The Journal of Experimental Biology
ABSTRACT
Larval and adult fruit flies are attracted to volatiles emanating from food substrates that have been occupied by larvae. We tested whether such volatiles are emitted by the larval gut bacteria by conducting tests under bacteria-free (axenic) conditions. We also tested attraction to two bacteria species, Lactobacillus brevis, which we cultured from larvae in our lab, and L. plantarum, a common constituent of fruit flies’ microbiome in other laboratory populations and in wild fruit flies. Neither larvae nor adults showed attraction to axenic food that had been occupied by axenic larvae, but both showed the previously reported attraction to standard food that had been occupied by larvae with an intact microbiome. Larvae also showed significant attraction to volatiles from axenic food and larvae to which we added only either L. brevisor L. plantarum, and volatiles from L. brevis reared on its optimal growth medium. Controlled learning experiments indicated that larvae experienced with both standard and axenic used food do not perceive either as superior, while focal larvae experienced with simulated used food, which contains burrows, perceive it as superior to unused food. Our results suggest that flies rely on microbiome-derived volatiles for long-distance attraction to suitable food patches. Under natural settings, fruits often contain harmful fungi and bacteria, and both L. brevisand L. plantarumproduce compounds that suppress the growth of some antagonistic fungi and bacteria. The larval microbiome volatiles may therefore lead prospective fruit flies towards substrates with a hospitable microbial environment.
KEY WORDS: Drosophila melanogaster, Fruit flies, Larvae, Microbiome, Social behaviour
INTRODUCTION
Social behaviour varies widely among animals. While most species show minimal social interaction, typically only in the context of sexual behaviour and aggression, other taxa are highly social within all facets of life, and some species are even obligatorily social. Although much of the research on social behaviour has focused on elaborate cases of sociality (Wilson, 1971; Michener, 1974; Wilson, 1975; Costa, 2006), there has been increased interest in assessing social behaviour in simple animal models that are highly amenable to neurogenetic research (Robinson et al., 2008; Sokolowski, 2010). Fruit flies (Drosophila melanogaster) are an ideal species for such investigation and their larvae are especially attractive owing to their exceptionally small number of a few thousand functional neurons (Nassif et al., 2003; Iyengar et al., 2006; Pauls et al., 2010; Huser et al., 2012).
RESEARCH ARTICLE
1Animal Behaviour Group, Department of Psychology, Neuroscience and Behaviour, McMaster University, 1280 Main Street West, Hamilton, ON L8S 4K1, Canada. 2Department of Biology, McMaster University, 1280 Main Street West, Hamilton, ON L8S 4K1, Canada.
*Author for correspondence ([email protected])
Received 5 November 2013; Accepted 26 December 2013
Fruit fly aggregations at fallen fruit in orchards are quite familiar to field biologists. Indeed, in addition to their natural attraction to odours from ripe and fermenting fruit (Barrows, 1907; Zhu et al., 2003; Stökl et al., 2010), fruit flies produce and show long-distance attraction to cisvaccenyl acetate (cVA), which effectively serves as an aggregation pheromone (Brieger and Butterworth, 1970; Bartelt et al., 1985; Wertheim et al., 2005). Such attraction can cause many females to lay eggs at the same site and, consequently, aggregation of eggs and larvae.
We have recently found that larval and adult fruit flies show significant attraction to odours emanating from food substrates that have been occupied by larvae. Furthermore, larvae and adults learn to prefer odours from food substrates that have been occupied by larvae over odours from unoccupied substrates of similar quality (Durisko and Dukas, 2013; Durisko et al., 2014). Our controlled experiments indicated that the social attraction we documented was not caused by odours from either fruit or yeast. We also found no larval or adult attraction to ammonia, the predominant nitrogen waste compound in fruit flies (Borash et al., 1998), which has a salient odour (Z.D. and R.D., unpublished data). We thus hypothesized that bacterial odours mediate social attraction in fruit flies because bacteria often produce salient volatiles used as cues by a wide variety of animals (Archie and Theis, 2011; Davis et al., 2013). To evaluate this hypothesis, we first tested whether larvae and adults are attracted to bacterial odours emanating from food occupied by larvae. After identifying gut bacteria as the source of the attractive odours, we examined whether the bacteria produce such odours on their own or only when residing in larval guts. Finally, we wished to examine why flies are attracted to the bacterial odours. To this end, we first assessed the relative values that larvae assign to food previously occupied by larvae with bacteria and to axenic food previously occupied by axenic larvae. Having shown that larvae do not prefer cues previously associated with bacteria, we tested whether the burrows generated by larvae digging into the food, which provide hiding sites, can explain larval preference for used over fresh food.
RESULTS
Origin of the attractive volatiles: larval attraction
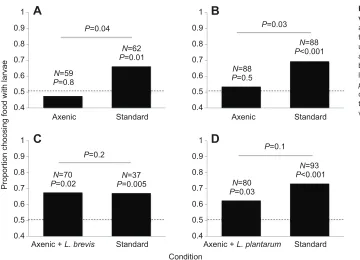
Focal larvae showed no stronger attraction to axenic used food with axenic larvae than to axenic fresh food with no larvae [47.5%, N=59, generalized linear model (GLM): Wald’s χ21=0.138, P=0.8; Fig. 1A, left bar]. However, focal larvae did show stronger attraction to standard used food with standard larvae than to standard fresh food with no larvae (66.1%, N=62, GLM: Wald’s χ21=6.2, P=0.01; Fig. 1A, right bar; Wald’s χ21=4.1, P=0.04 for attraction to standard used food with standard larvae versus attraction to axenic used food with axenic larvae; Fig. 1A, left versus right bars).
