Certainly the most consistent biochemical response of poikilothermic organisms to environmental cooling is an increase in fatty acid unsaturation of both membrane and depot lipids (Cossins, 1994; Hazel and Williams, 1990). This response has been observed over two distinctive time scales; first, when organisms are conditioned over days and weeks to the cold or warm and, second, when comparing species inhabiting extreme cold and warm thermal environments. The first is a clear short-term phenotypic response to altered temperature thought to be linked to seasonally adjusted thermal resistance (‘resistance acclimation’) of the whole organism. The second occurs over evolutionary time and again has been linked with adaptation of thermal resistance limits and physiological performance to specific thermal habitats.
This compositional response is generally interpreted as causing a disordering of the bilayer to offset the cold-induced ordering. It is thus compensatory in conserving a particular membrane physical condition in the face of thermally induced
disturbance, as reflected in the widely used term homeoviscous adaptation (Cossins, 1994). Although there is some uncertainty as to the precise nature of the conserved membrane condition (Cossins, 1994; Hazel, 1995), biophysical techniques, such as fluorescence polarisation spectroscopy, provide a useful semi-quantitative estimate of the extent to which biophysical structure is conserved, a parameter termed homeoviscous efficacy (HE) (Cossins and Raynard, 1987). Studies on membranes isolated from a range of tissues of thermally acclimated fish have shown HEs ranging from zero (i.e. no response in muscle sarcoplasmic reticulum; Cossins et al., 1978) to 100 %, when biophysical properties are conserved at each of the different acclimation temperatures (Crockett and Hazel, 1995; Lee and Cossins, 1990). Most preparations possessed HEs of 30–50 % (Cossins and Raynard, 1987).
With regard to interspecific comparisons, Cossins and Prosser (1977) examined brain synaptosomal membranes from an Arctic, a temperate and a warm-adapted desert spring JEB2742
Teleost species from cold environments possess more disordered brain synaptic membranes than species from warm habitats, thereby providing equivalent physical structures at their respective habitat temperatures. We have related this adaptive interspecific biophysical response to the fatty acid composition of brain membranes from 17 teleost species obtained from Antarctic, temperate and semi-tropical waters, as well as from rat and turkey as representative homeotherms. Cold-adaptive increases in membrane disorder (determined by fluorescence anisotropy with diphenylhexatriene as probe) were correlated with large and linear increases in the proportion of unsaturated fatty acids, from 35 to 60 % in phosphatidylcholine (PtdCho) and from 55 to 85 % in phosphatidylethanolamine (PtdEth). For PtdCho, the cold-adaptive increase in unsaturation was associated almost entirely with increased proportions (from 7 to 40 %) of polyunsaturated fatty acids (PUFAs), with mono-unsaturates (MUFAs) providing an approximately
constant proportion in all species. Exactly opposite effects were evident for phosphatidylethanolamine (PtdEth). Thus, the compositional adaptation for PtdCho occurred largely by exchange of polyunsaturated and mono-unsaturated fatty acid in the sn-2 position, whilst for PtdEth it involved exchanges between saturates and mono-unsaturates at the sn-1 position. This difference may be related to the different molecular shapes of the two phosphoglycerides and the need to maintain the balance between bilayer-stabilising and -destabilising tendencies. This comparative study provides a more comprehensive view of the compositional adjustments that accompany and perhaps account for temperature-adaptive interspecific differences in membrane physical structure.
Key words: temperature adaptation, brain, membrane, fluorescence anisotropy, DPH, phosphatidylcholine, phosphatidylethanolamine, fatty acid unsaturation, molecular species.
Summary
Introduction
LIPID COMPOSITIONAL CORRELATES OF TEMPERATURE-ADAPTIVE
INTERSPECIFIC DIFFERENCES IN MEMBRANE PHYSICAL STRUCTURE
JAMES A. LOGUE1, ART L. DE VRIES2, ELFRIEDA FODOR1 ANDANDREW R. COSSINS1,* 1Integrative Biology Research Division, School of Biological Sciences, University of Liverpool, PO Box 147,
Liverpool L69 3BX, UK and 2Department of Molecular and Integrative Physiology, University of Illinois, Urbana, IL 61801, USA
species of teleost fish and demonstrated 100 % HE for particular species pairs. This was extended to purified brain synaptic membranes by Behan-Martin et al. (1993), who showed 70–100 % HE, depending on the spectroscopic probe employed, for a somewhat wider range of species including, for the first time, an Antarctic teleost, Notothenia neglecta. This response was considerably greater than that observed in corresponding membranes in thermally acclimated goldfish suggesting that, in this respect at least, the evolutionary adaptation of a species greatly exceeds the potential for adaptation of any individual.
The compositional basis of interspecific adaptation has been linked to membrane unsaturation (Cossins and Prosser, 1977; Farkas and Roy, 1989), although little exact detail is given beyond this general statement. Here, we provide a detailed and more precise understanding of how lipid compositional adjustments underpin the conservation of membrane physical structure over the full range of temperatures experienced by vertebrate animals. Brain synaptic preparations have been isolated from 17 teleost species obtained from Antarctic, temperate and tropical habitats and also from a representative species of a bird and a mammal. We correlate differences between species in membrane biophysical structure with differences in lipid fatty acid composition and demonstrate positionally specific but opposing changes in the two major phosphoglyceride classes.
