Eiszeitalter und Gegenwart Band 12 Seite gg-124 Öhringen/Württ. ib. Januar ic,6l
Über d i e G r ö ß e d e r p l e i s t o z ä n e n Pferde d e r C a b a l l u s - G r u p p e
in Europa u n d N o r d a s i e n
V o n O. S I C K E N B E R G , H a n n o v e r Mit 5 Abbildungen im Text
Z u s a m m e n f a s s u n g : Neben Schädel- und Zahnbau ist die Körpergröße und die Pro-portionierung des metapodialen Gliedmaßenabschnittes ein sehr wichtiges Merkmal. Für beide wer den Beziehungen zu bestimmten Temperaturverhältnissen des jeweiligen Lebensraumes und zu dessen landschaftlichen Charakter wahrscheinlich gemacht. Ihre Ermittlung vermag auch wertvolle Hinweise in taxionomischen und stammesgeschichtlichen Fragen zu geben, als Merkmale allein genommen können sie aber nicht als Grundlage einer taxionomischen Ordnung dienen, da An gehörige verschiedener Formenkreise annähernd gleiche Körpergröße und -proportionierung zei gen. Im Gegensatz zu den Eseln, Halbeseln, Zebras, aber auch zum Przewalski-Pferd und wahr scheinlich auch zu den Pferden des Equus sanmeniensis — Formenkreises sind sämtliche pleisto zänen Pferde der Untergattung Caballus in der eurosibirischen Region unabhängig von der Körpergröße „schwer" gebaut, a. h. sie besitzen kurze, verhältnismäßig plumpe Metapodien. Sie waren Wald-, Bergland oder Tundrabewohner und nicht in trockenwarmen oder -kalten, ebenen Steppengebieten beheimatet. D a s gleiche dürfte auch für die altquartären Pferde des „Allohippus"-Formenkreises anzunehmen sein. Eine Gruppierung nach drei Größenkategorien kann vorgenom men werden (Groß-, Mittel- und Kleinpferde). Groß- und Kleinpferde gehören möglicherweise jeweils zu einer taxionomischen Einheit, die Mittelpferde sind dagegen ein Formengemenge. Zu mindest von der Holstein-Warmzeit an erscheinen in Mittel- und Osteuropa, sowie in Nordasien große Pferde in den Tiergesellschaften der wärmeren Phasen, das Auftreten von Klein- und Kleinst pferden steht im Zusammenhang mit dem Vorherrschen ungünstiger Klimabedingungen.
S u m m a r y . Apart from skull- and tooth-structure, the size of body and the proportioning of the metapodial section of limbs are very important characteristics. Both criteria are said to be related with certain conditions of temperature of the respective living area as well as with its specific scenery. Moreover, their investigation can yield valuable indications as to taxionomy and phylogeny. Merely taken as characteristics, they do not suffice to serve as a base for a taxio-nomic classification, since members of different type-series show approximately the same size and proportioning of body. In opposition to donkeys, zebras and even to the Przewalski horse as well as probably to the horses of the Equus sanmeniensis series, all pleistocene horses of the Caballus subgenus in the eurosiberian region are — independant of the size of body — "heavily" built, that means they have got short, relatively clumsy metapodial bones. They were dwellers in woods, in highlands or in the tundra and not in dry-warm or dry-cold plain steppes. The same may be assumed for the old-quaternary horses of the "Allohippus" series. A classification into three cate gories can be carried through (large-, medium- and small-sized horses). Large- and small-sized horses may possibly belong to one taxionomic unity each. The mediumsized horses on the other hand represent a product of an intermingling of types. Beginning with the Holstein-Warm-Period at the latest, in Central and Eastern Europe as well as in Northern Asia, large horses occur in the association of animals of the warmer phases. The occurrence of small and smallest horses is connected with the prevailing of unfavourable climatic conditions.
I
Wohl a l l e Bearbeiter sind sich über den nach wie vor unbefriedigenden S t a n d unserer Kenntnisse über die pleistozänen Equiden einig. D a wir uns nicht einmal über T a x i o n o m i e und N o m e n k l a t o r i k hinreichend klar sind, k a n n nicht erwartet werden, wir vermöchten bestimmte A u s s a g e n über die phyletischen Beziehungen, über die stratigraphische Reich weite einzelner Formen, über Lebensraum u n d Lebensgewohnheiten, über die g e o g r a phische Verteilung und über vielleicht eingetretene Arealverschiebungen zu treffen. D e r Ursachen, die dafür verantwortlich zu machen sind, sind mehrere. F u n d e ganzer Skelette oder größerer zusammenhängender Skeletteile, j a selbst nur gut erhaltener Schädel sind selten. Einigermaßen b e k a n n t sind nur die Equidenreste bestimmter Fundorte in Deutsch land, Schweiz und Österreich sowie in Frankreich, E n g l a n d u n d den N i e d e r l a n d e n . In den letztgenannten drei L ä n d e r n erfolgten in letzter Zeit allerdings hauptsächlich nur
Veröffentlichungen über altquartäre Pferde. In Schweden sind es wieder nur die holozänen Wildpferde, die, allerdings sehr gründlich, untersucht wurden. D e r gesamte Süden und Südosten E u r o p a s stellt hinsichtlich der Pferde praktisch eine terra incognita dar. D a n k der umfassenden Zusammenstellung GROMOVA'S u n d älterer russischer Arbeiten liegen einige Angaben über die Pferde des gewaltigen eurasiatischen R a u m e s nördlich der G e -birgsländer vor. D a b e i fehlen monographische Bearbeitungen aus neuerer Zeit, sei es aus regionaler oder stratigraphischer Sicht, allenthalben. D i e Beurteilung fossilen M a t e r i a l s w i r d weiter durch den U m s t a n d sehr erschwert, d a ß wir über A u s m a ß und A r t der V a r i a b i l i t ä t der Hartteile der lebenden W i l d p f e r d e einschl. des Przewalski-Pferdes, ge wonnen an einem genügend großen Material, keineswegs zufriedenstellend unterrichtet sind. Ungenügende Materialkenntnis allein bedingt aber nicht die Schwierigkeiten. D i e gesamten Pferde neigen vom Beginn ihrer Stammesgeschichte k a u m d a z u , ihren B a u p l a n wesentlich a b z u ä n d e r n . Eine gewisse Starrheit ist unverkennbar, die sich natürlich beson ders im L a u f e ihrer späteren, also jungtertiären und quartären Geschichte bemerkbar macht. D a s Formbildungsvermögen reicht im Gegensatz zu anderen, plastischeren Stämmen nicht hin, aus dem Grundstock in morphologischer u n d biologischer Hinsicht eigenständige Z w e i g e hervorzutreiben, es erschöpft sich sozusagen in einer vielfältigen A b w a n d l u n g des „ T y p u s " . Es ist groß genug, innerhalb der vergleichsweise wenigen, gut abgrenzbaren Formenkreise eine offenbar große A n z a h l taxionomischer Kategorien niederer und nied rigster R a n g o r d n u n g zu schaffen, z u schwach aber, um es noch einmal zu sagen, entschei dend über den R a h m e n des gegebenen hinauszuwachsen oder ihn g a r zu sprengen. D i e unverkennbare „ N e i g u n g " der späten Equiden, a u f offensichtliche Änderungen der kli matischen und d a m i t landschaftlichen Bedingungen ihrer Lebensräume höchstens nur mit Bildung von K l e i n - und Kleinstarten zu antworten, steht damit in Zusammenhang. Diese richtig zu erkennen und abzugrenzen, ihre Entstehung aus endogenen oder exogenen U r sachen heraus verstehen zu wollen, stellt daher den Palaeozoologen vor äußerst schwierige Probleme. W a s er an seinem und a m Material a u s der Gegenwart feststellen kann, ist dies: Durchgreifende, klar faßbare strukturelle Unterschiede im Schädelbau, in der B e -zahnung und bei den übrigen Skelettelementen fehlen selbst zwischen den H a u p t f o r m e n kreisen, die äußere Konfiguration des Schädels und die Proportionierung seiner Teile w i r d aber erheblich variiert, das Gebiß zeigt zwar v o n Population zu P o p u l a t i o n ein jeweils eigenes G e p r ä g e , nur sehr wenige M e r k m a l e scheinen aber einigermaßen gattungs- oder artkonstant zu sein. A m auffälligsten sind die Unterschiede in der Gesamtgröße der T i e r e und die Proportionierung der einzelnen Teile des B e w e g u n g s a p p a r a t e s .