Origin of the attractive volatiles: adult attraction
Adult females entered at similar frequencies into vials containing axenic used food with axenic larvae and vials containing axenic fresh food without larvae (53.4% versus 46.6%, respectively, N=88,
Social attraction mediated by fruit flies’ microbiome
The Journal of Experimental Biology
GLM: Wald’s χ21=0.455, P=0.5; Fig. 1B, left bar). Under thestandard conditions, females entered at higher frequencies into vials containing standard used food with standard larvae than into vials containing standard fresh food without larvae (69.3%, N=88, GLM: Wald’s χ21=12.5, P<0.001; Fig. 1B, right bar; Wald’s χ21=4.5, P=0.03 for the standard versus axenic conditions; Fig. 1B, left versus right bars).
Identification of and attraction test with Lactobacillus brevis
The bacterium we identified belonged to Lactobacillus brevis. Focal larvae showed stronger attraction to the axenic used food with axenic larvae and L. brevisthan to axenic fresh food with no larvae (67.1%, N=70, GLM: Wald’s χ21=5.3, P=0.02; Fig. 1C, left bar). As before, focal larvae showed stronger attraction to the standard used food with standard larvae than to standard fresh food with no larvae (67.6%, N=37, GLM: Wald’s χ21=7.7, P=0.005; Fig. 1C, right bar). Larval attraction to the used food was similar in the standard and axenic with L. brevistreatments (GLM: Wald’s χ21=1.5, P=0.2; Fig. 1C, left versus right bars).
Attraction test with L. plantarum
Larvae showed stronger attraction to the axenic used food with axenic larvae and L. plantarumthan to axenic fresh food by itself (62.5%, N=80, GLM: Wald’s χ21=4.6, P=0.03; Fig. 1D, left bar). Larval attraction to the used food was similar when it was axenic with L. plantarumand standard (GLM: Wald’s χ21=2.4, P=0.1; Fig. 1D, left versus right bars).
Attractiveness of free-living L. brevis
Larvae showed a significant attraction to L. brevison MRS agar (66.0%, N=50, GzLM: Wald’s χ21=4.9, P=0.03) but not to L. brevis on fly medium (45.2%, N=62, GLM: Wald’s χ21=0.568, P=0.451; Fig. 2). As before, the larvae showed significant attraction to L. breviswith larvae on fly medium (63.6%, N=66, GLM: Wald’s χ21=4.9, P=0.03; Fig. 2). Overall, there was a significant effect of treatment (GLM: Wald’s χ21=6.5, P=0.04), with larval attraction to
L. brevison MRS agar being similar to larval attraction to L. brevis with larvae on fly medium (GLM: Wald’s χ21=0.07, P=0.8), and larval attraction to L. brevison fly medium being significantly lower than larval attraction to L. breviswith larvae on fly medium (GLM: Wald’s χ21=4.6, P=0.03).
Do larvae perceive standard food as superior to axenic food?
Focal larvae showed no preference for odours previously paired with standard used food over odours previously paired with axenic used food (54.3%, N=35, GLM: Wald’s χ21=0.221, P=0.6; Fig. 3A). In the control condition, we did find the expected significant larval preference for novel odours previously paired with standard used food over novel odours previously paired with standard fresh food (70.3%, N=37, GLM: Wald’s χ21=5.2, P=0.02; GLM: Wald’s χ21=3.9, P=0.047 for the between-treatments comparison; Fig. 3A).
Do larvae prefer used food due to ease of burrowing? Focal larvae showed a preference for odours previously paired with artificially used food over odours previously paired with fresh food similar to their preference for odours previously paired with standard used food over odours previously paired with fresh food (70.8% and 73.9%, respectively; N=47; GLM: Wald’s χ21=0.03, P=0.856; Fig. 3B). Analyzed separately, in both the treatment and control conditions, we observed a significant preference for the odour previously paired with artificially used and standard used food (respectively, N=24, GLM: Wald’s χ21=4.2, P=0.041 and N=23, GLM: Wald’s χ21=4.0, P=0.046).
DISCUSSION
Attraction to microbiome volatiles
By generating axenic larval cultures and conducting experiments with two bacterial species, we critically established that microbiome volatiles serve as attractants for the fruit fly D. melanogaster. Both larval and adult stages were attracted to these volatiles (Fig. 1). While we used our long-term laboratory population in the present study, we have previously shown attraction to food occupied by
A
B
D
C
N=62
P=0.01
N=59
P=0.8
0.4 0.5 0.6 0.7 0.8 0.9 1
Axenic Standard
Proportion choosing food with larvae
P=0.04
N=88
P<0.001
N=88
P=0.5
0.4 0.5 0.6 0.7 0.8 0.9 1
Axenic Standard
P=0.03
N=37
P=0.005
N=70
P=0.02
0.4 0.5 0.6 0.7 0.8 0.9 1
Axenic + L. brevis Standard
Condition
P=0.2
Axenic + L. brevis
N=93
P<0.001
N=80
P=0.03
0.4 0.5 0.6 0.7 0.8 0.9 1
Standard
P=0.1
Axenic + L. plantarum
[image:2.612.47.407.56.316.2]The Journal of Experimental Biology
larvae in tests with wild-caught larval and adult fruit flies as well asin experiments using natural fruit (Durisko and Dukas, 2013; Durisko et al., 2014). Volatiles from both L. brevis, which we cultured from our fly population, and L. plantarum, which has been isolated from the Drosophilagut in other studies, were attractive. These two bacterial species are common constituents of the fruit-fly microbiome in both laboratory and field populations (Chandler et al., 2011; Erkosar et al., 2013). It would be interesting to see whether other major species of the fruit fly microbiome also produce volatiles attractive to fruit flies. Although L. brevisresides in the fly gut, it produces the attractive volatiles even when isolated from larvae and reared on its optimal growth medium (Fig. 2). Nevertheless, cultures of L. brevison fly medium did not grow well and were not attractive to larvae (Fig. 2), suggesting that the bacterial volatiles could actually serve as a long-distance cue indicating the presence of larvae feeding on adequate substrate.