Materials and methods
Animals
Table 1 lists the species used in this work. Where possible, fish were maintained at the temperature specified for at least 21 days. In Liverpool, the tilapia, striped bass and rainbow trout were fed to satiety once daily with standard trout pellets (Trouw, Longridge, Preston, UK). Gourami were fed once daily with tropical fish food (Tetramin, Tetra Werke, Melle, Germany). Wild-caught fish from Antarctic and Florida were held for a few days, during which time they were unfed. Specimens of the Antarctic Pagothenia borchgrevinki were acclimated at 4 °C for 17 days, during which time they were unfed. Black cod were caught in New Zealand and transported by air freight to the US Antarctic Research Program base at McMurdo Sound.
Isolation of brain synaptic membranes
The method was modified from that described previously (Behan-Martin et al., 1993; Cossins and Prosser, 1982). The lysed synaptosomal pellet was resuspended by gentle homogenisation in 2.5 ml of 10 mmol l−1 imidazole, pH 7.4, and layered on a discontinuous sucrose gradient consisting of equal volumes of 0.9 and 1.0 mol l−1 sucrose in 10 mmol l−1 imidazole, pH 7.4. After centrifugation at 100 000 g for 120 min, the material occupying the interface was removed, diluted 10-fold with lysing medium and pelleted at 100 000 g for 60 min. This pellet was resuspended by homogenisation (all-glass homogeniser) in a small volume of lysing medium
and either used immediately or stored frozen at −80 °C. Preliminary experiments demonstrated that storage over several months has no effect upon the values or temperature-dependence of 1,6-diphenyl-1,3,5-hexatriene (DPH) anisotropy. Previous studies have shown that these membrane fractions are of synaptosomal origin, as judged from marker enzyme assays, and have negligible mitochondrial contamination (Behan-Martin et al., 1993).
Lipid extraction, phospholipid fractionation and fatty acid analysis
Extraction of a total lipid fraction, separation of the major phospholipid headgroup classes and transmethylation were performed as described previously (Lee and Cossins, 1990). Fatty acid methyl esters were separated on a gas–liquid chromatograph (Series 610, ATI Unicam, Cambridge, UK) equipped with a fused silica, free fatty acid phase capillary column (30 m×0.25 mm, J&W Scientific, PhaseSep, Queensferry, Clwyd, UK). Methyl esters were identified by comparing peak retention times with authentic standards whose identity had been confirmed by mass spectrometry. Fatty acid compositions were calculated on a percentage by mass basis directly from the chromatographic output using data-analysis software from ATI Unicam.
Molecular species analysis by high-performance liquid chromatography
Phospholipid molecular species composition was determined following a published procedure (Takamura and Kito, 1991). High-performance liquid chromatography (HPLC) was performed on a PU4100 liquid chromatograph equipped with a PU4110 ultraviolet/visible detector (ATI Unicam, Cambridge, UK) using a reverse-phase octadecylsilyl (ODS) column (Supercosil, 5µm, 4.6 mm i.d. × 25 cm, Supelco, Suplechem UK Ltd) and an acetonitrile:2-propanol (80:20, v/v, HPLC grade) solvent system at a flow rate of 1 ml min−1. The detection wavelength was 254 nm. Peaks were identified by reference to retention times of authentic standards and published data (Bell and Dick, 1991; Takamura and Kito, 1991).
Fluorescence anisotropy
Membrane physical properties were determined by measurements of fluorescence anisotropy using 1,6-diphenyl-1,3,5-hexatriene (DPH) as probe. We used the T-format polarisation fluorometer described previously (Cossins and Macdonald, 1986) or a PC1 spectrofluorometer (ISS Inc, Urbana, Illinois, USA), both giving identical results.
Results
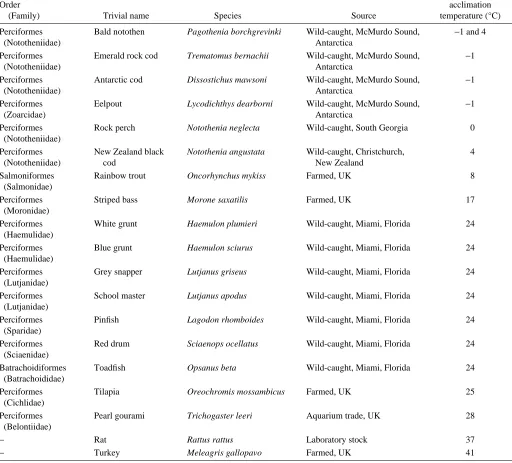
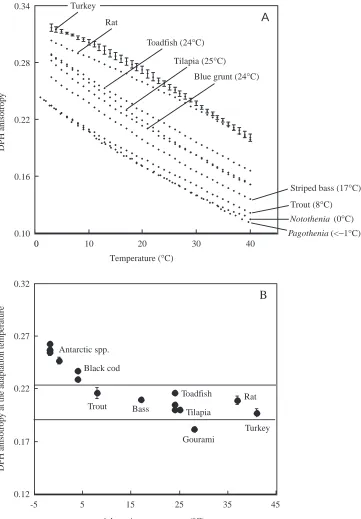
inference greater disorder, than those from temperate species over the full range of measurement temperatures. Temperate species possessed more disordered membranes than semi-tropical species and these in turn showed greater disorder than the membranes of the representative mammal and bird. Thus, a clear correlation exists between synaptic membrane order and species adaptation temperature. Measurement of anisotropy at each of the respective adaptation temperatures allows a direct comparison among species (Fig. 1B). With the exception of the Antarctic species and black cod, values for all the fish, avian and mammalian species were nearly identical, although the value for the pearl gourami was somewhat lower. This indicates that for these species the differences in anisotropy shown in Fig. 1A were sufficient to offset completely the direct
effects of temperature upon anisotropy. The Antarctic species showed somewhat higher anisotropies indicating that they were not sufficiently disordered to provide the same anisotropy as all the other species.
