at400 mAfor2 h. Blockingof nonspecific binding sites was with 0.5% BSA (Fraction V, Sigma)in25 mmTris-HCl, pH7.5, 140 mmNaCl, orlowfat milkdiluted1:20inthesamebuffer, for1hat roomtemper- ature. The blots were incubated overnight at 250C with appropriatedilutions(generally 1:1000) ofthe antisera. The polyclonal antibodies were raised in rabbits against dena- tured, purified subunits ofthe variousphotosynthetic com- plexes (13, 14, 19). Antibody waslocatedon theblots with ProteinA-horseradishperoxidase (Sigma) with diaminoben- zidine as substrate. Protein concentrationsweremeasuredby
theCoomassieblue R spot test with BSA asstandard, and 30 ug of protein were loaded on each lane. Typical blots are illustrated in the figures; all experiments were done on at least three independent sets of seedlings.
RESULTS
PSI'
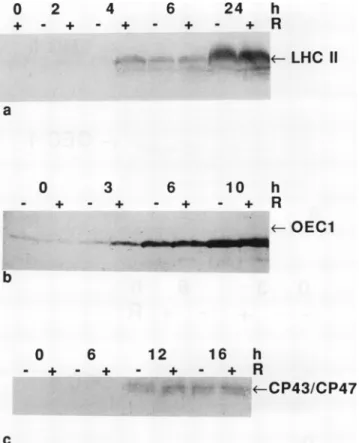
Specific polyclonal antibodies were used to follow the abundance of polypeptidesassociated with PSII in the mem- branefraction(Fig. 1). The LHC II polypeptides were absent inthedark and accumulatedupontransfertolight. The rate of accumulation wasmuch higher in the cotyledonsof seed- lings thatreceived a pulseofR 4 h prior totransferto white light(Fig. la). The33-kDextrinsic protein(OEC1)was pres- ent in thedarkatlow levels and increasedinthe light.An R pulse also potentiatedtherapid accumulation of OEC1in the membrane fraction (Fig. lb). Nosignificant effect of the R pulse was detected when blots of the membrane fractions wereprobedwithantibody to the chloroplast-encoded CP43 andCP47 Chl a proteinsofPSII (Fig. ic). Thedoubleband on immunoblots corresponding to these two proteins in-
2 4 6 24 h
- + - + + - + R
< LHC 11 a
0 3 6 10 h
+ + + - + R
<- OEC1 b
0 6 12 16 h
+ + - + - + R
t tt ~<-- CP43/CP47
C
Figure 1. Immunological detection oftwo nuclear-encoded poly- peptides of PSII during greening, with and without an R pulse pretreatment. +R, R pulse (6000 umol m2) given 4 h prior to transfertowhitelight.-R, No Rpulse.Cotyledonswereharvested after the indicated periodsinwhite light(100 Mmol m-2 S-1 in the 400-700 nmrange). Blots ofgelseparations of30tigof membrane fraction proteinwereprobedwithantisera to:(a)LHCIl;(b)33-kD extrinsic polypeptide(OEC1);and(c)CP43andCP47.
1935
creasedbyonly 6 to 12h in whitelight, whenthe effect of thered pulseon Chl accumulationwas nolongerapparent.
The increase in LHC II accumulation that resulted from the R pulsewas, to a large extent, canceledby FR(Fig. 2a).
Identical blots,whenprobedwithantibodytoOEC1, didnot showFRreversal(Fig. 2a). On blots of total protein extracts, onlyamoderateincrease wasdetectedduringgreening (Fig.
2b). The OEC1 signal was alreadypresent in the dark (Fig.
2b, 0 h in whitelight)athigher levels thaninthemembrane fraction (Fig. lb). Furthermore,the Rpulse hadnoeffect (Fig.
2b),incontrast tothe markedeffect detectedinthemembrane fraction (Fig. lb). The OEC1 was clearly detectable in the supernatantfraction, and,inthisfraction,thepatternoflight regulation (data not shown) was very similar to that found withtotalprotein extracts(Fig. 2b).