Why are larvae and adults attracted to microbiome volatiles?
It is well established that fruit flies are highly attracted to volatiles associated with fermenting fruit (Barrows, 1907; Sturtevant, 1921; Hutner et al., 1937; Spencer, 1950; Ruebenbauer et al., 2008; Becher et al., 2010), and recent data indeed indicate strong olfactory receptor activity in response to volatiles associated with yeast volatiles (Stökl et al., 2010; Becher et al., 2012). Such findings are expected given that D. melanogasterfeed on yeast that grows on fruit (Begon, 1982). Fruit flies are also attracted to ripe-fruit volatiles (Zhu et al., 2003), which is also sensible given that such substrates may already contain yeast and that fruit flies can inoculate fruit with yeast (Gilbert, 1980; Wertheim et al., 2002; Stamps et al., 2012). It has also been known for several decades that adult fruit flies show long-distance attraction to cVA, which is produced by males and transferred to females during copulation (Brieger and Butterworth, 1970; Bartelt et al., 1985). cVa seems to serve a social function because it informs males about the presence of females and informs females about the presence of mated females at adequate egg-laying substrates. Given these direct cues indicating food and conspecifics, it is not immediately clear why larvae and adults also show attraction to microbiome-derived volatiles.
Focal larvae that experienced both standard and axenic food that had been previously occupied by larvae did not prefer the novel odour associated with the standard used food, which contained bacterial volatiles (Fig. 3A). In contrast, focal larvae perceived artificially used food as better than unused food (Fig. 3B). Our data
thus support the notion that larvae prefer used food because it is easier to burrow into than fresh food. Burrowing into the food probably allows larvae to reduce attack rates by parasitoid wasps (Carton and David, 1985; Rohlfs and Hoffmeister, 2004), which are a major cause of larval mortality (Carton et al., 1986; Fleury et al., 2004). Thus, our current evidence indicates that microbiome volatiles serve as public information (Danchin et al., 2004) cues directing larvae towards potentially superior feeding sites.
Bacteria as prey?
Perhaps the simplest explanation for fruit flies’ attraction to bacterial volatiles is that fruit flies feed on bacteria. Indeed, bacteria-feeding insects such as tephritid flies (Tephritidae) are attracted to odours emanating from their bacterial prey (Robacker et al., 2004; Robacker et al., 2009). It is well established, however, that fruit flies are strongly attracted to and feed on yeast (Spencer, 1950; Begon, 1982). Furthermore, the fruit fly microbiome, which includes the two Lactobacillus species we have examined, is well studied (Sharon et al., 2010; Wong et al., 2011; Broderick and Lemaitre, 2012; Ridley et al., 2012), and critical experiments have shown that L. plantarumis a fruit fly gut commensal rather than prey (Storelli et al., 2011).
N=50
P=0.03
N=62
P=0.4
N=66
P=0.03
0.4 0.5 0.6 0.7 0.8 0.9 1
Proportion choosing food
with
L. brevis
Treatment
P=0.03
P=0.8
L. brevis on MRS L. brevis on fly
medium L. brevis
+ larvae + fly medium
Fig. 2. Attraction of focal larvae in three binary choices. Choices involved Lactobacillus brevison MRS medium versus MRS medium, L. brevison fly medium versus fly medium, and L. breviswith larvae on fly medium versus fly medium.
A
B
N=37
P=0.02
N=35
P=0.6
0.4 0.5 0.6 0.7 0.8 0.9 1
Standard used versus axenic used
Standard used versus standard fresh
Proportion choosing odour paired with standard used food
P=0.047
N=23
P=0.046 N=24
P=0.041
0.4 0.5 0.6 0.7 0.8 0.9 1
Artificially used versus standard fresh
Standard used versus standard fresh Condition
P=0.86
The Journal of Experimental Biology
Microbiome volatiles as salient cues
It is possible that the fruit flies’ microbiome produces the most salient cues available for larvae and adults seeking either others or suitable food. While it is not clear to us why this would be the case, the fact is that bacteria release a remarkable number of volatiles (Schulz and Dickschat, 2007) salient to many animals, including humans (Lam et al., 2007; Archie and Theis, 2011; Leroy et al., 2011; Davis et al., 2013). The level of saliency of a volatile, however, is not merely a passive chemical property, because it can also reflect an evolved adaptation by an animal. Indeed, fruit flies possess a dedicated olfactory circuit for detecting geosmin, a volatile aversive to adults, which is emitted by species of bacteria and fungi that are harmful to fruit flies (Becher et al., 2010; Stensmyr et al., 2012). We thus propose that fruit flies possess specialized abilities to detect microbiome volatiles because such odour cues are inadvertent by-products of the successful foraging of other individuals, providing larvae and adults with the best available information about the suitability of a food patch. While adult and larval fruit flies can taste a substrate in order to obtain some information about food quality, taste alone cannot indicate the presence of all essential nutrients. Furthermore, fruit flies are under intense competition with numerous species of fungi and bacteria for feeding on fallen fruit, and many microbes produce secondary compounds that harm other microbes as well as many other animals, including fruit flies (Janzen, 1977; Demain and Fang, 2000; Rohlfs et al., 2005; Rozen et al., 2008). It is thus possible that microbiome volatiles signal to flies the availability of a food substrate with suitable microbial species.