[image:3.609.55.567.100.563.2]The fatty acid composition of the two major phospholipid headgroup classes, phosphatidylcholine (PtdCho) and phosphatidylethanolamine (PtdEth), were determined in replicate membrane preparations for each species. The major fatty acids were 16:0, 18:0, 18:1n-9, 20:4n-6, 22:4n-6 and 22:6n-3. Phosphatidylethanolamine also contained appreciable quantities of alkenyl chains, predominantly alk16:0 and alk18:0 together with alk18:1n-7 and alk18:1n-9 as minor components. Fig. 2 shows that the proportion of unsaturated fatty acids for each phosphoglyceride class was inversely and Table 1. List of species
Adaptation/
Order acclimation
(Family) Trivial name Species Source temperature (°C)
Perciformes Bald notothen Pagothenia borchgrevinki Wild-caught, McMurdo Sound, −1 and 4
(Nototheniidae) Antarctica
Perciformes Emerald rock cod Trematomus bernachii Wild-caught, McMurdo Sound, −1
(Nototheniidae) Antarctica
Perciformes Antarctic cod Dissostichus mawsoni Wild-caught, McMurdo Sound, −1
(Nototheniidae) Antarctica
Perciformes Eelpout Lycodichthys dearborni Wild-caught, McMurdo Sound, −1
(Zoarcidae) Antarctica
Perciformes Rock perch Notothenia neglecta Wild-caught, South Georgia 0 (Nototheniidae)
Perciformes New Zealand black Notothenia angustata Wild-caught, Christchurch, 4
(Nototheniidae) cod New Zealand
Salmoniformes Rainbow trout Oncorhynchus mykiss Farmed, UK 8 (Salmonidae)
Perciformes Striped bass Morone saxatilis Farmed, UK 17
(Moronidae)
Perciformes White grunt Haemulon plumieri Wild-caught, Miami, Florida 24 (Haemulidae)
Perciformes Blue grunt Haemulon sciurus Wild-caught, Miami, Florida 24 (Haemulidae)
Perciformes Grey snapper Lutjanus griseus Wild-caught, Miami, Florida 24 (Lutjanidae)
Perciformes School master Lutjanus apodus Wild-caught, Miami, Florida 24 (Lutjanidae)
Perciformes Pinfish Lagodon rhomboides Wild-caught, Miami, Florida 24 (Sparidae)
Perciformes Red drum Sciaenops ocellatus Wild-caught, Miami, Florida 24 (Sciaenidae)
Batrachoidiformes Toadfish Opsanus beta Wild-caught, Miami, Florida 24 (Batrachoididae)
Perciformes Tilapia Oreochromis mossambicus Farmed, UK 25
(Cichlidae)
Perciformes Pearl gourami Trichogaster leeri Aquarium trade, UK 28 (Belontiidae)
− Rat Rattus rattus Laboratory stock 37
linearly related to species adaptation temperature. Thus, the Antarctic species possessed the greatest degree of phospholipid unsaturation, whereas synaptic membranes from the representative homeotherms contained the lowest proportions of unsaturated fatty acids.
Thus, adaptation to cold is associated with increased unsaturation and physical disordering of brain synaptic membranes. Fig. 3 shows how these two variables are related. The unsaturation index (UI) is a measure of the number of double bonds within a sample and is calculated as the sum of
the percentage of each unsaturated fatty acid multiplied by the number of double bonds within that fatty acid. It can be seen that a linear correlation exists between increasing bilayer unsaturation and decreasing anisotropy, i.e. increasing membrane disorder.
Increases in the proportion of unsaturated fatty acids incorporated into membrane phosphoglycerides was also associated with changes in the types of unsaturated fatty acids. Fig. 4 shows how the unsaturation index (UI) varies with the increasing proportion of unsaturated fatty acids. The UI
40 30
20 10
0 0 0.10 0.16 0.22 0.28 0.34
Temperature (°C)
A
Blue grunt (24°C) Turkey
Rat
Tilapia (25°C) Toadfish (24°C)
Striped bass (17°C) Trout (8°C) Notothenia (0°C) Pagothenia (<−1°C)
DPH anisotrop
y
Adaptation temperature (°C)
Turkey Rat Tilapia
Gourami Toadfish Bass
Trout Antarctic spp.
Black cod
Β
45 35
25 15
5 -5
0.12 0.17 0.22 0.27 0.32
DPH anisotrop
[image:4.609.199.560.72.591.2]y at the adaptation temperature
increases in a clear and linear fashion for PtdCho (Fig. 4A). The dashed lines plot the values of UI if all unsaturated fatty acids had one unsaturation bond (n=1), two bonds (n=2) and so on. The line connecting the data points for different species crosses these transects, indicating an increasing number of unsaturation bonds per unsaturated fatty acid. The values for rat and turkey fell between the isopleths for mono- and di-unsaturates, i.e. an average of 1–2 double bonds per unsaturated fatty acid. The values for the tropical marine fish all fell between di- and tri-unsaturates, and the Antarctic species clustered at approximately four unsaturates. Thus, the increasing proportion of unsaturated fatty acids of cold-adapted species is linked with linearly increasing average numbers of unsaturation bonds for each unsaturated fatty acid. The relationship is distinctly different for PtdEth (Fig. 4B). Here, despite the increase in percentage unsaturation with reducing adaptation temperature, the average number of unsaturation bonds per unsaturated fatty acid for rat, turkey and gourami (4–5 double bonds) was similar to that of trout and the Antarctic species. Values for semi-tropical species were slightly higher.