D a s Ziel der nachstehenden Studie ist, die G r ö ß e der Pferde des Pleistozäns E u r o p a s und des nördlichen Asiens in Beziehung auf R a u m , Zeit und Umweltsbedingungen zu untersuchen.
D a fossile Belege dieses Alters aus der Z e b r a - H a l b e s e l - und E s e l - G r u p p e entweder sehr spärlich sind oder überhaupt fehlen, gebot sich v o n selbst die Beschränkung auf die
Caballus-Gmppe, d. h. auf die G a t t u n g Equus s. str., wobei vor allem deren
jungpleisto-z ä n e Vertreter berücksichtigt wurden. D i e altpleistojungpleisto-zänen Formen bleiben etwas außer halb der Betrachtung, einmal weil ihre Zugehörigkeit zur Caballus-Gruppe bei den A n gehörigen des „Stenonis-Kreises" im weiteren Sinne zweifelhaft ist, z u m anderen, weil auswertbare R e s t e noch seltener als im jüngeren Q u a r t ä r sind. D a von den E x t r e mitätenknochen unter fossilem M a t e r i a l die M e t a p o d i e n am häufigsten sind und den besten Erhaltungszustand zeigen, wurden diese allein der Untersuchung zu Grunde gelegt. Vorausgesetzt w i r d dabei allerdings, daß die L ä n g e von Mittelhand und -fuß R ü c k schlüsse auf die Gesamtgröße erlaubt, w o z u allerdings bemerkt werden muß, daß bei g e wissen Formen bei eintretender „ V e r p l u m p u n g " (Längenabnahme bei gleichbleibender Breite) vorzugsweise nur der m e t a p o d i a l e Extremitätenabschnitt betroffen wird ( H O F -S T E T T E R 1952). D i e plumpen K l e i n p f e r d e waren daher in Wirklichkeit etwas größer als
Pleistozäne Pferde der Caballus-Gruppe in Europa und Nordasien 101
m a n nach den Längen ihrer Metapodien vermeinen könnte. Z u r Feststellung der jeweiligen Größenverhältnisse dienten die größte L ä n g e und die Breite in der D i a p h y s e n -mitte der einzelnen E x e m p l a r e . D i e Angaben w u r d e n fast ausnahmslos der Literatur ent nommen, besonders aus den Arbeiten von S C H W A R Z 1927 und G R O M O V A 1949, die aller dings selbst wieder zu einem guten Teil auf die Messungen früherer Bearbeiter zurück gehen. Ungenauigkeiten der verschiedensten A r t schleichen sich d a m i t natürlich ein und beeinträchtigen die Ergebnisse (verschiedene Meßmethoden, H e r e i n n a h m e der Meßzahlen jugendlicher, als solche aber nicht gekennzeichneter Individuen in die Maßtabellen, m a n gelnde Berücksichtigung des Erhaltungszustandes usw.). Dies mußte in K a u f genommen werden, um überhaupt verfahren zu können, wenn solche Fehler auch angesichts der s p ä r lichen Stückzahl schwerer wiegen als bei reicher Materialfülle. D a s Gesamtbild m a g d a durch an der einen oder anderen Stelle vielleicht etwas verzerrt, bis zur Unkenntlichkeit verfälscht aber nicht werden. Auch die sexuellen Unterschiede, die sich zumindest beim H a u s p f e r d bei den Metapodien feststellen lassen ( K U N K A I T I S nach L U N D H O L M 1949) konn ten naturgemäß nicht berücksichtigt werden. D i e Maßzahlen w u r d e n in D i a g r a m m e ein getragen, welche zunächst die Position jedes einzelnen Exemplares nach Gesamtlänge und Diaphysenmitten-Breite erkennen lassen. A u ß e r d e m wurde von den jeweils extremsten Werten ausgehend versucht, Rechtecksfelder z u konstruieren, um den Variationsbereich bestimmter G r u p p e n graphisch darzustellen. D u r c h die Überbetonung der Breite werden z w a r die Proportionen verzerrt, zugleich bestimmte Verhältnisse aber auch verdeutlicht. Selbstredend bedeutet ein solches Verfahren nur eine sehr grobe Annäherung an die W i r k lichkeiten, d a die Stückanzahl, welche diesen Konstruktionen z u G r u n d e liegt, jeweils sehr verschieden und in keinem der Fälle ausreichend genug ist. V o n keiner echten P o p u lation — die ja sowieso nur bei rezenten Formen untersuchbar ist — liegt genügend großes Material vor, das erlauben würde, Größe und Konfiguration des Variationsfeldes festzu stellen und die Anordnung der einzelnen E x e m p l a r e in diesem einwandfrei zu erkennen. Aus theoretischen Erwägungen heraus kann als sicher gelten, d a ß sich die den einzelnen Exemplaren entsprechenden M a r k e n nicht gleichmäßig über das F e l d verteilen, sondern sich entlang einer oder zweier Achsen, — bei Bestehen merklicher sexueller Unterschiede, — anordnen. E i n Überschneiden der einzelnen F e l d e r in der graphischen Darstellung bedeu tet daher noch keineswegs, daß sich die Werte realiter überschneiden, dies darf erst bei bedeutenderem Deckungsgrade zweier Felder angenommen werden.
Aus den gleichen Gründen wurde auch auf eine rechnerisch-variationsstatistische B e handlung der Materialien verzichtet. Über d a s , w a s durch M a ß a n g a b e n genau belegbar ist, wurde nicht hinausgegangen, denn die in der Literatur sehr häufigen, allgemein ge haltenen Angaben wie „große F o r m " , „kleines P f e r d " sind zu subjektiv gefärbt, um viel von N u t z e n zu sein.
Im Rahmen der vorliegenden Studie bot sich bei dem gegenwärtigen Stand der D i n g e keine andere Möglichkeit, als sich der konventionellen T a x i o n o m i e und N o m e n k l a t o r i k zu bedienen. D a die Auffassungen darüber, w a s als Art und w a s als Unterart anzusehen sei, bei den einzelnen Autoren weit auseinandergehen, die Zeit für Entscheidungen auch noch nicht reif ist, erscheinen die einzelnen, bisher benannten F o r m e n durchgehend als Arten. V o n den verschiedenen Untergattungen der Großgattung Equus sind in den U n tersuchungskreis nur folgende einbezogen: Caballus im Sinne D I E T R I C H S , Asinus,
Hemio-nus und Onager (hier zusammengefaßt zur HemioHemio-nus-Gruppe), Hippotigris, Dolicho-hippus, Zebra und Quagga (hier zusammengefaßt zur „Hippotigris"-Gruppe), Amer-bippus und schließlich Allohippus, die letztgenannte, derzeit noch nicht genauer zu kenn
zeichnende und abgrenzbare Untergattung gebraucht als Sammelbezeichnung für alle alt-quartären E q u i d e n , die nicht eindeutig zur U n t e r g a t t u n g Caballus oder zu einer der anderen aufgeführten Untergattungen gehören (SIMPSON 1945, H O F S T E T T E R 1952, D I E T RICH 1950). D i e oben genannte Bemerkung bezüglich T a x i o n o m i e und N o m e n k l a t o r i k
bezieht sich v o r allem auf die Untergattung Caballus. Unter ihren zahlreichen „ A r t e n " scheint mir nur als selbständige taxionomische Einheit das Pferd a u s den Mosbacher S a n -den ( = C.mosbacbensis v. R E I C H E N A U ) gesichert, alles andere erscheint derzeit sowohl taxionomisch, wie nomenklatorisch unsicher (E. germanicus N E H R I N G , E. steinheimensis
v . R E I C H E N A U , E. taubachensis v . R E I C H E N A U , E. chosaricus G R O M O V A , E. missi M . P A V L . , E.woldrichi A N T O N I U S USW.): v o n einer wirklichen Ordnung sind w i r noch weit entfernt.