Benefits of the microbiome
While the importance of the complex interactions between animals and their microbiome has been appreciated for a long time (e.g. Drasar and Hill, 1974; Savage, 1977), such interactions have recently been subjected to intensive research fuelled by powerful and relatively cheap genetic tools (e.g. Ben-Yosef et al., 2010; Ezenwa et al., 2012; Chu et al., 2013; Faith et al., 2013; Foster and McVey Neufeld, 2013). In agreement with findings in other species, research on fruit flies indicates that their gut bacteria accelerate the larval developmental rate under nutritional deficiency (Storelli et al., 2011; Ridley et al., 2012). Another possible benefit of gut bacteria is suppression of competing or harmful microbial species. Mould can cause high rates of mortality among fruit fly larvae, and groups of larvae can suppress mould growth as well as enhance the growth of certain yeast species (Rohlfs et al., 2005; Stamps et al., 2012). Intriguingly, both species of Lactobacilluswe have studied produce compounds that suppress fungal and bacterial growth (Ruiz-Barba et al., 1994; Laitila et al., 2002; Schnürer and Magnusson, 2005; Mauch et al., 2010; Crowley et al., 2012). The fruit fly microbiome may thus help larvae control the species composition of fungi and bacteria on their fruit substrates. Such symbiotic interactions between insects and bacteria for controlling harmful microbes are known in the well-studied fungus-growing ants and beetles (Currie et al., 1999; Scott et al., 2008), but may be prevalent in other species as well (e.g. Fredenhagen et al., 1987).
Prospects
There has recently been a resurgence in research on animal–microbiome interactions in which fruit flies are emerging as an important model system (Erkosar et al., 2013). Our work links such research, which has focused on mechanisms of such interactions, with animal behaviour and ecology. We have established that fruit flies are attracted to volatiles originating from
their microbiome and that such attraction can lead them to favourable feeding sites. We propose that the microbiome volatiles guide fruit flies to patches with a hospitable microbial environment, which has been generated at least in part via suppression of hostile microbes by the larval microbiome. Fruit flies, given the availability of powerful genetic tools, and Lactobacillusspp., owing to ample research by the food industry (Gobbetti, 1998; Schnürer and Magnusson, 2005), may be an ideal model system for testing hypotheses linking host–microbiome interactions with animal social behaviour and ecology.
MATERIALS AND METHODS
We maintained two population cages of several hundred Drosophila
melanogasterCanton-S following standard protocol (Sarin and Dukas, 2009). We generated axenic cultures under a laminar flow cabinet by
sterilizing 12 h embryos with 2 min of immersion in 2.5% sodium
hypochlorite, followed by two washes each with 70% ethanol and sterile distilled water (Brummel et al., 2004). We transferred sterilized embryos to autoclaved axenic food dishes, which consisted of autoclaved standard fly
food supplemented with two types of antibiotics (50 mg l−1ampicillin and
20 mg l−1 chloramphenicol) and two antifungal agents (2 g l−1 methyl
paraben and 10 mg l−1fluconazole). We verified that the axenic cultures
were microbial-free by plating homogenates on Luria-Bertani agar plates.
For standard flies, we washed 12 h embryos four times with sterile distilled
water before transferring to standard food dishes supplemented only with methyl paraben. We transferred Petri dishes containing standard or axenic embryos to a plastic chamber maintained at 25°C, 90% relative humidity and kept in total darkness. For all experiments, an observer blind to the experimental treatments recorded the data. Our main statistical analyses involved generalized linear models (GLMs) with a binomial distribution and logit link function.
Origin of the attractive volatiles Larval attraction
We first tested whether larvae and adults show the same strong attraction to axenic used food with axenic larvae as they do to standard used food with standard larvae. Having found no attraction to axenic used food with axenic
larvae, we cultured and identified Lactobacillus brevisfrom our fly larvae.
We then tested whether focal larvae are attracted to axenic larvae
supplemented with L. brevis. To broaden our investigation, we also tested
whether focal larvae are attracted to axenic larvae supplemented with L.
plantarum, which has been identified as an important component of fruit flies’ microbiome in other laboratories (Sharon et al., 2010; Storelli et al.,
2011; Wong et al., 2011), as well as in all wild populations of D.
melanogastersampled by Chandler et al. (Chandler et al., 2011).
We tested axenic mid third-instar focal larvae (n=128) individually under
one of two conditions. The standard condition was identical to that of Durisko and Dukas (Durisko and Dukas, 2013), and consisted of testing each focal for attraction to standard used food that has been occupied and
consumed by 30 standard larvae for 24 h versus standard fresh food aged for
24 h. The axenic condition involved testing each focal for attraction to
axenic used food that has been occupied and consumed by 30 axenic larvae
for 24 h versus axenic fresh food aged for 24 h. We placed the 2.5 ml food
discs 1 cm apart, equidistant to the midline, alternating sides between trials.
We tested focals individually, placing one at a time facing away from the
experimenter, parallel to the midline of a fresh 100 mm agar Petri dish,
through a 1 cm opening in the lid. We recorded the choice of focal larvae,
as indicated by the first physical contact with a food disc. We terminated trials after larval first contact with a disc. We discarded the few focals that
did not make a choice within 5 min.
Adult attraction
We used 3-day-old mated adult females (n=180) placed individually in cages
(20×12×13 cm) each containing two regular 40 ml vials located in the far
corners of each cage and each containing 5 ml fly medium. Funnels at the top
The Journal of Experimental Biology
attraction experiment: standard used food with standard larvae versus freshfood, and axenic used food with axenic larvae versus axenic fresh food. We
recorded the presence of females inside vials after 16 h as an indication of
choice, discarding females that did not enter either vial.