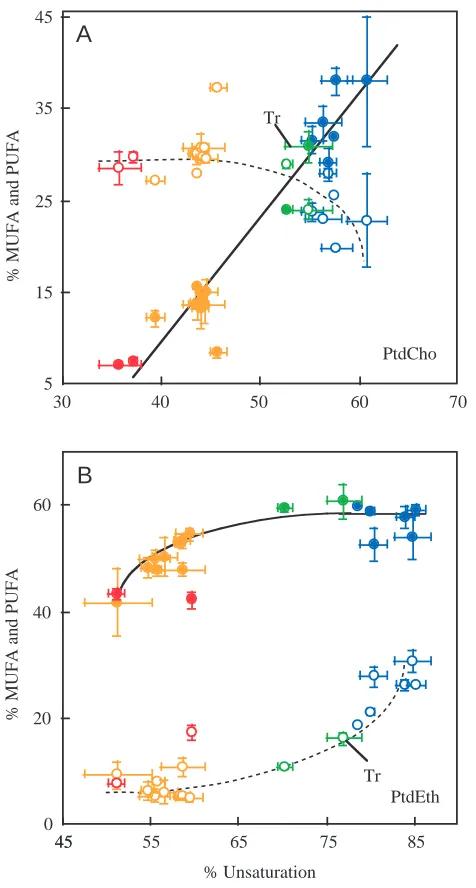
We have sought to separate the contribution of mono-(MUFAs) and polyunsaturated fatty acids (PUFAs) to the interspecific variation in fatty acid composition. For PtdCho (Fig. 5A), the PUFAs showed large increases from 7–8 % in rat and turkey and 12–15 % in tropical fish and to 35 % in the Antarctic fish. In contrast, MUFAs showed very small changes; with all species except gourami possessing 20–30 %. For PtdEth (Fig. 5B), the trends were quite different. For the teleost species, there were comparatively large differences in MUFAs, from 5–10 % in tropical and semi-tropical species, to 10–16 % in temperate and to 25–30 % in Antarctic species. Conversely,
there were only small changes in PUFAs which ranged from 45–55 % for tropical species to 50–60 % for Antarctic species. Thus, the temperature-related variation in the proportion of unsaturated fatty acids was shown to be lipid-specific, with cold adaptation associated with increases in PUFAs in PtdCho but increases in MUFAs in PtdEth.
Polyunsaturated fatty acids tend to occupy the sn-2 position whilst saturates and mono-unsaturates occupy the sn-1 position, and we have therefore explored the interspecific variation in positional specificity. Fig. 6 and Table 2 compare the molecular species composition for both phosphoglyceride classes for membranes from the Antarctic species Pagothenia
borchgrevinki and the rat. For PtdCho (Fig. 6A), the sn-1
position was largely occupied by saturates, although there was a slightly greater proportion of mono-unsaturates in P.
borchgrevinki. In the sn-2 position, the rat possessed
mainly saturates (SFAs) and mono-unsaturates whilst P.
Adaptation temperature (°C) Antarctic spp.
Trout Bass
Tropical spp. Rat Turkey Gourami
45 35
25 15
5 -5
30 50 70 90
%U
n
sa
tu
ra
ti
o
[image:5.609.64.280.69.267.2]n
Fig. 2. The relationship between adaptation temperature and percentage unsaturated fatty acids in phosphatidylethanolamine (open symbols) and phosphatidylcholine (filled symbols) of brain synaptic membrane fractions prepared from the different species. Each data point represents a separate species (mean ± S.D., N=3 separate preparations). Lines were fitted by the method of least squares; PtdCho, r2=0.93, P<0.0001; PtdEth, r2=0.89, P<0.0001.
Table 2. Molecular species analysis of phosphatidylcholine
(PtdCho) and phosphatidylethanolamine (PtdEth) from brain synaptic membranes of rat and Pagothenia borchgrevinki
Diacyl
Mass %
species PtdCho PtdEth
sn-1/sn-2 Rat P. borchgrevinki Rat P. borchgrevinki
16:1/22:6 4.4
16:1/20:4 1.0
18:1/20:5 0.9 9.1
16:0/20:5 6.1
18:1/22:6 3.3 39.2
16:0/22:6 4.1 51.5 12.3 23.8
18:1/22:5 5.2
16:0/22:5 1.4
18:1/20:4 3.8 1.4 1.3
16:0/20:4 0.9 1.1
16:1/16:1 1.1
18:0/20:5 7.8
16:1/18:1 15.8
18:0/22:6 1.8 2.9 37.6 12.2
16:0/18:2 1.6 14:0/16:0 1.1
20:1/20:4 0.8
18:0/20:4 2.7 1.3 18.4
18:1/18:1 1.4 9.2
16:0/18:1 42.6 18.8 3.2
16:0/16:0 25.7 2.5
18:0/22:4 2.5
18:0/18:1 8.8 2.9
18:0/16:0 6.4
SFA/SFA 33.2 0 2.5 0
SFA/MUFA 51.4 18.8 6.1 0
SFA/PUFA 10.2 62.6 72.3 44.9
MUFA/MUFA 1.4 16.9 9.2 0
MUFA/PUFA 3.8 1.8 9.9 55.1
[image:5.609.317.565.112.489.2]borchgrevinki possessed mainly PUFAs with a smaller
proportion of MUFAs. Thus, rat PtdCho is composed mainly of disaturates (33.2 %) and SFA/MUFA molecular species (51.4 %) (Table 2), compared with SFA/MUFA (18.8 %) and SFA/PUFA (62.6 %) for P. borchgrevinki. Regarding PtdEth (Fig. 6B), the sn-1 position was composed largely of saturates in rat but split between saturates and MUFAs in P.
