Work dependent deactivation of a crustacean muscle
15
0
0
Full text
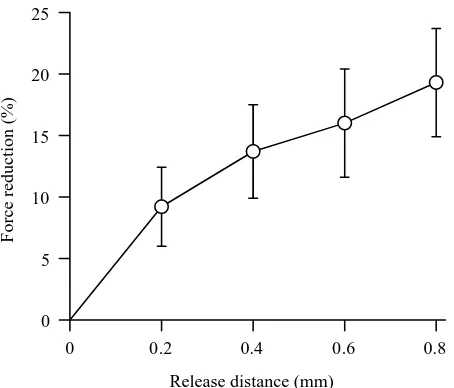
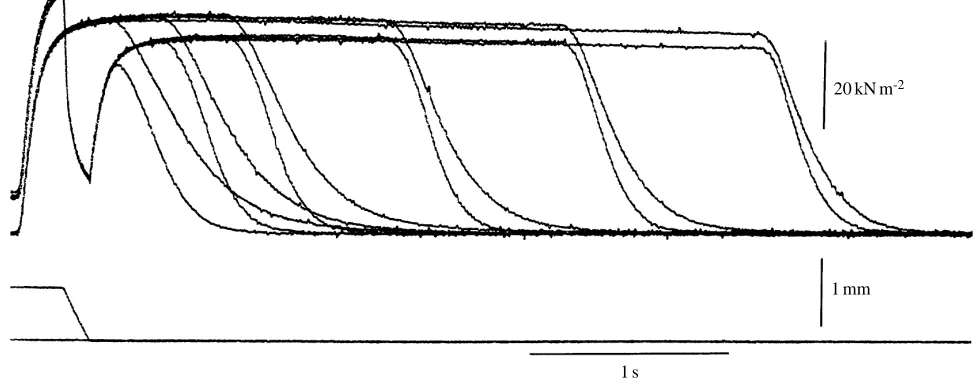
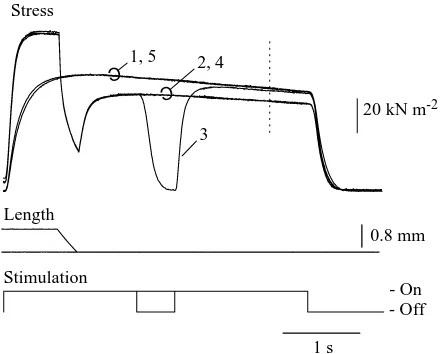
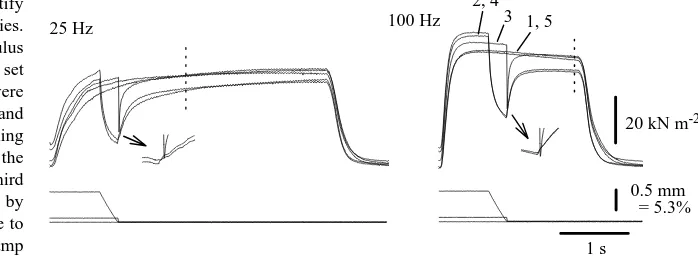
Figure



+7



Related documents