M a n wird verstehen, daß angesichts dieser Sachlage das gewählte T h e m a nur m i t den größten Vorbehalten angegangen werden kann. E i n derartiger Versuch darf überhaupt nur in der Zuversicht unternommen werden, d a ß jede kritische Materialsichtung im Hinblick auf eine bestimmte F r a g e v o n N u t z e n sein wird, wenn nicht von anderem, so doch wenigstens v o n dem einen, diese F r a g e schärfer und enger z u fassen, um andere U n tersuchungen anzuregen und d a m i t die Möglichkeit z u eröffnen, eine der Wirklichkeit ge mäße A n t w o r t z u finden. Dies m ö g e bedacht werden, wenn hier entschieden mehr P r o bleme gestellt als gelöst erscheinen, w a s mit gewissem Recht als enttäuschend empfunden werden m a g . E i n umfassender Versuch, an H a n d v o n fossilem M a t e r i a l (Metapodien) den Größenverhältnissen der pleistozänen Equiden E u r o p a s nachzugehen, um damit z u g e wissen Schlußfolgerungen z u k o m m e n , wurde erst einmal gemacht (SCHWARZ 1 9 2 7 ) . E r w a r ziemlich berechtigter K r i t i k ausgesetzt, d a chronologische D a t e n nicht entsprechend berücksichtigt u n d die Verschiedenartigkeit der oekologischen Verhältnisse vernachlässigt wurden. D i e Ergebnisse, namentlich im taxionomischen Bereich, ruhen daher zwangsläufig auf mehr als schwankem B o d e n .
II
D i e H a u p t g r u p p e n d e r p l e i s t o z ä n e n u n d h o l o z ä n e n E q u i d e n ( D i a g r a m m l a u n d l b )
M a t e r i a l : M e t a c a r p a l e
1. Caballus-Gruppe einschl. Allohippus:
U m f a ß t nahezu das gesamte veröffentlichte, mit einiger Sicherheit auf die Caballus-G r u p p e beziehbare pleistozäne Material E u r o p a s und N o r d a s i e n s (einschl. E. san
meniensis T E I L H . D E J A R D . et Piv., ~ 2 0 0 E x e m p l . ) sowie E. przewalski P O L . ( 8 E x
empl.).
2 . Hemionus-Gruppe:
H. bemionus P A L L . U. H. onager BODO-, teils fossil aus N - A s i e n , teils subfossil u n d
rezent, insges. 1 1 E x e m p l . 3 . v4«'w«s-Gruppe:
Asinus asinus L . , A. somaliensis, A. hydruntinus R E G . , insgesamt 1 3 Exempl. E i n v o n
P O M E L veröffentlichtes E x e m p l a r (n. GROMOVA) aus dem N e o l i t h i k u m N o r d a f r i k a s wurde nicht berücksichtigt; die extreme Schlankheit dieses M t c w ü r d e das F e l d b e deutend nach links erweitern. E b e n s o wurde ein sehr großes E x e m p l a r v. A. hydrun
tinus, ( F u n d l o k . T i r a s p o l ) , dessen Zugehörigkeit z . A. h. m. E . fraglich ist, nicht b e
rücksichtigt.
4. Hippotigris-Gtuppe (Hippotigris s. 1.):
Hippotigris mauretanicus P O M . , H. boehmi, H. burchelli, H. zebra, insges. 4 E x e m p l .
5. Amerhippus-Gruppe:
A. Andium W A G N . - B R A N C O , A. neogaeus L U N D , A. curvidens O W E N , insges. 5 1 E x e m p l .
M e t a t a r s a l e
1. Caballus-Gruppe, wie oben, ~ 2 5 0 E x e m p l a r e .
2 . Hemionus-Gruppe, wie oben, insgesamt 1 1 E x e m p l a r e aber z . T . v o n anderen F u n d orten als die M e t a c a r p a l i a ) .
Pleistozäne Pferde der Caballus-Gruppe in Europa und Nordasien 103 10 Breite in mm Caballus - Gruppe (einschl. "Allohippus J MU/a/ü/U/ü- £ Przewa/sh, Hemionus-Gruppe Asinus -Gruppe '//////////ss///// A, hydruntinus Zebra-Gruppe — o — o — Amerhippus-Gruppe
Abb. 1. Die Hauptgruppen der pleistozänen und rezenten Pferde (Europa, Asien, Afrika, Südamerika), a: Metacarpale, b : Metatarsale.
3 . v4s:'tt«s-Gruppe:
Wie oben, insgesamt 1 1 ; auch hier wurden nur die mitteleuropäischen E x e m p l a r e v o n
A. hydruntinus berücksichtigt, d a bei den größerdimensionierten osteuropäischen V e r
-tretern Fehlbestimmungen vorliegen können (Verwechslung mit Angehörigen der
Hemionus-Gruppe!).
4. Hippotigris-Gruppe:
H. mauretanicus P O M . , H. oldowayensis H O P W . , H. boehmi, H. burchelli, H. zebra,
insgesamt 5 E x e m p l a r e . 5. Amerhippus-Gruppe:
A. Andium W A G N . - B R A N C O , A. Martinei S P I L L M . , A. Santae-Elenae S P I L L M . , A. neo-gaeus L U N D , A. insulatus C . A M E G H . , A. curvidens O W E N , insgesamt 55 E x e m p l .
Die D i a g r a m m e wurden aufgestellt, um die größenmäßigen Beziehungen zwischen den wichtigsten altweltlichen Untergattungen zu ermitteln. D i e nordamerikanischen Q u a r t ä r pferde wurden also nicht mit einbezogen, die südamerikanischen Vertreter der U n t e r
-gattung Amerbippus erscheinen aber zu bestimmten Vergleichszwecken in den D i a g r a m men.
Folgende Feststellungen ergeben sich:
1. D i e einzelnen Felder überschneiden sich z w a r , die einzelnen G r u p p e n als solche kön nen aber mit hinreichender Deutlichkeit als selbständige Einheiten erkannt werden. 2. Zwei H a u p t g r u p p e n zeichnen sich a b : D i e eine umfaßt kleine bis mittelgroße Formen
mit schlanken Metapodien; d a z u zählen die Hemionus-, die Asinus- und die Hippoti-g r i s - G r u p p e , am schlanksten sind die Esel, p l u m p e r die Zebras, die Halbesel v e r m i t teln, sind aber größer als die beiden (Asinus hydruntinus reiht sich in das Hemionus-F e l d ein. D i e sichere Bestimmung einzelner isolierter Metapodien von A. hydruntinus oder H. spec, scheint mir daher auf unüberwindliche Schwierigkeiten zu stoßen). Zur anderen H a u p t g r u p p e gehören die echten Pferde einschließlich Allohippus u n d die Arten der G a t t u n g Amerbippus. E s sind sehr kleine bis sehr große Pferde mit mittel schlanken bis sehr plumpen Metapodien (über das Przewalski-Pferd s. u.).
3. D i e Proportionsverhältnisse sind bei den Metatarsalien „gefestigter", bei den M e t a -c a r p a l i a ist offenbar im allgemeinen der S p i e l r a u m für den S-chlankheitsgrad größer (Ausnahme E. przewalski?).