Identification of and attraction test with L. brevis
To isolate the internal bacteria from larvae, we transferred 25 third-instar
larvae into a 12 ml two-staged centrifuge tube. We washed these larvae twice
with 70% ethanol and twice with sterile distilled water. Using a sterilized metal rod, we crushed the larvae and streaked the liquid remains on a
LactobacilliMRS plate (Man et al., 1960), which we incubated in a high
humidity chamber at 25°C. After 2 days of incubation, we found only one
type of bacterial colony morphology and we streaked cells from a single
colony onto a fresh plate of LactobacilliMRS to obtain a pure culture. Using
a sterilized loop, we transferred cells from a single colony of the second
plate to an Erlenmeyer flask containing LactobacilliMRS broth incubated
at 37°C on a rotary shaker. Prior to testing the attraction abilities of bacterial
cells, we centrifuged the bacteria in two-stage capped test tubes at 2000 g
for 4 min and washed the cells with sterile double-distilled water and then
centrifuged again to prevent carryover of medium components.
We conducted DNA extractions with an alkaline lysis miniprep procedure
(Miller, 1992). We added cells to a 1.5 ml microfuge tube containing 500 µl
of nuclease-free water. We centrifuged the tube at 10,000 gfor 5 min. We
then discarded the supernatant and re-suspended the pellet in 467 μl of TE
buffer. We added 30 μl of 20% sodium dodecyl sulfate and 3 μl of proteinase
K before incubating for 1 h at 37°C. We added phenol-chloroform and
followed up with vortexing and centrifugation to separate the DNA from proteins and other cell debris and repeated this step another time. We
precipitated the DNA by adding 100% ethanol and 50 μl sodium acetate.
Following subsequent vortexing, centrifugation and washing with 70% ethanol, we air-dried the pellet by placing the tube in an oven at 37°C for
30 min. The following two primers were used to amplify a portion of the
16S rRNA gene that spanned nucleotide positions 27–801: forward primer
27F: 5′-AGAGTTTGATCMTGGCTCAG-3′ and reverse primer 801R: 5′
-GGCGTGGACTTCCAGGGTATCT-3′. The obtained 16S rRNA sequence
was identical to that of strain ATCC 367. We used gel electrophoresis and UV light exposure for PCR product visualization. The PCR product was sequenced by fluorescence-based DNA sequencing (Mobix Laboratory, McMaster University, Hamilton, ON, Canada). The sequence was then used to search for homologues through the BLAST program at the National Center for Biotechnology Information.
To test whether the gut bacteria we isolated (L. brevis) were the source of
attractive volatiles, we tested axenic third-instar focal larvae for their
attraction to axenic food containing axenic larvae supplemented with L.
breviswe had isolated from larvae versus fresh axenic food. As a control, we repeated the standard used food with standard larvae versus fresh standard food condition. For the axenic food + gut bacteria condition, we
placed 30 axenic larvae and 50 μl of gut bacteria culture 24 h prior to testing
on axenic food disks, which consisted of autoclaved standard food supplemented with two antifungals (methyl paraben and fluconazole at the standard concentrations detailed above) and no antibacterials. We tested 120 focals as described above.
Attraction test with L. plantarum
This experiment was identical to the previous experiment except that we
used L. plantarumobtained from the American Type Culture Collection
(ATCC strain 14917). This L. plantarumstrain, which was isolated from
pickled cabbage, has been sequenced as a reference genome and thus represents traits of a general strain not specifically associated with fruit flies.
We cultured the bacteria in Erlenmeyer flasks containing LactobacilliMRS
broth incubated at 37°C on a rotary shaker. We tested the attraction of focal
larvae (n=200) to either (1) axenic used food with axenic larvae
supplemented with L. plantarumversus axenic fresh food or (2) standard
used food with standard larvae versus standard fresh food.
Attractiveness of free-living L. brevis
The purpose of this experiment was to test whether L. brevisare attractive
to larvae on their own or only when they are a part of the larval microbiome.
That is, we wished to test whether L. brevisrequires some by-products
available in the larval gut to produce the attractive volatiles. This would make the attractive volatiles specific cues indicating larval presence.
The general methods were similar to those above except that we tested the
attraction of axenic third-instar focal larvae (n=200) under three conditions:
(1) L. brevison LactobacilliMRS versus LactobacilliMRS alone, (2) L. brevison axenic fly medium versus axenic fly medium alone, and, as a
control, (3) L. breviswith axenic larvae on axenic food versus axenic food.
To ensure adequate contact with the food in the conditions with no larvae, we used a sterilized needle to create grooves in the surface and bottom of each food disk. To assess how well bacteria could grow on the fly medium, we used plate counting to compare the number of viable bacterial cells on
fly medium and on LactobacilliMRS agar [see p. 401 in Boyd (Boyd,
1988)]. The number of colony-forming units (cfu) was significantly lower on the fly medium than on MRS agar (mean ± s.e.m.=2275±69.36 and
5065±43.98 cfu 50 μl−1, respectively, t
5=34, P<0.001).
Do larvae perceive standard food as superior to axenic food? We have established that larval and adult attraction to sites with larvae is mediated by volatiles emanating from gut bacteria. As a first attempt at revealing what fruit flies gain from such attraction, we wished to examine whether focal larvae perceive food that has contained standard larvae harbouring intact get bacteria as superior to food that has contained larvae lacking bacteria. We thus allowed focal larvae to experience the two food conditions each associated with a distinct novel odour in a controlled associative learning experiment and then let them choose between the odours. We predicted that larvae would prefer the odour paired with standard food, which contains bacterial volatiles, over the odour paired with axenic food, which lacks bacterial volatiles.
We followed a standard associative learning protocol (Durisko and Dukas,
2013) and tested axenic mid-third-instar larvae (n=96). The main treatment
involved standard used food occupied and consumed by 30 standard larvae
for 24 h and axenic used food occupied and consumed by 30 axenic larvae for
24 h. The control treatment was identical to the one used by Durisko and
Dukas (Durisko and Dukas, 2013) and included standard used food occupied
and consumed by 30 standard larvae for 24 h and standard fresh food. For both
treatments, we removed larvae from the used food prior to testing. The novel odours were equally preferred by inexperienced larvae (Durisko and Dukas,
2013) and consisted of 10 μl 1-butanol and 10 μl propyl acetate diluted in
paraffin oil (1:300). Each focal larva received three 3-min training sessions on each of the two food + odour pairings. Between each training session, we rinsed focal larvae with a fresh droplet of water. Immediately following the
six training sessions, we placed each focal larva at the center of a 100 mm
Petri dish and let it choose between the two odours. Choice was defined as the first contact with a food disk under the odour cup. We randomized odour–treatment pairings during training, and odour sides in the tests.