borchgrevinki. The sn-2 position is entirely made up of PUFAs
for the Antarctic teleost and over 80 % for rat, the remainder being MUFAs. Rat PtdEth is therefore composed primarily of SFAs/PUFAs (72.3 %) and the Antarctic fish PtdEth of similar proportions of SFAs/PUFAs (44.9 %) and MUFAs/PUFAs (55.1 %). Consequently, for PtdCho, the differences between the rat and the Antarctic teleost lay in the composition of the sn-2 position (increased PUFA in P. borchgrevinki), whilst in PtdEth it occurred mainly in the sn-1 position (increased MUFA in P.
borchgrevinki). These differences were largely due to variation
in specific molecular species. Fig. 7A shows that, for PtdCho,
P. borchgrevinki possessed reduced proportions of 16:0/18:1
molecular species but increased proportions of 16:0/22:6. In contrast, for PtdEth, the difference between species was due largely to a decrease in 18:0/22:6 in the Antarctic species and a corresponding increase in 18:1/22:6 (Fig. 7B).
Discussion
Previous interspecific analyses of thermoadaptation of brain
membranes have been limited to one species from each of the principal thermal environments (Behan-Martin et al., 1993; Cossins and Prosser, 1977), and the conclusions drawn have obviously been dependent upon the representative status of each species. Thus, a particularly important feature of the analysis reported here is the inclusion of multiple species from each thermal environment including five species from Antarctica and nine from warm semi-tropical waters. This tests in a much more comprehensive manner the degree to which
DPH anisotropy at 20°C
0.28 0.24
0.20 0.16
0 100 200 300 400
Tr
[image:6.609.318.551.72.478.2]Unsaturation index, UI
Fig. 3. The relationship between 1,6-diphenyl-1,3,5-hexatriene (DPH) anisotropy of brain synaptic membranes from a range of vertebrate species and the unsaturation index (UI) of the major phospholipid classes phosphatidylethanolamine (open symbols) and phosphatidylcholine (filled symbols). Anisotropy was measured at 20 °C. Species are divided into four groups; Antarctic (blue), temperate (green) and tropical (orange) fish species and homeotherms (red symbols). Each data point represents a separate species (mean ±S.D., N=3 separate preparations). The value for trout is indicated by ‘Tr’. Lines were fitted by the method of least squares; PtdCho, r2=0.87, P<0.0001; PtdEth, r2=0.76, P<0.0001.
Unsaturation index, UI
Unsaturation index, UI
% Unsaturation
n=4 n=5
n=6
PtdEth
B
85 75
65 55
45 45 200 250 300 350 400
Tr
n=1 n=2 n=3 n=4
PtdCho
A
70 60
50 40
30 0 100 200 300
Tr
[image:6.609.52.279.72.264.2]compositional and structural characteristics are truly conserved within members from each thermal environment and provides greater confidence in the broad applicability of the inferences drawn.
Membrane ultrastructure is complex, comprising dynamic and static components as well as the potential for microdomains co-existing in either or both of the asymmetrically disposed monolayers (Gennis, 1989; Stubbs
and Smith, 1984). We recognise that the biophysical technique employed in our work to characterise membrane structure does not encompass all these diverse elements, but emphasises the degree of static ordering. However, we have recently used a catalytic hydrogenation technique to demonstrate that DPH anisotropy measured under steady-state illumination is very sensitive to variations in membrane saturation in native membranes from trout (Logue et al., 1998). Using DPH, we
% MUFA and PUFA
% MUFA and PUFA
% Unsaturation
B
PtdEth 85 75
65 55
45 45 0 20 40 60
Tr
A
PtdCho 70 60
50 40
30 5 15 25 35 45
[image:7.609.57.295.182.626.2]Tr
Fig. 5. The contribution of mono-unsaturated (MUFA, open symbols) and polyunsaturated fatty acids (PUFA, filled symbols) to the interspecific differences in percentage unsaturated fatty acids in phosphatidylcholine (PtdCho) (A) and phosphatidylethanolamine (PtdEth) (B). Species are divided into four groups; Antarctic (blue), temperate (green) and tropical (orange) fish species and homeotherms (red symbols). Each data point represents a separate species (mean ± S.D., N=3 separate preparations). The trend lines
(solid for PUFA, dashed for MUFA) were drawn by inspection. The values for trout are indicated by ‘Tr’.