III
U m für den weiteren Untersuchungsgang einen Ausgangspunkt zu finden, mußte ver sucht werden, an H a n d eines einigermaßen homogenen Materials über U m f a n g u n d Art der Größenvariabilität zumindest einiger weniger der in F r a g e kommenden F o r m e n be stimmtere Vorstellungen zu gewinnen. "Wenn eingangs erklärt w u r d e , es ließen sich so gut wie keine gesicherten taxionomischen Einheiten aufstellen, so soll dies nicht bedeuten, es hätte solche überhaupt nicht gegeben. I m Gegenteil, es besteht aller G r u n d zur A n nahme, daß die Untergattung Caballus unter allen die formenreichste war und in eine große Anzahl zumindest von „guten" Unterarten und Rassen zerfiel. Ihre E r k e n n u n g und A b g r e n z u n g bereitet aber derzeit fast unüberwindliche Schwierigkeiten, aus G r ü n d e n , die gleichfalls schon eingangs dargelegt wurden. Entweder handelt es sich beim fossilen Material um Einzelfunde oder das Material ist z w a r umfangreicher, aber wahrscheinlich häufig uneinheitlich, denn bei fast allen älteren Aufsammlungen wurden die Knochen eines Fundortes nicht genügend nach den einzelnen Fundschichten getrennt und dadurch nicht selten verschiedenartiges vermengt. D i e Fundlager, Höhlenablagerungen, T e r r a s s e n schotter und Lößmassen sind meist, wie sich immer mehr herausstellt, recht k o m p l e x e r N a t u r und nur selten zeitliche und ökologische Einheiten. Auch wenn eine wirkliche E i n heit gegeben erscheint, so muß in Rechnung gesetzt werden, daß möglicherweise zu gleicher Zeit zwei oder mehr Formen den gleichen R a u m besetzten, wie m a n dies von den heutigen Zebras kennt. U m aber mit Sicherheit ein derartiges Kollektiv in seine Bestandteile auf lösen zu können, d a z u bedarf es eines größeren Materials, als in der Regel von eurasiati-schen F u n d p l ä t z e n zur Verfügung steht. I m G r u n d e genommen entspricht keine der bis herigen Aufsammlungen, von einer Ausnahme abgesehen, den anzulegenden M a ß s t ä b e n , eine wenig ermutigende Feststellung, die aber um der Klarheit willen notwendig erscheint. U m nichts besser steht es bei dem rezenten Vertreter der Untergattung, bei E. przewalski P O L . , das bekanntlich nur noch in wenigen E x e m p l a r e n als W i l d f o r m v o r k o m m t u n d d a her in keiner S a m m l u n g der Welt in reichen Serien vertreten ist. Unsere K e n n t n i s be schränkt sich daher auf das, w a s die Untersuchung nur weniger E x e m p l a r e ergibt. Außer dem kann eine wenn auch geringe Beimengung v o n H a u s p f e r d b l u t bei den heute lebenden Wildpferdbeständen nicht g a n z ausgeschlossen werden ( M O H R 1 9 5 9 ) . Ein anderes sub rezentes W i l d p f e r d , der T a r p a n (E. gmelini A N T . ) kann zwar auf G r u n d verschiedener E r w ä g u n g e n als ehemals existent angenommen werden, als taxionomische K a t e g o r i e ist es aber das Ergebnis einer Konstruktion, hinter der eine wahrscheinliche, aber keineswegs
Pleistozäne Pferde der Caballus-Gruppe in Europa und Nordasien 105
gesicherte und zweifelsfreie R e a l i t ä t s t e h t1) , auf deren in Sammlungen befindlichen B e legen bei Untersuchungen zurückgegriffen werden k a n n . I n den T a b e l l e n bei GROMOVA ( 1 9 4 9 ) ist daher der T a r p a n nur m i t einem E x e m p l a r vertreten.
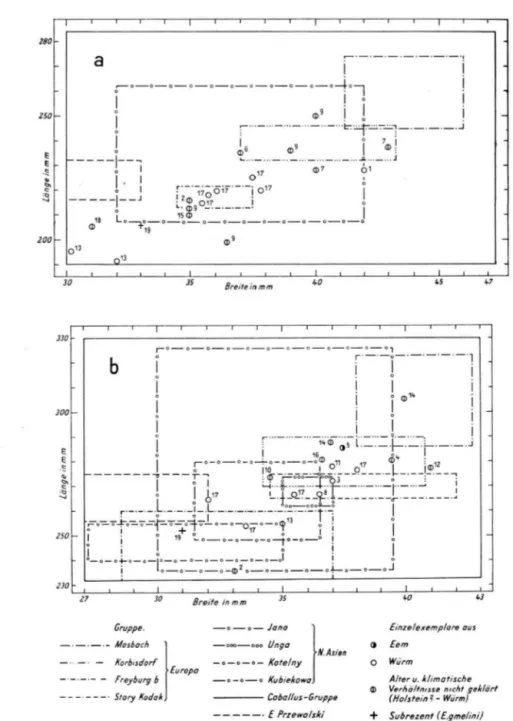
Bei der geschilderten L a g e blieb mithin kein anderer Weg offen, a l s d a s Material ver schiedener Fundpunkte, soweit es der leider immer sehr bescheidene U m f a n g überhaupt noch zuläßt, z u m A u f b a u des D i a g r a m m e s (2a u. b ) z u verwerten. Ü b e r die chronologi schen und klimatischen Daten der einzelnen Fundlokalitäten b z w . - k o m p l e x e gibt der A n h a n g S. 120) Aufschluß, auf den auch für die später folgenden Ausführungen zu ver weisen ist.
Equus przewalski P O L . , Rezent;
Stückzahl: M e t a c a r p a l i a : 8, M e t a t a r s a l i a : 8 (nach GROMOVA 1949).
E. gmelini A N T . , Subrezent;
Stückzahl: M t c : 1, M t t : 1 (n. GROMOVA 1949).
E.spec, Frey burg a. d . U . : (kleine F o r m = Freyburg b) W ü r m ; S t ü c k z a h l : M t c : 5, M t t : 8 (nach S C H W A R Z 1927 u. v . R E I C H E N A U 1 9 1 5 ) . E.spec. Freyburg a. d . U . : (größere F o r m = F r e y b u r g a ) , W ü r m ;
Stückzahl Mtc 2, M t t 5 (nach SCHWARZ 1927).
N . B . I m Freyburger L ö ß kommen offensichtlich zwei verschiedengroße Pferde vor, wie aus dem D i a g r a m m ersichtlich.
E.spec. Solutre, S p ä t w ü r m ;
Stückzahl: M t c : 4 0 (Länge) 7 (Breite), M t t : 44 ( L ä n g e ) , 4 (Breite)
( a u s G R O M O V A 1 9 2 7 u. n. V I R E T 1 9 5 4 ) .
V o n allen Materialien ist hier zweifelsohne die größte Einheitlichkeit in Bezug auf Zeit-S t e l l u n g und K l i m a gegeben. D a in Mitteleuropa im Zeit-S p ä t w ü r m aber mindestens zwei ver schiedene Pferde auftreten, ist die taxionomische Einheitlichkeit fraglich. Leider sind in der Literatur nicht Einzel-, sondern nur Grenzwerte für die Metapodien angegeben, eine Ent scheidung, ob eine oder zwei GrößengruDpen vorhanden sind, kann erst durch eine neue umfassende Bearbeitung erfolgen.
F.. spec. Körbisdorf, R i s s ;
Stückzahl: M t c 8, M t t : 14 (nach SCHWARZ 1927)
Eine gewisse stratigraphische und klimatische Einheitlichkeit der Fundschichten darf ver mutet werden. E i n e morphologische A n a l y s e der K ö r b i s d o r f e r Equiden fehlt.
E. mosbachensis v . R E I C H E N A U , Mosbacher Sande, J u n g . Altpleistozän;
Stückzahl: M t c 9, M t t 1 6 (n. SCHWARZ 1927 u. v. R E I C H E N A U 1915). D i e Mosbacher S a n d e bilden weder eine Alters- noch eine oekologische Einheit. D i e meisten Pferdereste stammen aber aus dem H a u p t l a g e r , so d a ß eine gewisse Einheitlichkeit angenommen werden darf, was auch durch die morphologischen Befunde gestützt wird (v. R E I C H E N A U 1915).
E. stenonis C O C H . , S t . Vallier, A l t . Altpleistozän,
Stückzahl: M t c : 15, M t t : 32 (n. V I R E T 1954).