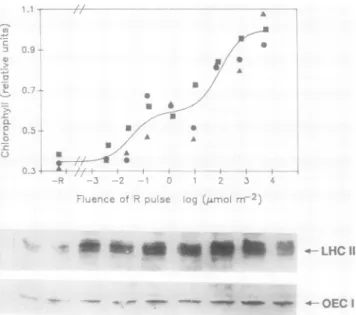
Theabundance ofChl, LHC II, and OEC1 increasedas a function of the fluence of theR pulse (Fig. 3). The fluence dependence forpotentiationof Chl accumulation in tomato seedlingsspans 6orders ofmagnitude andcanbe wellfitby abiphasiccurveconsistingof componentsintheVLFandLF phytochromeresponse ranges (11; Fig. 3). Thelevels ofboth
C: R FR R/FR
e- LHC 11
Figure3. FluencedependenceofChiand oftwoPSIIpolypeptides.
Top panel, Fluence-response curve for the acceleration of Chi accumulation in tomato seedlings byan R pulse. Seedlingswere
given the indicatedRfluences,keptinthe darkfor4h, exposedto
white light for3 h, harvested, and assayed for total ChI content.
Solid line, Nonlinearleast-squares fittothesum oftwosaturation
curves.Differentsymbols indicate differentexperiments;eachsym-
bolrepresentsthevalue foranextractofatleast20cotyledonpairs.
Theimmunoblots shown below the fluence-response curve indi-
catetheabundance ofLHCIIandOEC1 polypeptidesforsamples oftheseedling populations indicated by* in thetop panel. Blots ofmembraneproteinwereprobedasforFigure1.
i- OEC
iLHC II andOEC1 wereincreased byan RpulseintheVLF andLFranges(Fig. 3).
a
(e 3 6 t.
N*- 'Wo
_OEW _W Or- QOEC
b
Figure 2. Photoregulationof two PSII polypeptides.a, FR reversal ofthe effect ofRlightontheabundance of LHC II (top, 24-26kD) andOEC1 (33 kD).C,Control, nolight pulse;R, 100 s at 30jmol m-2s-1;FR,FR light alone,200s at95,umol m-2s-1;R/FR, Rpulse given 20 s after the start of the FR pulse. Blots of 30 gg/lane of membrane fraction protein were probed with each antiserum. b, Immunologicaldetection ofOEC1 insamplesof30yg/laneof total protein extracts.
OtherPhotosynthetic Complexes
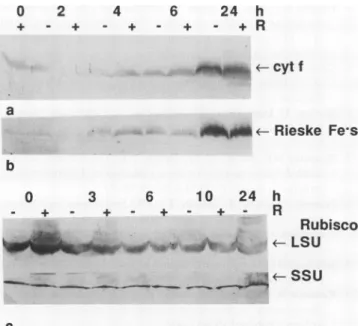
The accumulation ofsubunitII ofPSI (21 kD; psaDgene product), showed a marked potentiation by the R pulse evident at 3 h inFigure4. The aand fi subunits ofchloroplast ATPase (CF1) were probed for on blots of the membrane fraction, and their levels didnotchange during the first hours ofgreening (datanotshown). Representatives oftwo other photosyntheticcomplexes, Cyt b6f and Rubisco,areshownin Figure 5. Subunit I (Cyt f) and subunit III (Rieske Fe-S protein) of the Cyt b6f complex were present at low levels
0
+ - 11 + - + R
of--subunit
Figure 4. Immunologicaldetection of subunit11 of PSIduringgreen- ingwith and without an R pulse pretreatment. Blot of membrane fraction protein, probedwithantiserum tosubunit 11 of PSI(21 kD).
RED PULSE ACCELERATION OF PLASTID POLYPEPTIDE ACCUMULATION
6 10 24 h
+- + - R
Rubisco
4--LSU
; SS U
C
Figure 5. Immunological detection of members of the Cyt b6f
complex and Rubisco.a,Immunoblot of30,ug of membranefraction protein/lane, probed withantiserumto Cytf(subunit ofthe Cyt b6f complex). b, Immunoblotas inpanela, probedwith antiserum tothe Rieske Fe-S protein (subunit III of theCyt b6f complex). Of threereplicate experiments,intwo,the+Rsignalat4hwasslightly higher than the -R signal as shown here, and in the third, the proteinwasnotyetvisibleat4h inwhite light.c, Immunoblots of the soluble fraction (10 Mg/lane) probed with antiseratothe large (firstpanel) and small (second panel)subunits of Rubisco.
during the early hours of greening (Fig. 5, a and b). The signals corresponding to the Rieske Fe-S protein and Cyt f
were not strong enough to state with certainty whether or
notthe Rpulse causedanincrease. BothsubunitsofRubisco
werepresentinthecotyledons of etiolated seedlingsand did notchangeduring the periodstudied(Fig.5c).