MUFAs PUFAs SFAs
0 20 40 60 80 100
Rat Pagothenia Rat Pagothenia PtdEth
B
0 20 40 60 80 100
Rat Pagothenia Rat Pagothenia PtdCho
A
% Unsaturation
sn-1 sn-2
% Unsaturation
[image:7.609.329.545.183.660.2]sn-1 sn-2
Fig. 6. Distribution of saturated (open columns), mono-unsaturated (hatched columns) and polyunsaturated fatty acids (filled columns) in the sn-1 and sn-2 positions of phosphatidylcholine (PtdCho) (A) and phosphatidylethanolamine (PtdEth) (B) for brain synaptic membranes isolated from rat and the Antarctic fish Pagothenia
show here that all the five Antarctic species have a very similar biophysical order, which is quite different from that shared by the eight semi-tropical species. We have previously shown that the membranes isolated from the brains of four small mammal species were also highly conserved (Behan-Martin et al., 1993). All species, including the birds and mammals, show DPH anisotropies that relate very closely to their respective adaptation temperatures and agree closely with results obtained from single representative species (Behan-Martin et al., 1993; Cossins and Prosser, 1977). As before (Behan-Martin et al.,
1993), these interspecific differences were sufficiently large to provide an almost complete conservation of biophysical order, as indicated by DPH anisotropy, determined at each of the respective adaptation temperatures. This applies particularly to all the non-Antarctic species; for example, trout and rat were almost identical. Although the Antarctic species possessed the most disordered membranes, the membranes of none of them were sufficiently disordered to present identical DPH anisotropies at Antarctic temperatures to those of the other species at their respective adaptation temperatures. This was noticeable in previous studies comparing single representative species where, however, its significance was not recognised (Behan-Martin et al., 1993; Cossins and Prosser, 1977). This may reflect a limitation to the degree of cold compensation that can be achieved.
The lipid composition data show that the important features of membrane fatty acid composition were also highly conserved within each species group. Thus, all the semi-tropical marine species possessed between 44 and 46 % unsaturated fatty acids in PtdCho, a value similar to that of the two tropical freshwater species. The corresponding value for the Antarctic group was 55–61 %, whilst for the bird and mammal it was less than 40 %. Similar conservation was evident in the PtdEth lipid class. The proportion of unsaturated fatty acids in both headgroup classes was closely and linearly related to adaptation temperature, although the dependence was substantially greater for PtdEth than for PtdCho. Hazel and Landrey (1988), in reviewing a variety of studies of thermally acclimated animals, found that a 20 °C decrease in temperature was associated with a 19 % increase in unsaturated fatty acids. The comparable values established from the interspecific comparison shown here were 11 % in PtdCho and 16 % in PtdEth.
The unsaturation index for PtdCho was closely and linearly related to adaptation temperature, whilst for PtdEth the relationship was more complex. In the former case, this was not simply related to the increase in the proportion of unsaturated fatty acids, but to a progressive increase in the average number of unsaturation bonds per unsaturated fatty acid as adaptation temperature decreased. This was due to the preferential replacement of saturated fatty acids by long-chain (C20–C22) polyunsaturated fatty acids at lower temperatures, the proportion of mono-unsaturated fatty acids in PtdCho remaining broadly similar in all species. In contrast, for PtdEth, the unsaturation index for most teleost species, in particular for the semi-tropical and Antarctic species, did not vary. This was largely because the proportion of long-chain polyunsaturates remained similar in these groups. The proportion of mono-unsaturated fatty acids, however, showed large interspecific differences that related closely both to changes in percentage unsaturates and to adaptation temperature.
[image:8.609.51.283.215.682.2]Thus, an important outcome of the compositional analyses presented here is a recognition of the different responses to temperature of the two major phospholipid headgroup classes; varying long-chain PUFAs in PtdCho and mono-unsaturated fatty acids in PtdEth. Polyunsaturated fatty acids are generally
Fig. 7. Comparison of the major molecular species of phosphatidylcholine (PtdCho) (A) and phosphatidylethanolamine (PtdEth) (B) for brain synaptic membranes from rat (open columns) and the Antarctic fish Pagothenia borchgrevinki (filled columns).
16:0/18:1 16:0/22:6
0 20 40 60
A
PtdCho18:0/22:6 18:1/22:6
0 10 20 30
40
B
PtdEthPagothenia Rat
% of total molecular species
substituted on the sn-2 position, and the comparison of rat and
Pagothenia borchgrevinki molecular species supports the idea
that, with respect to PtdCho, it is changes specifically at this position that account for the cold-adaptive response. Conversely, the sn-1 position is largely occupied by saturates and mono-unsaturates. The molecular species comparison for PtdEth shows a cold-related substitution of mono-unsaturates for saturates at the sn-1 position, with little change at the sn-2 position. Thus, thermally related changes in lipid composition are positionally specific and differ for the two principal phosphoglycerides.
These headgroup-specific differences correspond well with observations of the membranes of thermally acclimated animals (Buda et al., 1994; Farkas and Roy, 1989; Tiku et al., 1996). Thus, in carp liver microsomes (Tiku et al., 1996), the MUFA composition of PtdEth increased from 15 to 30 % within a 10-day period after transfer from 30 to 10 °C, whilst there were little change in the proportion of long-chain PUFAs. Conversely, the proportion of PUFAs in PtdCho increased from 38 to 46 %, whilst MUFAs were unaltered. The response of PtdEth was shown to be due to replacement of a saturate with a mono-unsaturate at the sn-1 position of molecular species containing 22:6n-3 in the sn-2 position.
The significance of these headgroup-specific compositional modifications is not clear, but may be related to the maintenance of an appropriate phospholipid molecular shape over the full biological range of temperatures. Molecular shape is thought to be critically important for lipid organisation (Israelachvilli et al., 1980; Wieslander et al., 1980), with a mixture of cylindrical and conical lipid molecules producing an appropriate balance between bilayer stabilising (cylinders) and destabilising (cones) effects in biological membranes (Gruner, 1985). Normally, the destabilising lipids are present as a small mole fraction, and a decrease in temperature must be accompanied either by increasing the proportion of destabilising lipids or by accentuating the wedge-shape properties of these molecules by promoting increased fatty acid unsaturation.