N a c h den geologischen Verhältnissen der F u n d l o k a l i t ä t und nach d e m morphologischen Befund darf Einheitlichkeit angenommen werden; E. stenonis wird aber an dieser F u n d stelle noch von einem größeren Pferd, E. bressanus V I R E T begleitet.
E. „sanmeniensis" T . D E J A R D . et P i v . , China, Altpleistozän;
S t ü c k z a h l : M t c : 2 2 , M t t : 29 (aus G R O M O V A 1949 u n d nach V I R E T 1 9 5 4 ) .
Die weite geographische Streuung der Fundorte und die große S p a n n e des zeitlichen Auf tretens v o n E. „sanmeniensis" läßt v o n vornherein Uneinheitlichkeit vermuten; dies wird durch V I R E T (1954) bestätigt. N a c h diesem Autor besteht E. sanmeniensis a u s einer v e r
-') M. W. ist auch für die Art bzw. Unterart E. gmelini ANT. bisher auch noch kein Typen exemplar festgelegt worden.
hältnismäßig großen und einer sehr kleinen F o r m (im D i a g r a m m ist nach den Angaben V I R E T ' S das Feld auch für die M t c aufgegliedert, bei den M t t sind nur die M a ß e der großen F o r m berücksichtigt). N a c h den von ZDANSKY ermittelten Zahlen (n. GROMOVA) scheinen aber Ü b e r g ä n g e v o r z u k o m m e n . : 3® ibq K ° i + I • I Breite in mm 27 jl -»_ " d t i. Z . ~ -b : g 1. .;. »! _ _ J J I -D—l -Q-iO o o ' o öji 30 Breite i, Gruppe: Mosbach Körbisdorf Freyburg b Solutre 0 f- stenonis (St. Va liiert a E.'sanmeniensis" (Grosse form)
•
b E."sanmeniensis" (Kleine Form) 3 E.Przewalski 0 Caballus - Gruppe O+
Einzelexemplare von E. mosbachensis E.spec. Körbisdorf E.spec Frey bürg (Grosse Form) E.spec. Freyburg (KleineForm) E. gmeliniAbb. 2. Verschiedene Typen unter den Pferden der Caballus-Gruppe (einschl. „Allohippus"). a: Metacarpale, b: Metatarsal.
Pleistozäne Pferde der Caballus-Gruppe in Europa und Nordasien 1 0 7
D a s D i a g r a m m 2 erlaubt folgende Feststellung:
1) D i e untersuchten west- und mitteleuropäischen Formen gestatten eine Aufgliederung in 4 Größengruppen ( „ T y p e n " ) :
a) T y p u s M o s b a c h : Groß- und plumpwüchsig2) b) T y p u s K ö r b i s d o r f : Mittelgroß und p l u m p
c) T y p u s E. stenonis, St. V a l l i e r . "Wie Körbisdorf, nur etwas kleiner
d) T y p u s F r e y b u r g b : Klein u n d p l u m p . N B . F r e y b u r g a gehört z u m T y p u s K ö r b i s dorf.
2) Bei den T y p e n a, b und d verhalten sich die M t c und M t t in B e z u g auf ihre D i m e n sionierung gleichsinnig, das kleinere Pferd von S t . Vallier besitzt dagegen vergleichs weise längere M t c und kürzere M t t ; es vertritt daher einen grundsätzlich anderen Proportionierungstyp.
3) D i e 4 Gruppen bilden eine „allometrische R e i h e " , d. h. mit der G r ö ß e nimmt auch die relative Plumpheit zu. Merkwürdigerweise entspricht dem nicht die Verteilung der einzelnen E x e m p l a r e im zugehörigen Feld. D i e Anordnung zeigt im Gegenteil eher das Bild einer negativen Allometrie, d. h. die größeren E x e m p l a r e sind zugleich relativ schlanker. Bei den M t t (abgeschwächt auch bei den M t c ) des T y p u s Mosbach lassen sich z w e i U n t e r g r u p p e n erkennen, j e d e mit einer merklichen T e n d e n z z u einem negativ allometrischen Verhalten (Geologische Altersverschiedenheit? Sexuelle Unterschiede?). 4) Sämtliche 4 T y p e n gehören in die Reihe der „schweren" Pferde mit kurzen M e t a
podien.
5) Bei den geologisch ältesten T y p e n (Mosbach), abgeschwächt auch beim T y p u s St. V a l lier, schwankt die Länge verhältnismäßig stärker (Längeninstabilität), bei den jün geren T y p e n K ö r b i s d o r f und F r e y b u r g b besonders beim letztgenannten (Mtc!) d a gegen die Breite (Breiteninstabilität). Zieht man die große Zahl der E x e m p l a r e beim T y p u s Mosbach und T y p u s S t . V a l l i e r in Betracht, dann ist die Größenschwankungs breite, ausgedrückt durch die r e l a t i v e Feldgröße überhaupt bei jenen bedeutender als bei den älteren T y p e n .
6) D a s Przewalskipferd hat mittelgroße, aber zugleich schlanke Metapodien. M t c und M t t verhalten sich gleichsinnig. D a s durch das rezente Wildpferd besetzte Feld ordnet sich n i c h t der oben aufgestellten allometrischen Reihe ein, es ist kein plumpes Pferd. 7) D a s gleiche gilt v o n E. „sanmeniensis", wenigstens w a s die G r o ß f o r m angeht. Diese ist in ihrer M t c groß wüchsig und zugleich schlank, in ihren Mtt dagegen ist sie nur über mittelgroß und dabei mäßig p l u m p . M t c und M t t verhalten sich a l s o ebenso wie bei
E. stenonis v o n S t . Vallier n i c h t gleichsinnig. N o c h ausgeprägter als bei den beiden
europäischen Altpleistozäntypen ist die relative Längeninstabilität und die relative Breitenstabilität.
8) D a s eine T a r p a n e x e m p l a r n i m m t seinen P l a t z im Felde des T y p u s Freyburg b ein. 9 ) D a s Solutrematerial läßt sich nicht den vier ermittelten T y p e n zuordnen. N a c h den M t c gehört es z u m T y p u s F r e y b u r g b , die M t t sind aber im Verhältnis zu den M t c e t w a s größer u n d plumper, ihr F e l d überdeckt sich in seiner oberen Hälfte mit jenen des T y p u s K ö r b i s d o r f , in seiner unteren dagegen mit jenen des T y p u s Freyburg b. O b tatsächlich sämtliche M e t a p o d i e n nur e i n e r F o r m angehören, die einen T y p u s repräsentieren w ü r d e , wie er sonst bei den europäischen Pferden nicht auftritt ( U n -2) Ein in der Tabelle bei SCHWARZ ( 1 9 2 7 ) geführtes Mtt (Mosbacher Sande, Lok. „Hessler") m. einer größten Länge von 3 7 1 mm und einer Diaphysenbreite von 3 4 , 5 mm weicht dermaßen von den für E. mosbacbensis gewöhnlichen Dimensionen ab, daß es hier unberücksichtigt blieb. Wenn nicht überhaupt ein Druckfehler vorliegt, so ist dieses Stück entweder überhaupt nicht auf
E. mosbacbensis zu beziehen, was wahrscheinlicher ist, oder es stammt von einem sehr jugend
gleichsinnigkeit der Proportionen der M t c und Mtt, aber anders als bei E. stenonis von St. Vallier und E. sanmeniensis, große F o r m ) oder ob ein Formengemisch vorliegt, kann erst entschieden werden, wenn ausreichende, von einer H a n d unternommene Messungen durchgeführt werden.
IV
Die bisher getroffenen Feststellungen und Überlegungen gestatten einen Schritt weiter zu gehen und D i a g r a m m e aufzustellen, in welchen sämtliche mir zugänglich gewordenen Werte des europäischen und nordasiatischen nachmindelzeitlichen Materials eingetragen sind, soweit es sich auf Angehörige der Untergattung Caballus bezieht.