DISCUSSION
Representativesof all themajorphotosynthetic complexes
weretested forphytochrome regulationatthe level ofpoly- peptide abundance with the helpofspecific antibodies. An Rpulse givento tomatoseedlings4 hbefore transfertowhite light increased theabundance of the LHC IIduringgreening,
as previously reported for cucumber (2) and tomato (11).
Furthermore, the time course shown in Figure la demon- strates that the rate of accumulation in the first hours of greeningisgreatly enhanced bythe Rpulse. Therateof Chl accumulationin tomatocotyledonsisincreased about 3-fold by anR pulsetreatment4 h priorto transfertowhite light.
The LHC II polypeptides are thus regulated in a manner
similartothat of Chl. ThemostabundantLHCIIpolypeptide (uppermostband visible inFig. la)seems toberesponsible formostof thephotoregulation, and thismightbe theresult
ofphytochrome regulation ofspecific genefamilymembers (seeref.29).
Although the 33-kD extrinsic protein of the OECi does not directly bind Chl, it, too, was more abundant in the membrane fractionfrom cotyledons that received an R pulse (Fig. lb). OEC1 was alreadypresent in the dark (time zero, -R,Fig. lb).LightregulationofOEClintomatodiffers from thatreported for maize (26). In maize, as in tomato, OEC1 waspresent in the dark; a pulse of R increased its abundance, as measured after an additional 48 h in the dark (26). In tomato, an R pulse alone had no effect within the period tested (up to 10 h after the pulse; data not shown), but strongly increased the accumulation of OEC1 in the mem- branefractionduring greening in white light (Fig. lb).
The marked effect of light on the accumulation of OEC1 (Fig. lb) was unexpected, based on results obtained with greeningspinach seedlings(13). Liveanu etal.(13)used total protein samples, whereas the data in Figure lb are for a membranefraction. As shown in Figure 2b, the intensityof the immunoblotsignal obtainedwithtotalproteinincreased only moderately during greening of tomato cotyledons, as reported for spinach (13). Furthermore, the effect of the R pulsewasnotapparent(Fig. 2b).Theeffectofan Rpulseon the abundance of OEC1 in the membrane fractionwas not canceled by irradiation with FR,eventhoughthelevelofthe LHC IIpolypeptides showedclearreversal in the same sam- ples (Fig. 2a). There is considerable evidence from action spectroscopy,photoreversibility, andgenetics (11, 25)thatan R pulse acts through phytochrome to potentiate greening.
The photoregulation of OEC1 differs, though, from that of LHC II.ThefluencedependenceinFigure 3 leavesopenthe possibilitythatOECl levelsinthe membranefraction could be controlledprincipally byR intheVLFrange,anda more detailed quantitative fluence-response study could answer this question. Alternatively, a photoreceptor in addition to phytochrome may be involved in the regulation of OEC1 polypeptide levelsinthethylakoid. It seemslikely that pho- toregulation of the abundance of OEC1 in the membrane fraction is indirect. Sutton et al. (26) found that in maize, OEC1 accumulates underFR light, but doesnotbindtothe innersurface of the thylakoid membrane. Ifthe numberof binding sites iscontrolledby continuouswhitelight andby theRpulse,suchamodel couldexplain ourfindings.There is nodirect evidencethat an Rpulseincreasesthenumber of PSIIreactioncenters,butthis remainstobe tested under the conditions usedhere.
Subunit II of PSI increased in white light (Fig. 4), as reported forrice(8),oat,bean, andspinach (19).The Rpulse stronglyincreased the abundanceofthis subunit(Fig. 4). Like the33-kD extrinsicproteinofPSII(OECi),subunit IIofPSI is a nuclear-encoded, peripheral membraneprotein (22). In contrast, the chloroplast-encodedpsaA and psaBgeneprod- ucts inbarley increasedstronglyinthelight,withnoevidence ofphytochromeregulation (12).