Phosphatidylethanolamine has a predominantly conical molecular shape because of the small headgroup relative to the large swept volume of the disordered and mobile fatty acid chains. It is noticeable both from our data and from that of others that the PtdEth of mammalian brain synaptic membranes contains a long-chain polyunsaturated fatty acid, mainly 22:6n-3, on the sn-2 position, a feature contributing to the conical shape of the entire molecule. Therefore, increases in unsaturation on cold adaptation cannot occur in this position, but must be limited to the sn-1 position by conversion of a saturated fatty acid to a mono-unsaturated fatty acid (Buda et al., 1994; Farkas and Roy, 1989; Tiku et al., 1996). In contrast, PtdCho has a more cylindrical shape in which the combined cross-sectional area of the two fatty acids matches that of the bulkier headgroup. The PtdCho of the mammalian membranes contains a saturate on the
sn-1 position and a less than highly polyunsaturated fatty acid
on the sn-2 position. Adaptation to lower body or environmental temperatures can therefore be accommodated by increasing the
unsaturation in the sn-2 position. Why the alternative strategy of replacing the saturate in sn-1 with a mono-unsaturate is not employed is not clear, but there does appear to be a preferred tendency for increasing PUFA at sn-2. It has been suggested that increased polyunsaturation at sn-2 imparts a greater degree of compressibility to the bilayer, enabling the conformational changes required of membrane-bound enzymes (Litman and Mitchell, 1996). Either way, it is clear that the positional distribution of fatty acids is a critical and apparently adaptive component of the lipid compositional response to temperature change.
It is important in comparative studies of this type to separate truly adaptive responses from characters that are distributed among species for reasons of biogeography or phylogenetic lineage. A number of approaches have been suggested to resolve this issue, all of which require the phylogenetic relationships of the species compared to be fully understood (Harvey and Pagel, 1991; Pagel, 1999). Unfortunately, most of the species examined here were members of the order Perciformes, for which the phylogeny of the constituent genera has not been adequately resolved (Nelson, 1994). This makes a proper description of the influence of phylogeny on the characters described here impossible to achieve. However, we have shown that two families with resolved phylogenies, the Salmonidae and the Nototheniidae, and which both inhabit cold environments possessed brain membranes that were equally unsaturated and disordered. Similarly, the warm-water toadfish (Batrachoidiformes) had low levels of unsaturation and ordered brain membranes similar to those of the other tropical marine species (Perciformes). These correlations lend support to the idea that these membrane attributes have adaptive significance for cold rather than being due to phylogenetic inheritance.
Finally, it is instructive to question the physiological significance of interspecific differences in lipid composition and physical structure. The conservation of membrane physical structure, as defined by the fluorescence polarisation technique with DPH as probe, is thought to be a compensatory response maintaining this property largely independent of temperature. Any functional property of these neuronal membranes that is sensitive to lipid unsaturation and biophysical order should therefore be compensated. There is a significant literature detailing the concept of ‘viscotropic control’ and the effects of the hydrocarbon disorder upon the properties of membrane proteins (Hazel and Williams, 1990). However, the general concept of membrane ‘fluidity’ affecting membrane protein function is controversial and, at least in the case of the Ca2+
With regard to heat-induced damage and heat death at the level of the whole animal, Friedlander et al. (1976) demonstrated in a classic experiment that heat death of goldfish was linked to disturbed functioning of central neurones. Further correlative experiments have linked changes in high-temperature resistance during thermal acclimation of goldfish with changes in lipid unsaturation (Cossins, 1977; Roots and Prosser, 1962). The species used in the present study have thermal death temperatures that broadly increase with increasing habitat temperature, with the Antarctic species undergoing heat death at 6–10 °C (Somero and De Vries, 1967), the palearctic and temperate species at 20–30 °C (Brett, 1952), the semi-tropical, tropical and continental species at 30–40 °C (Friedlander et al., 1976) and mammals at 42–46 °C (Gordon, 1993). Lipid composition and the physical structure of the brain membranes are also related to adaptation temperature, and it follows that heat injury is broadly correlated with the lipid biochemistry of the brain membranes. However, the correlation is not precise for all combinations of species; thus, in common with most salmonids, rainbow trout display heat death at approximately 22–25 °C, yet the brain lipid saturation and membrane structural order are virtually identical to those of members of the Antarctic species group with much lower heat death temperatures.
We thank Professor P. J. Walsh and Mr S. Fitzherbert-Brockholes for providing fish specimens. J.A.L. was supported by a postgraduate studentship from the Biotechnology and Biological Research Council (UK).
References
Behan-Martin, M., Bowler, K., Jones, G. and Cossins, A. R.
(1993). A near perfect temperature adaptation of bilayer order in vertebrate brain membranes. Biochim. Biophys. Acta 1151, 216–222.
Bell, M. V. and Dick, J. R. (1991). Molecular species composition
of the major diacyl glycerophospholipids from muscle, liver, retina and brain of cod (Gadus morhua). Lipids 26, 565–573.
Brett, J. R. (1952). Temperature tolerances in young Pacific salmon
Oncorhynchus. J. Fish. Res. Bd Can. 9, 265–323.
Buda, C., Dey, I., Baloch, N., Horvath, L. I., Maderspach, K., Juhasz, M., Yeo, Y. K. and Farkas, T. (1994). Structural order
of membranes and composition of phospholipids in fish brain cells during thermal acclimatisation. Proc. Natl. Acad. Sci. USA 91, 8234–8238.