Aus den D i a g r a m m e n läßt sich nicht nur die Größenposition jedes einzelnen Restes erkennen, sondern es wurde auch versucht, seine chronologische und ökologische Stellung sichtbar zu machen. D a ß dabei nur sehr grob verfahren werden konnte, bedarf keiner be sonderen Begründung. Als chronologische K a t e g o r i e n erscheinen: Holstein, Riss, Eem, W ü r m und „Alter ungewiß, aber zweifelsfrei nachmindelzeitlich", die ökologische G r u p pierung erfolgte nach: w a r m (vollwarmzeitlich), kühl-kalt, vollkaltzeitlich und T e m peraturverhältnisse ungewiß (z. T . mit den Varianten „ u n g e k l ä r t " und „vermutlich mit Warmeinfluß" ( = gemäßigt, Interstadial, wärmere Phasen einer K a l t z e i t ) . Die Zahlenwerte wurden den Arbeiten von GROMOVA ( 1 9 4 9 ) , SCHWARZ ( 1 9 2 7 ) , W E R N E R T ( 1 9 5 7 ) , W E I L E R ( 1 9 3 7 ) , ARAMBOURG ( 1 9 5 8 ) , v. R E I C H E N A U (1915) u . a . entnommen, zum geringsten Teil beruhen sie auch auf eigenen Messungen (Steinheim/Murr, Wunstorf, Aschenstein).
a) W e s t - u n d M i t t e l e u r o p a ( D i a g r a m m e 3a u. b)
Ohne den Verhältnissen zu großen Z w a n g anzutun, kann eine Gruppierung des ge samten Materials nach den oben aufgestellten T y p e n Mosbach, K ö r b i s d o r f und Freyburg b vorgenommen werden. D e r altpleistozäne T y p u s St. Vallier, dessen Zugehörigkeit zur Untergattung Caballus unwahrscheinlich ist, bleibt in diesem Zusammenhang unbeachtet. SCHWARZ hat in seiner Arbeit ebenfalls unter Benutzung der Metapodien sein Material nach drei Größengruppen geordnet, die den T v o e n Mosbach, K ö r b i s d o r f und Freyburg b entsprechen (1927). Im G r u n d s a t z herrscht also zwischen seiner und unserer Auffassung Übereinstimmung. N u r ein allerdings sehr wesentlicher Unterschied besteht, während SCHWARZ in diesen drei G r u p p e n zugleich drei selbständige Unterarten erblickt, die von ihm als E. caballus robustus P O M . , E. caballus plicidens O W E N U. E. caballus caballus L . bezeichnet werden, wird hier eine derartig vereinfachende taxionomische Ausdeutung abgelehnt.
1. Der Größentypus Mosbach tritt in der H o l s t e i n - W a r m z e i t (Achenheim, Wunstorf) und in der E e m - W a r m z e i t auf (Weimar, Burgtonna, Gräfentonna, Phöben), der K l i macharakter der Fundschichten kann als ziemlich gesichert angesehen werden. Dagegen bleibt dieser bei den unten zu nennenden Fundlokalitäten zweifelhaft, da entweder die betreffenden Ablagerungen verschiedene, sich klimatisch nicht entsprechende Alters stufen umfassen (Fontechevade und Roter Berg) oder der Verdacht auf V o r k o m m e n auf sekundärer L a g e r s t ä t t e nicht von der H a n d zu weisen ist (Mittenwalde, R i x d o r f e r H o r i z o n t ! ) , oder die geologischen und d a m i t klimatischen Verhältnisse nicht k l a r ge nug sind (Vieselbach, H e i l i g e n s t a d t ) . Sicher ist aber, daß nirgends ein Pferd von der Größe des T y p u s Mosbach in Gesellschaft einer typischen K a l t f a u n a auftritt. Weiter läßt sich feststellen, daß in keinem Falle ein würmzeitliches Alter eines großen Pferdes sich beweisen läßt, wenn auch im Falle Heiligenstadt und Mittenwalde die Anwesen heit eines solchen im frühen W ü r m (Göttweiger Interstadial?) sich andererseits nicht g a n z ausschließen läßt. D i e Proportionsunterschiede bei der großen warmzeitlichen F o r m sind bedeutender als beim T y p u s Mosbach s. Str., d a neben einer sehr plumpen Form auch eine sehr große, schlanke auftritt (Burgtonna, s. auch S. 107).
Breite
Gruppe: Solutre Mosbach Pfeddersheim (grosse Form) Körbisdorf E.Przewalski Freyburg b Caballus-Gruppe Einzelexemplare aus • Holstein o R/ss © Riss-Eem » Eem O Würm Q Würm (Interstadial bzw eemzeitl. auf sekund. Lagerstätte ) _ Alteru. klimot. Verhältnisse
nicht geklärt + Subrezent (E.gmelini)
._| . 36Ü • 5 6 » r : . . B : . . . . o _ . I 15 ! I o o o 8 8 15 : . ? _ ? ' o
°
25 15 f->i 6 28 23! ID O: J 4 3 5 3 5 15 8 29 o 0*0 35 e ©2 I ; n 35„ O <.8o O 08 0<-3 O20 o1*3 ! °29 O S ^35 024 J C i S _ Breite in mm Gruppe •• Mosbach Körbisdorf Frey burg b • Solutre• PFeddersheim (grosse Form} • E.Przewalski • Caballus-Gruppe Einzelexemplare aus • Holstein 9 Piss © Riss - Eem 0> Eem O Würm Würm (Interstadial bzw. eemzeitl. auf sekund. Lagerstätte ) Alter u. hlimat. Verhältnisse nicht geklärt
Pleistozäne Pferde der Caballus-Gruppe in Europa und Nordasien 111
2. Die überwiegende Zahl der nachmindelzeitlichen Pferde zeigt Mittelgröße, die durch den T y p u s K ö r b i s d o r f bestimmte G r ö ß e n o r d n u n g wird aber z . T . nicht unwesentlich über-, v o r allem aber häufig unterschritten, w a s die Längendimension angeht, auch der Breitenspielraum erweist sich als bedeutender; für eine in sich geschlossene F o r m , etwa im R a n g e einer guten Unterart ist diese Variabilität entschieden zu groß, auch wenn die bedeutende zeitliche Streuung in Rechnung gestellt w i r d . D e r T y p u s K ö r b i s dorf stellt offenbar ein Formengemisch d a r , über dessen N a t u r noch zu sprechen sein wird.
Die Mehrzahl der Reste läßt sich ohne weiteres dem K ö r b i s d o r f e r T y p u s s. str. z u ordnen. E s sind jene der Fundlokalitäten Westeregeln z. T., Quedlingburg (große F o r m ) , Freyburg (gr. F . ) , sämtliche Lokalitäten des Rixdorfer H o r i z o n t e s mit A u s n a h m e Mittenwalde (Niederlehme, Rixdorf, K ö r b i s k r u g , Kreuzberg, Königswusterhausen), Görsdorf, B a r l e y , H a l b e , R o t e r Berg (kleine F . ) , Unkelstein ( T y p e von E. germanicus), Pfeddersheim (gr. F . ) , Fontechevade ( z . T . ) , Steeden ( z . T . ) , Oreston C a v e , V o g e l h e r d
(gr. F . ) , Pössneck, Lengefeld, Veltheim, Ofnet ( z . T . ) . D e m Alter nach gehört das M a t e rial entweder ins Riß oder Würm, vermutlich mit Ausnahme des einen Steinheimer E x e m p l a r e s . Bei keinem Stück ist eemzeitliches Alter sicher (Fontechevade?, R o t e r Berg?). F ü r einen größeren T e i l sind kältere K l i m a b e d i n g u n g e n w ä h r e n d der B i l d u n g s zeit der Fundschichten nachweisbar, die T e m p e r a t u r v e r h ä l t n i s s e einer V o l l w a r m z e i t umgekehrt höchstens für Fontechevade, vielleicht auch für Steinheim anzunehmen. Größer als es dem Zentralbereich des T y p u s entspricht, sind die E x e m p l a r e von Stein heim (mit einer A u s n a h m e ) , Achenheim, R o t e r Berg z . T . , Q u e d l i n b u r g (?). Sie k ö n n ten gegebenenfalls auch für kleinwüchsige Vertreter des Mosbacher T y p u s angesehen werden. E i n höheres als würmzeitliches A l t e r ist bei Steinheim und Achenheim sicher, bei „ R o t e r B e r g " möglich. D i e T e m p e r a t u r e n sind w a r m bis gemäßigt. Altersmäßig und ökologisch paßt das große Quedlinburger E x e m p l a r jedoch nicht in diese G r u p p e . Kleiner s i n d : Quedlinburg z . T . , Westeregeln z . T . , Steeden z . T . , Roter Berg z . T . , Vogelherd z. T . , Fontechevade z. T . , T h i e d e , O f n e t z. T , D a t t e l n , Aschenstein. K e i n e Fundstelle ist älter als das R i ß und nur Fontechevade und vielleicht auch R o t e r B e r g älter als W ü r m . Sieht m a n von den beiden letztgenannten F u n d p l ä t z e n ab, können kaltzeitliche K l i m a z u s t ä n d e vorausgesetzt w e r d e n .