Do larvae prefer used food due to ease of burrowing?
Our finding that larvae do not perceive food with bacterial volatiles as superior to axenic food led us to search for another factor that may lead to larval preference for used over fresh food (Durisko and Dukas, 2013). Our observations suggested that larvae prefer media that they can burrow into over media that are difficult to penetrate. Food used by other larvae is typically easier to burrow into because the other larvae have already broken the food surface. We thus conducted another experiment to assess whether larvae perceive artificially used food, in which we simulated larval burrowing into the food, as superior to fresh food. Given the results above, we predicted that focal larvae would prefer the odour paired with artificially used food over the odour paired with fresh food.
The Journal of Experimental Biology
Acknowledgements
We thank C. Baxter, A. Vogan and S. Golden for assistance, and A. Campos, K. Theis and two anonymous referees for comments on the manuscript.
Competing interests
The authors declare no competing financial interests.
Author contributions
I.V., Z.D., J.X. and R.D. conceived and designed the experiments, I.V. and Z.D. performed the experiments, I.V., Z.D., J.X. and R.D. performed the analyses, and I.V. and R.D. wrote the manuscript.
Funding
This research was funded by the Natural Sciences and Engineering Research Council of Canada, Canada Foundation for Innovation, and Ontario Innovation Trust (to R.D. and J.X.) and McMaster School of Graduate Studies (to Z.D.).
References
Archie, E. A. and Theis, K. R.(2011). Animal behaviour meets microbial ecology.
Anim. Behav.82, 425-436.
Barrows, W. M.(1907). The reactions of the Pomace fly, Drosophila ampelophila
Loew, to odorous substances. J. Exp. Zool.4, 515-537.
Bartelt, R. J., Schaner, A. M. and Jackson, L. L.(1985). cis-Vaccenyl acetate as an
aggregation pheromone in Drosophila melanogaster. J. Chem. Ecol.11, 1747-1756.
Becher, P. G., Bengtsson, M., Hansson, B. S. and Witzgall, P.(2010). Flying the fly:
long-range flight behavior of Drosophila melanogasterto attractive odors. J. Chem. Ecol.36, 599-607.
Becher, P. G., Flick, G., Rozpędowska, E., Schmidt, A., Hagman, A., Lebreton, S.,
Larsson, M. C., Hansson, B. S., Piškur, J. and Witzgall, P.(2012). Yeast, not fruit
volatiles mediate Drosophila melanogasterattraction, oviposition and development. Funct. Ecol.26, 822-828.
Begon, M.(1982). Yeasts and Drosophila. In The Genetics and Biology of Drosophila,
Vol. 3b (ed. M. Ashburner, H. L. Carson and J. N. Thompson), pp. 345-384. London: Academic Press.
Ben-Yosef, M., Aharon, Y., Jurkevitch, E. and Yuval, B.(2010). Give us the tools
and we will do the job: symbiotic bacteria affect olive fly fitness in a diet-dependent fashion. Proc. Biol. Sci.277, 1545-1552.
Borash, D. J., Gibbs, A. G., Joshi, A. and Mueller, L. D. (1998). A genetic
polymorphism maintained by natural selection in a temporally varying environment. Am. Nat.151, 148-156.
Boyd, R. F.(1988). General Microbiology. St Louis, MO: Times Mirror Mosby College.
Brieger, G. and Butterworth, F. M.(1970). Drosophila melanogaster: identity of male
lipid in reproductive system. Science167, 1262.
Broderick, N. and Lemaitre, B. (2012). Gut-associated microbes of Drosophila
melanogaster. Gut Microbes3, 1.
Brummel, T., Ching, A., Seroude, L., Simon, A. F. and Benzer, S.(2004). Drosophila
lifespan enhancement by exogenous bacteria. Proc. Natl. Acad. Sci. USA 101, 12974-12979.
Carton, Y. and David, J. R.(1985). Relation between the genetic variability of digging
behavior of Drosophilalarvae and their susceptibility to a parasitic wasp. Behav. Genet.15, 143-154.
Carton, Y., Bouletreau, M., Alphen, J. J. M. v. and Lenteren, J. C. v.(1986). The
Drosophilaparasitic wasps. In The Genetics and Biology of Drosophila(ed. M. Ashburner, H. L. Carson and J. N. Thompson), pp. 347-934. London: Academic Press.
Chandler, J. A., Lang, J. M., Bhatnagar, S., Eisen, J. A. and Kopp, A.(2011).
Bacterial communities of diverse Drosophilaspecies: ecological context of a host-microbe model system. PLoS Genet.7, e1002272.
Chu, C.-C., Spencer, J. L., Curzi, M. J., Zavala, J. A. and Seufferheld, M. J.(2013).
Gut bacteria facilitate adaptation to crop rotation in the western corn rootworm. Proc. Natl. Acad. Sci. USA110, 11917-11922.
Costa, J. T.(2006). The Other Insect Societies. Cambridge, MA: Harvard University
Press.
Crowley, S., Mahony, J. and van Sinderen, D.(2012). Comparative analysis of two
antifungal Lactobacillus plantarumisolates and their application as bioprotectants in refrigerated foods. J. Appl. Microbiol.113, 1417-1427.
Currie, C. R., Scott, J. A., Summerbell, R. C. and Malloch, D.(1999).