Cossins, A. R. (1977). Adaptation of biological membranes to
temperature: The effect of temperature acclimation of goldfish upon the viscosity of synaptosomal membranes. Biochim. Biophys. Acta
470, 395–411.
Cossins, A. R. (1994). Temperature Adaptation of Biological
Membranes. London: Portland Press.
Cossins, A. R., Christiansen, J. and Prosser, C. L. (1978).
Adaptation of biological membranes to temperature: Lack of homeoviscous adaptation in the sarcoplasmic reticulum of goldfish muscle during thermal adaptation. Biochim. Biophys. Acta 511, 442–454.
Cossins, A. R. and Macdonald, A. G. (1986). Homeoviscous
adaptation under pressure. III. The fatty acid composition of liver mitochondrial phospholipids of deep sea fish. Biochim. Biophys.
Acta 860, 325–335.
Cossins, A. R. and Prosser, C. L. (1977). Evolutionary adaptations
of membranes to temperature. Proc. Natl. Acad. Sci. USA 75, 2040–2043.
Cossins, A. R. and Prosser, C. L. (1982). Variable homeoviscous
responses of diferent brain membranes of thermally-acclimated goldfish. Biochim. Biophys. Acta 687, 303–309.
Cossins, A. R. and Raynard, R. S. (1987). Adaptive responses of
cell membranes to temperature. In Temperature and Animal Cells, vol. 41 (ed. K. Bowler and B. J. Fuller), pp. 95–111. Cambridge: Company of Biologists Ltd.
Crockett, E. L. and Hazel, J. R. (1995). Cholesterol levels explain
inverse compensation of membrane order in brush border but not homeoviscous adaptation in basolateral membranes from the intestinal epithelia of rainbow trout. J. Exp. Biol. 198, 1105–1113.
Cunningham, J. R. C. and Hyde, D. (1995). The short and long term
effects of temperature on the dynamic range of signalling in horizontal cella of carp retina. J. Therm. Biol. 20, 223–229.
Farkas, T. and Roy, R. (1989). Temperature-mediated restructuring
of phosphatidylethanolamines in livers of freshwater fishes. Comp.
Biochem. Physiol. 93B, 217–222.
Friedlander, M. J., Kotchabhakdi, N. and Prosser, C. L. (1976).
Effects of cold and heat on behaviour and cerebellar function in goldfish. J. Comp. Physiol. 112, 19–45.
Gennis, R. B. (1989). Biomembranes. Molecular Structure and
Function. New York: Springer Verlag.
Gordon, C. J. (1993). Temperature Regulation in Laboratory
Rodents. Cambridge: Cambridge University Press.
Gruner, S. M. (1985). Intrinsic curvature hypothesis for biomembrane lipid composition: a role for non-bilayer lipids. Proc.
Natl. Acad. Sci. USA 82, 3665–3669.
Harvey, P. H. and Pagel, M. D. (1991). The Comparative Method
in Evolutionary Biology. New York: Oxford University Press.
Hazel, J. R. (1995). Thermal adaptation in biological membranes –
is homeoviscous adaptation the explanation? Annu. Rev. Physiol.
57, 19–42.
Hazel, J. R. and Landrey, S. R. (1988). Time-course of thermal
adaptation in plasma membranes of trout kidney. I. Headgroup composition. Am. J. Physiol. 255, R622–R627.
Hazel, J. R. and Williams, E. E. (1990). The role of alterations in
membrane lipid composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 29, 167–227.
Israelachvilli, J. N., Marcelja, S. and Horn, R. G. (1980). Physical
principles of membrane organisation. Q. Rev. Biophys. 13, 121–200.
Lee, A. G. (1991). Lipids and their effects on membrane proteins:
Evidence against a role for fluidity. Prog. Lipid Res. 30, 323–348.
Lee, J. A. C. and Cossins, A. R. (1990). Temperature adaptation of
biological membranes: differential homoeoviscous responses in brush-border and basolateral membranes of carp intestinal mucosa.
Biochim. Biophys. Acta 1026, 195–203.
Litman, B. and Mitchell, D. (1996). A role for phospholipid
polyunsaturation in modulating membrane protein function. Lipids
31, S193–S197.
Logue, J. A., Vigh, L., Joo, F. and Cossins, A. R. (1998). Catalytic
hydrogenation of polyunsaturated biological membranes: effects on membrane fatty acid composition and physical properties. Biochim.
Nelson, G. (1994). Fishes of the World. New York: J. Wiley & Sons,
Inc.
Pagel, M. (1999). Inferring the historical patterns of biological
evolution. Nature 401, 877–884.
Roots, B. I. and Prosser, C. L. (1962). Temperature acclimation and
the nervous system in fish. J. Exp. Biol. 39, 617–629.
Somero, G. N. and De Vries, A. L. (1967). Temperature tolerance
of some Antarctic fishes. Science 156, 257–258.
Stubbs, C. and Smith, A. D. (1984). The modification of mammalian
membrane polyunsaturated fatty acid composition in relation to membrane fluidity and function. Biochim. Biophys. Acta 779, 89–137.
Takamura, H. and Kito, M. (1991). A highly sensitive method
for quantitative analysis of phospholipid molecular species by high-performance liquid chromatography. J. Biochem. 109, 436–439.
Tiku, P. E., Gracey, A. Y., Macartney, A. I., Beynon, R. J. and Cossins, A. R. (1996). Cold-induced expression of ∆9-desaturase in carp by transcriptional and posttranslational mechanisms.
Science 271, 815–818.
Wieslander, A., Christiansson, A., Rilfors, L. and Lindblom, G.