Schlanker s i n d : Steeden z . T . , Solutre ( z . T . ? ) , Streckau, Lauchstedt, Fontechevade z . T . , Westeregeln (subfossil?). In den P r o p o r t i o n e n nähert sich diese S u b v a r i a n t e den Proportionen des Przewalski-Pferdes. Möglicherweise handelt es sich auch um
beson-Abb. 3 (vor- u. nebenstehend). Nachmindelzeitliche Pferde in Mittel- und Westeuropa, a: Meta c a r p a l , b: Metatarsale. Die Bezifferung kennzeichnet folgende Fundorte:
1 Achenheim 2 Barley 3 Burgtonna 4 Koburg 5 Datteln 6 Dörstewitz 7 Fontechevade 8 Freyburg u.d.U. 9 Görsdorf 10 Gräfentonna 11 Halbe 12 Heiligenstadt
13 Kalla 25 Oreston Cave 14 Königswusterhausen 26 Pfeddersheim 15 Körbisdorf 16 Körbiskrug 17 Kreuzberg 18 Lauchstedt 19 Lengefeld 20 Lingolsheim 21 Mainz-Kastell 22 Mittenwalde 23 Niederlehme 24 Ofnet 27 Phoeben 28 Poessneck 29 Quedlinburg 30 Rixdorf 31 Roter Berg 32 Schussenquelle 33 Solutre 34 Sossenheim 35 Steeden 36 Steinheim/Murr 37 38 39 40 41 42 43 44 45 46 47 Streckau Thiede Kosten b. Türmitz Unkelstein Veltheim Vieselbach Vogelherd Weimar Westeregeln Wunstorf E. gmelini Aschenstein
ders große Vertreter des T y p u s Freyburg b, jedoch ist dies nicht sehr wahrscheinlich. Sieht m a n wieder von Fontechevade ab, ist würmzeitliches Alter sicher oder v e r m u t b a r . Das G e s a m t b i l d erlaubt folgende Feststellungen:
Innerhalb des Formengemisches T y p u s K ö r b i s d o r f sind einerseits die geologisch ä l t e ren (Steinheim, Achenheim, K ö r b i s d o r f ) , andererseits die ökologisch „ w ä r m e g e t ö n ten" F o r m e n (z. B . R o t e r Berg z. T . , R i x d o r f e r Horizont) mit einiger Deutlichkeit durchschnittlich größer als die jüngeren und zugleich unter ungünstigeren K l i m a bedingungen lebenden (über die Pferdereste v o n Fontechevade s. S . 1 2 0 ) .
3. D a zwischen den einzelnen Varianten des T y p u s keine scharfen Grenzen bestehen, Übergänge also vorhanden sind, besonders zwischen dem Zentraltypus und seinen Kleinvarianten, bietet sich die Annahme, es hätte in den einzelnen unabhängig v e r laufenden Stammlinien eine fortschreitende Größenabnahme v o m Holstein bis z u m E n d w ü r m stattgefunden, als nicht unwahrscheinlich an, d a j a die geologisch jüngeren Vertreter des T y p u s im allgemeinen kleiner sind. Der Längenschwund war aber allem Anschein nach nicht von einer Verringerung der Breite im Sinne einer positiven A l l o -metrie begleitet, mit anderen Worten, die kleineren Pferde w a r e n verhältnismäßig plumper.
4. Zu dem durch das kleinere Freyburger Pferd (Freyburg b) gegebenen T y p u s zählen ausnahmslos E x e m p l a r e , deren würmzeitliches Alter entweder beweisbar ist oder w a h r scheinlich gemacht werden kann. Allerdings besteht bei dem einen oder anderen Stück gerade dieses T v p u s aus älteren Aufsammlungen die Möglichkeit, daß altholozänen Wild- oder g a r Hauspferdresten ein höheres Alter zugeschrieben wurde, da V e r m e n gung schon a m F u n d p l a t z (gestörte Höhlenablagerungen!) oder durch unvollkommene Grabungstechnik eingetreten ist. Der Verdacht auf interstadiales Alter und dement-sprechendem K l i m a besteht in keinem einzigen Falle. Vielfach läßt sich sogar eine Einstufung in den Zeitabschnitt Würm II u. I I I vornehmen. I m Falle des F u n d k o m plexes Vogelherd, Achenheim. Pfeddersheim, Quedlinburg und wie schon e r w ä h n t , Freyburg selbst, wird das kleinwüchsige Pferd des T y p u s F r e y b u r g b von einem g r ö ßeren P f e r d begleitet, das dem T y p u s K ö r b i s d o r f angehört u n d das offenbar z u m i n dest teilweise gleichzeitig mit ihm zusammen gelebt hat. Z u ähnlichen Feststellungen sind bereits andere Autoren gelangt, wobei sie sich nicht nur auf metapodiales M a t e r i a l beziehen ( z . B . LEHMANN 1 9 5 3 , LUNDHOLM 1 9 4 9 ) . Weitere V o r k o m m e n sind: L i n g o l s -heim, D ö r s t e w i t z , K o b u r g , K o s t e n b. T ü r m i t z , M a i n z - K a s t e l l , Sossen-heim, Schussen-quelle, Streckau. U n k l a r ist, wie bereits erwähnt, die Stellung des Solutrematerials, d a s nach den Ausmaßen der M t c eindeutig z u m T y p u s Freyburg b gehört, in B e z u g auf die Dimensionen der M t t sich dagegen stark dem T y p u s K ö r b i s d o r f nähert. Auch die Zugehörigkeit der K l e i n f o r m von Fontechevade, des Lauchstädter Pferdes und eines Teiles der Steedener und T h i e d e r Materialien muß offen bleiben. V o n Wichtigkeit ist aber, daß wenigstens nach d e m einen vermessenen E x e m p l a r zu schließen, der T a r p a n zum T y p u s Freyburg b zählt. Keine V e r b i n d u n g kann aber z u m P r z e w a l s k i - P f e r d hergestellt werden, auf dessen besondere Stellung im R a h m e n der gesamten U n t e r gattung bereits hingewiesen wurde. D a s Fehlen eines Pferdes v o n den E x t r e m i t ä t e n proportionen des E. Przewalski unter den pleistozänen E q u i d e n E u r o p a s k a n n d a h e r als erwiesen gelten, wenn m a n von dem durch zwei Mtt belegten Pferd des F u n d p l a t z e s K a l l a bei Biebrich absieht. D i e Stücke sollen aber oberflächennah (?) in einem S a n d l ö ß gefunden worden sein, der Mosbacher S a n d e überlagert. D i e Möglichkeit besteht also immerhin, daß es sich um F u n d g u t aus einer prähistorischen S t a t i o n handelt, w a s ich stark vermuten möchte. S i n d es aber tatsächlich Reste eines pleistozänen W i l d p f e r d e s , wäre dies als Hinweis aufzufassen, daß im W ü r m Mitteleuropas neben einem genü gend belegtem tarpanoiden Pferd auch eine dem heutigen mongolischen W i l d p f e r d entsprechende Form als Seltenheit v o r k a m .