Fungus-growing ants use antibiotic-producing bacteria to control garden parasites. Nature
398, 701-704.
Danchin, E., Giraldeau, L.-A., Valone, T. J. and Wagner, R. H.(2004). Public
information: from nosy neighbors to cultural evolution. Science305, 487-491.
Davis, T. S., Crippen, T. L., Hofstetter, R. W. and Tomberlin, J. K.(2013). Microbial
volatile emissions as insect semiochemicals. J. Chem. Ecol.39, 840-859.
Demain, A. and Fang, A.(2000). The natural functions of secondary metabolites. In
History of Modern Biotechnology I, Vol. 69 (ed. A. Fiechter), pp. 1-39: Berlin: Springer.
Drasar, B. S. and Hill, M. J.(1974). Human Intestinal Flora. London: Academic Press.
Durisko, Z. and Dukas, R.(2013). Attraction to and learning from social cues in fruitfly
larvae. Proc. R. Soc. B280, 20131398.
Durisko, Z., Anderson, B. and Dukas, R.(2014). Adult fruit fly attraction to larvae
biases experience and mediates social learning. J. Exp. Biol.[Epub ahead of print] doi:10.1242/ jeb.097683.
Erkosar, B., Storelli, G., Defaye, A. and Leulier, F.(2013). Host–intestinal microbiota
mutualism: ‘learning on the fly’. Cell Host Microbe13, 8-14.
Ezenwa, V. O., Gerardo, N. M., Inouye, D. W., Medina, M. and Xavier, J. B.(2012).
Microbiology. Animal behavior and the microbiome. Science338, 198-199.
Faith, J. J., Guruge, J. L., Charbonneau, M., Subramanian, S., Seedorf, H., Goodman, A. L., Clemente, J. C., Knight, R., Heath, A. C., Leibel, R. L. et al.
(2013). The long-term stability of the human gut microbiota. Science341, 1237439.
Fleury, F., Ris, N., Allemand, R., Fouillet, P., Carton, Y. and Boulétreau, M.(2004).
Ecological and genetic interactions in Drosophila-parasitoids communities: a case study with D. melanogaster, D. simulansand their common Leptopilinaparasitoids in south-eastern France. Genetica120, 181-194.
Foster, J. A. and McVey Neufeld, K.-A.(2013). Gut-brain axis: how the microbiome
influences anxiety and depression. Trends Neurosci.36, 305-312.
Fredenhagen, A., Tamura, S. Y., Kenny, P. T. M., Komura, H., Naya, Y., Nakanishi,
K., Nishiyama, K., Sugiura, M. and Kita, H. (1987). Andrimid, a new peptide
antibiotic produced by an intracellular bacterial symbiont isolated from a brown planthopper. J. Am. Chem. Soc.109, 4409-4411.
Gilbert, D.(1980). Dispersal of yeasts and bacteria by Drosophilain a temperate
forest. Oecologia46, 135-137.
Gobbetti, M.(1998). The sourdough microflora: interactions of lactic acid bacteria and
yeasts. Trends Food Sci. Technol.9, 267-274.
Huser, A., Rohwedder, A., Apostolopoulou, A. A., Widmann, A., Pfitzenmaier, J.
E., Maiolo, E. M., Selcho, M., Pauls, D., von Essen, A., Gupta, T. et al.(2012).
The serotonergic central nervous system of the Drosophila larva: anatomy and behavioral function. PLoS ONE7, e47518.
Hutner, S. H., Kaplan, H. M. and Enzmann, E. V.(1937). Chemicals attracting
Drosophila. Am. Nat.71, 575-581.
Iyengar, B. G., Chou, C. J., Sharma, A. and Atwood, H. L.(2006). Modular neuropile
organization in the Drosophilalarval brain facilitates identification and mapping of central neurons. J. Comp. Neurol.499, 583-602.
Janzen, D. H.(1977). Why fruits rot, seeds mold, and meat spoils. Am. Nat.111,
691-713.
Laitila, A., Alakomi, H. L., Raaska, L., Mattila-Sandholm, T. and Haikara, A.(2002).
Antifungal activities of two Lactobacillus plantarumstrains against Fusariummoulds in vitroand in malting of barley. J. Appl. Microbiol.93, 566-576.
Lam, K., Babor, D., Duthie, B., Babor, E.-M., Moore, M. and Gries, G.(2007).
Proliferating bacterial symbionts on house fly eggs affect oviposition behaviour of adult flies. Anim. Behav.74, 81-92.
Leroy, P., Sabri, A., Verheggen, F., Francis, F., Thonart, P. and Haubruge, E.
(2011). The semiochemically mediated interactions between bacteria and insects. Chemoecology21, 113-122.
Man, D. J., Rogosa, M. and Sharpe, M. E.(1960). A medium for the cultivation of
lactobacilli. J. Appl. Microbiol.23, 130-135.
Mauch, A., Dal Bello, F., Coffey, A. and Arendt, E. K. (2010). The use of
Lactobacillus brevisPS1 toin vitroinhibit the outgrowth of Fusarium culmorumand other common Fusariumspecies found on barley. Int. J. Food Microbiol.141, 116-121.
Michener, C. D.(1974). The Social Behavior of the Bees. Cambridge, MA: Harvard
University Press.
Miller, J. H.(1992). A Short Course in Bacterial Genetics: A Laboratory Manual And
Handbook For Escherichia Coli And Related Bacteria. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Nassif, C., Noveen, A. and Hartenstein, V. (2003). Early development of the
Drosophilabrain: III. The pattern of neuropile founder tracts during the larval period. J. Comp. Neurol.455, 417-434.
Pauls, D., Selcho, M., Gendre, N., Stocker, R. F. and Thum, A. S.(2010). Drosophila
larvae establish appetitive olfactory memories via mushroom body neurons of embryonic origin. J. Neurosci.30, 10655-10666.