TwomembersoftheCytb6f complex, CytfandtheRieske Fe-Sprotein(Fig. 5,aandb), increasedinabundanceduring greening in white light. In spinach, both ofthese subunits were already present in etiolated seedlings and did not in- creaseduring greening (19). The differencebetweentomato andspinachmightreflect the fact thatweanalyzed amem- branefraction(Fig. 5a),whereas thespinachdataarefor total protein (19). In tomato, subunit I (Cyt f) and subunit III
0 2
+ - +
4 6 24 h
- + - + - R
_dommal__e'iw <-- cytf
a
__h*- Rieske Fe-s
b
0 3
- +1 - +
1937
(Rieske Fe-S protein) wereclearly detectableat20h inwhite light. Thelargeand small subunits of Rubiscowere notlight regulated (Fig. 5c), as also reported for spinach (19). In mustardcotyledons, Rubisco activity (23, 24) aswell asthe immunochemically detectable levels(23) ofthesubunits are light regulated. Asidefromthe obviouspossiblityof species differences, one must note the temporal complexity of the control of gene expression in seedlings. Only one set of conditions (time after sowing, time between R pulse, and transfer to white light) was used in this study. The time courseofcompetence(23)torespondtolightmaydiffer from one gene to another, and certainly from one species to another.
An R pulse potentiates the rapidaccumulation of Chl in tomatocotyledons. This rapid greening is accompaniednot onlybyarapid accumulation of LHC II,amajorChlapopro- tein,but alsooftwoperipheralthylakoid membrane proteins that do not bind Chl: the 33-kD polypeptide (OECl) and subunit IIofPSI. Other subunitsstudiedwereeitheralready present in thedarkorpresentonlyatverylow levelsthrough- out the first 4 h of greening, when the potentiation effect was mostevident.It isthus clear thatpotentiationofgreening byan Rpulseisnotuniformlyexpressedinall thephotosyn- thetic subunits.
It will be important to determine whether the class of nucleargenes whosetranscription isphytochrome regulated always coincideswith theclass whose polypeptideproducts accumulate preferentially in seedlings that received an R pulse. One gene (family) that belongs to both classes in tomatoseedlingsiscab. ThemRNAsfor theother twopoly- peptides whoselevels wereclearly increased bythe Rpulse, OEC1 and subunitII, are alsoincreasedbya pulse of R; in spinach, psaDmRNA wasinducedbyRandreversedby FR (1), and in tomato, the level of message for OEC1 was likewisephotoreversible(21).Particularly for OEC1, the pho- toregulation of polypeptide abundance cannotbeexplained bymRNAregulationalone: OEC1mRNA isphotoreversibly inducedby phytochrome(21),whereas thepolypeptidelevel is not photoreversible under the same conditions (Fig. 2a), andphotoregulationwasevidentinmembranefractions but not total protein (Fig. 2b). The pronounced effect of the R pulse on accumulation of OEC1 in the membrane fraction (Fig. lb)musthave anotherexplanation.
LITERATURE CITED
1. BrunnerH, ThummlerF,Song G,RudigerW(1991)Phyto- chrome-dependent mRNA accumulation for nuclear coded photosystem I subunits in spinach seedlings. J Photochem PhotobiolB 11:129-138
2. CohenL, ArzeeT,ZilbersteinA(1988) Mimicry by cytokinin ofphytochrome-regulated inhibition of chloroplast develop- ment in etiolated cucumber cotyledons. Physiol Plant 72:
57-64
3. Deng XW, TonkynJC, PeterGF, ThornberJP,Gruissem W (1989) Post-transcriptional control ofplastidmessenger RNA accumulation during adaptation ofchloroplasts to different light qualityenvironments.Plant Cell1:645-654
4. Fromm H,DevicM, Fluhr R, Edelman M (1985) Controlof psbAgeneexpression:In maturechloroplasts,lightregulation of 32 kDaprotein synthesisinindependentoftranscriptlevel.
EMBOJ4:291-295
5. GresselJ(1978)Light requirementsfor theenhancedsynthesis of a plastid mRNA during Spirodela greening. Photochem Photobiol 27: 167-169
6. Harlow E, Lane D (1988) Antibodies: A LaboratoryManual.
Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY
7. Harpster MH, Mayfield SP, Taylor WC (1984) Effects ofpig- ment-deficient mutants on theaccumulationofphotosynthetic proteinsinmaize.PlantMol Biol3:59-71
8. IwasakiY, Sasaki T, Takabe T (1990)Sequencing and expres- sionof thegene that encodesa20-kDapolypeptide of thePSI complex from cucumber cotyledon. Plant Cell Physiol 31:
871-879
9. Jenkins GI (1988) Photoregulation of gene expression in plants.
Photochem Photobiol48:821-832
10. KasemirH, Oberdorfer U, MohrH(1973)Atwofoldactionof phytochromeincontrollingchlorophyllaaccumulation. Pho- tochemPhotobiol18:481-486
11. Ken-Dror S,Horwitz BA (1990) Alteredphytochrome regula- tionofgreeningin anaurea mutantoftomato.PlantPhysiol 92:1004-1008
12. Laing W, Kreuz K, Apel K (1988) Light-dependent, but phyto- chrome-independent, translational control of the accumula- tion of the P700 chlorophyll-a protein of photosystem I in barley (Hordeum vulgareL.).Planta176:269-276
13. Liveanu V,Yocum CF, Nelson N (1986)Polypeptidesof the oxygen-evolving photosystem II complex. Immunologicalde- tectionandbiogenesis. J Biol Chem261:5296-5300
14. LivneA, Gepstein S(1988)Abundanceof themajorchloroplast polypeptides during development and ripening of tomato fruits. PlantPhysiol 87: 239-243
15. MoranR,PorathD(1980)Chlorophyll determinationinintact tissues using N,N-dimethylformamide. Plant Physiol 65:
478-479
16. Mosinger E, Batschauer A, Apel K, Schifer E, Briggs WR (1988)Phytochrome regulation ofgreening inbarley.Effects onmRNA abundance andontranscriptionalactivity ofiso- lated nuclei. Plant Physiol86: 706-710
17. Mullet JE,Klein PG, Klein RR (1990) Chlorophyll regulates accumulationof theplastid-encoded chlorophyllapoproteins CP43 and Dl by increasing apoprotein stability. Proc Natl AcadSciUSA87:4038-4042
18. NagyF,Kay SA,ChuaNH (1988) Gene regulationbyphyto- chrome. Trends Genet4:37-42
19. Nechushtai R, Nelson N (1985) Biogenesis of photosystem I reaction center during greening of oat, bean and spinach leaves. Plant MolBiol 4:377-384
20. OelmullerR,KendrickRE,BriggsWR (1989)Blue-lightme- diated accumulation ofnuclear-encodedtranscriptscoding for proteins of the thylakoid membraneis absent in the labile phytochrome-deficientaureamutantoftomato.PlantMol Biol 13:223-232
21. Palomares R, Herrmann RG, Oelmiuler R (1991) Different blue-lightrequirementfortheaccumulationoftranscriptsfrom nucleargenesfor thylakoidproteins in Nicotianatabacumand Lycopersicon esculentum. J Photochem Photobiol B 11:
151-162
22. Scheller HV, M0ller BL (1990) Photosystem I polypeptides.
PhysiolPlant 78: 484-494
23. Schmidt S, Drumm-Herrel H, Oelmuiller R,Mohr H (1987) Time course of competence in phytochrome-controlled ap-
RED PULSE ACCELERATION OF PLASTID POLYPEPTIDE ACCUMULATION
pearanceofnuclear-encoded plastidicproteinsandmessenger
RNAs. Planta170:400-407
24. SchusterC, Oelmiiller R, Mohr H (1988) Controlby phyto- chrome of theappearance ofribulose-1,5-bisphosphate car-
boxylase and the mRNA for its small subunit. Planta 174:
426-432
25. Sharrock RA,ParksBM,KoornneefM, QuailPH(1988)Mo- lecular analysis of the phytochrome deficiency in an aurea mutantoftomato.Mol GenGenet213: 9-14
26. Sutton A, SieburthLE,Bennett J(1987) Light-dependentac-
cumulation and localization of photosystem II proteins in maize. EurJBiochem164:571-578
27. Takabe T (1986) Accumulation of chloroplast proteinsduring greeningofriceleaves. Plant Sci43: 193-199
28. Taylor WC(1989)Regulatoryinteractionsbetweennuclearand plastid genomes. Annu Rev Plant Physiol Mol Biol 40:
211-233
29. Thompson WF, Kaufman LS, Watson JC (1985) Induction of plantgeneexpressionby light. Bioessays3:153-159 30. Wehmeyer B, Cashmore AR, Schafer E (1990) Photocontrol of
the expression of genes encoding chlorophyll a/b binding proteinsand smallsubunitofribulose-1,5-bisphosphate car-
boxylaseinetiolated seedlings ofLycopersicon esculentum(L.) andNicotiana tabacum(L.). PlantPhysiol93:990-997
1939