Ridley, E. V., Wong, A. C. N., Westmiller, S. and Douglas, A. E.(2012). Impact of the
resident microbiota on the nutritional phenotype of Drosophila melanogaster. PLoS ONE7, e36765.
Robacker, D. C., Lauzon, C. R. and He, X. (2004). Volatiles production and
attractiveness to the Mexican fruit fly of Enterobacter agglomeransisolated from apple maggot and Mexican fruit flies. J. Chem. Ecol.30, 1329-1347.
Robacker, D. C., Lauzon, C. R., Patt, J., Margara, F. and Sacchetti, P.(2009).
Attraction of Mexican fruit flies (Diptera: Tephritidae) to bacteria: effects of culturing medium on odour volatiles. J. Appl. Entomol.133, 155-163.
Robinson, G. E., Fernald, R. D. and Clayton, D. F. (2008). Genes and social
behavior. Science322, 896-900.
Rohlfs, M. and Hoffmeister, T. S.(2004). Spatial aggregation across ephemeral
resource patches in insect communities: an adaptive response to natural enemies? Oecologia140, 654-661.
Rohlfs, M., Obmann, B. and Petersen, R.(2005). Competition with filamentous fungi
and its implication for a gregarious lifestyle in insects living on ephemeral resources. Ecol. Entomol.30, 556-563.
Rozen, D. E., Engelmoer, D. J. P. and Smiseth, P. T.(2008). Antimicrobial strategies
in burying beetles breeding on carrion. Proc. Natl. Acad. Sci. USA105, 17890-17895.
Ruebenbauer, A., Schlyter, F., Hansson, B. S., Löfstedt, C. and Larsson, M. C.
(2008). Genetic variability and robustness of host odor preference in Drosophila melanogaster. Curr. Biol.18, 1438-1443.
Ruiz-Barba, J. L., Cathcart, D. P., Warner, P. J. and Jiménez-Díaz, R.(1994). Use of
Lactobacillus plantarum LPCO10, a bacteriocin producer, as a starter culture in Spanish-style green olive fermentations. Appl. Environ. Microbiol.60, 2059-2064.
Sarin, S. and Dukas, R. (2009). Social learning about egg-laying substrates in
The Journal of Experimental Biology
Savage, D. C.(1977). Microbial ecology of the gastrointestinal tract. Annu. Rev.
Microbiol.31, 107-133.
Schnürer, J. and Magnusson, J. (2005). Antifungal lactic acid bacteria as
biopreservatives. Trends Food Sci. Technol.16, 70-78.
Schulz, S. and Dickschat, J. S. (2007). Bacterial volatiles: the smell of small
organisms. Nat. Prod. Rep.24, 814-842.
Scott, J. J., Oh, D.-C., Yuceer, M. C., Klepzig, K. D., Clardy, J. and Currie, C. R.
(2008). Bacterial protection of beetle-fungus mutualism. Science322, 63.
Sharon, G., Segal, D., Ringo, J. M., Hefetz, A., Zilber-Rosenberg, I. and
Rosenberg, E.(2010). Commensal bacteria play a role in mating preference of
Drosophila melanogaster. Proc. Natl. Acad. Sci. USA107, 20051-20056.
Sokolowski, M. B.(2010). Social interactions in ‘simple’ model systems. Neuron65,
780-794.
Spencer, W. P.(1950). Collection and laboratory culture. In Biology of Drosophila(ed.
M. Demerec). Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Stamps, J. A., Yang, L. H., Morales, V. M. and Boundy-Mills, K. L. (2012).
Drosophilaregulate yeast density and increase yeast community similarity in a natural substrate. PLoS ONE7, e42238.
Stensmyr, M. C., Dweck, H. K., Farhan, A., Ibba, I., Strutz, A., Mukunda, L., Linz,
J., Grabe, V., Steck, K., Lavista-Llanos, S. et al.(2012). A conserved dedicated
olfactory circuit for detecting harmful microbes in Drosophila. Cell151, 1345-1357.
Stökl, J., Strutz, A., Dafni, A., Svatos, A., Doubsky, J., Knaden, M., Sachse, S.,
Hansson, B. S. and Stensmyr, M. C. (2010). A deceptive pollination system
targeting drosophilids through olfactory mimicry of yeast. Curr. Biol.20, 1846-1852.
Storelli, G., Defaye, A., Erkosar, B., Hols, P., Royet, J. and Leulier, F.(2011).
Lactobacillus plantarum promotes Drosophila systemic growth by modulating hormonal signals through TOR-dependent nutrient sensing. Cell Metab.14, 403-414.
Sturtevant, A. H.(1921). The North American Species of Drosophila. Washington, DC:
Carnegie Institution of Washington.
Wertheim, B., Marchais, J., Vet, L. E. M. and Dicke, M.(2002). Allee effect in larval
resource exploitation in Drosophila: an interaction among density of adults, larvae, and micro-organisms. Ecol. Entomol.27, 608-617.
Wertheim, B., van Baalen, E.-J. A., Dicke, M. and Vet, L. E. M.(2005).
Pheromone-mediated aggregation in nonsocial arthropods: an evolutionary ecological perspective. Annu. Rev. Entomol.50, 321-346.
Wilson, E. O.(1971). The Insect Societies. Cambridge, MA: Harvard University Press.
Wilson, E. O.(1975). Sociobiology: The New Synthesis.Cambridge, MA: Harvard
University Press.
Wong, C. N. A., Ng, P. and Douglas, A. E.(2011). Low-diversity bacterial community
in the gut of the fruitfly Drosophila melanogaster. Environ. Microbiol.13, 1889-1900.
Zhu, J., Park, K.-C. and Baker, T. C.(2003). Identification of odors from overripe mango