Pleistozäne Pferde der Caballus-Gruppe in Europa und Nordasien 113
b) O s t e u r o p a u n d N o r d a s i e n ( D i a g r a m m e 4 a u. b)
Anzahl der für die Aufstellung der D i a g r a m m e herangezogenen Exemplare v o n fol genden L o k a l i t ä t e n :
Mtc: J a n a 17
M t t : U n g a 3, J a n a 16, S t a r y K o d a k 3, K u b i e k o w 5, K o t e l n y ?
Die A n z a h l der Stücke der anderen L o k a l i t ä t e n läßt sich unmittelbar aus den D i a grammen ablesen.
Die Mehrzahl der Funde ist, ähnlich wie in M i t t e l - und Westeuropa, geologisch jung ( W ü r m I I u. I I I ) . Sicher älter sind ein Teil der nordsibirischen Stücke, die aus Schichten stammen, die nach F a u n a und F l o r a nur in einem Vollinterglazial entstanden sein können (wahrscheinlich Eem, vielleicht auch Holstein z . T . ) . Ins Riß sind möglicherweise die Fundschichten der Pferde v o n M y s y (E. caballus misst P A V L . ) und Tungur (E. caballus
chosaricus G R O M . ) einzustufen. Weitere Einzelheiten können den Angaben des A n h a n g e s
entnommen werden; auch auf die Schwierigkeiten, die quartären Bildungen des osteuropä isch-nordasiatischen Raumes in richtige zeitliche Beziehungen zu jenen des übrigen E u r o p a s zu bringen, sei hier hingewiesen. Besonderer Betonung bedarf es nicht, daß eine zutref fende Beurteilung der klimatischen Verhältnisse, die während der einzelnen Phasen des Pleistozäns in den verschiedenen Teilräumen dieses riesigen Gebietes geherrscht haben, noch bedeutend schwieriger ist, als für die vergleichsweise winzige Fläche West- und Mittel europas. Die kontinentale G r ö ß e des Raumes, der in keinem Zeitabschnitt des Pleistozäns auch nur annähernd gleichgeartete Landschaften umschloß, ließe erwarten, daß in diesen eine erheblich größere A n z a h l verschieden proportionierter P f e r d e als im Westen gelebt hätten. U m so überraschender ist daher, daß sich die allerdings spärlichen Funde, die bis her gemacht wurden, leicht in die für die westlichen Gebiete aufgestellten Größenkate gorien einreihen lassen. Aber nicht nur diese Einreihung vollzieht sich unschwierig, allem Anschein w a r auch das stammesgeschichtliche Geschehen sehr ähnlich, in dem nämlich auch im Osten E u r o p a s und N o r d e n Asiens offenbar die geologisch älteren Formen die größe ren, die kleineren zugleich die jüngeren sind. Auch das Fehlen v o n großen Pferden in kalttemperierten Gebieten ist wahrscheinlich, wenn auch natürlich nicht schlüssig z u b e weisen. Im einzelnen ergibt sich folgendes B i l d :
1. Große Pferde vom T y p u s Mosbach sind nur aus dem hohen N o r d e n belegt: T a v d a und J a n a ( z . T . ) . D a aus Schichten ü b e r d e m Bodeneis ( = W ü r m ) bis jetzt nur kleine Pferde bekannt wurden ( T a i m i r - H a l b i n s e l , K o t e l n y ) , kann m i t einiger Sicherheit für die großen Formen vollwarmzeitliches Alter angenommen werden (Eem?, Holstein ? ) . 2. Die Mehrzahl der Pferde gehört auch hier d e m T y p u s K ö r b i s d o r f an. Dieser T y p u s wird als Formengemenge aufzufassen sein, das auch die als E. caballus missi P A V L . und E. caballus chosaricus G R O M . beschriebenen Pferde umfaßt. In der oberen Hälfte des Feldes liegen die Werte jener Pferde, für die ein höheres A l t e r als Würm I I u. I I I bzw. eine wärmegetönte F a z i e s ihrer U m g e b u n g anzunehmen ist: T a v d a , I l s k a j a , Tungur, Scholchowo, in der unteren Hälfte befinden sich die geologisch jüngeren F o r men bzw. solche, die unter kühlen bis kalten Temperaturbedingungen lebten: M y s y , Südural ( z . T . ) , Borchievo, Kostenki, N o v g o r o d - S e v e r s k i , K o c h - K o b a , A d j i - K o b a , Stary K o d a k . D a die Fundschichten von K u b i e k o w a eine Zeitspanne umfassen, die vom Holstein (?) bis ins S p ä t w ü r m reichen soll, überrascht es nicht, daß die Werte weit gestreut sind und ein übermittelgroßes P f e r d des T y p u s Körbisdorf und eines des T y p u s Freyburg b anzeigen.
3. Für die Kleinpferde des T y p u s Freyburg b ist ein spätwürmzeitliches Alter entweder erwiesen oder zu vermuten (Tschikoi, J a n a z . T . , A n a b a r , K u b i e k o w a z. T . , V i e r kholenskaja gora). D a s S ü d u r a l p f e r d lebte anscheinend unter nicht ungünstigen K l i m a -8 Eiszeit und Gegenwart
280 ZOO 1 I i I 1 1 I 1 I 1 1 ' 1 1 ' L
-
--a
, — . . . i -_,———
j °1
-
j i1
-I , . . J i ? 7 1 i 1 • ] a J I 1
1
• 1 0„
61
- o 1 r 1 7 oo . 7 1 o" I -1 i.S,PK ! I 18 L . _0 15 (DI
0 - A/1o
1 3 i I I I 1 I > Breite in m m ' ' 1 i i i i i i I I 1 1 1 1 1-b
r i — - • 1--
i i i r-i L . l ® * . . -- T i 1 0i 7 L.l_Q17__o_'.l
. . i L
°°°r-:°J°|-
S r ^r — f — -h~ i ? i i - I\
1 „ o . . o o o J j oJ i I . I I 30 Gruppe. Mosbach — Korbisdorf - — Freyburg b Stary Kodak Europa Breite in mm — o — o — Jana —ooo—ooo Unga — o — o — o — Kotelny — o-o — o Kubiekowa. Caballus-Gruppe N. Asien -- £ Pnewalski 10 i3 Einzelexemplare aus O Eem O Wurm Alter u. klimatische d) Verhältnisse nicht geklärt(Holstein ? - Wurm) 4- Subrezent (E.gmelini)
Abb. 4. Nachmindelzeitliche Pferde in Osteuropa und Nordasien, a: Metarcarpale, b: Metatarsale. Die Bezifferung bezeichnet folgende Fundorte:
1 AdjiKoba 2 Anabar 3 Borchevo II 4 Bugatsch 5 Ilskaia 6 KochKoba 7 Kodak (Stary Kodak) 8 Kostenki I 9 Kubiekova 10 Mysy 11 Novgorod-Severski 12 Scholokova 13 Taimir 14 Tavda 15 Tschikoi 16 Tungur
17 Ural (Südural:
Bura-novka, Griebnievaia, Kotchari II, Kliut-schewa, Orlovsky-Höhle, Ust-Katav II) 18 Vierkholenskaja gora 19 E. gmelini
Pleistozäne Pferde der Caballus-Gruppe in Europa und Nordasien 115
bedingungen, die nordsibirischen T i e r e (Taimir, A n a b a r , K u b i e k o w a , J a n a z. T . ) sind die kleinsten bis jetzt bekanntgewordenen pleistozänen (und rezenten) Angehörigen der Untergattung überhaupt. Ebenso wie die europäischen, zeigen auch die asiatischen Kleinpferde in ihren Proportionen den T a r p a n - und nicht den P r z e w a l s k i - T y p u s , auch in Asien ist dieser unter dem fossilen M a t e r i a l nicht vertreten. Z u ihm könnte allenfalls nur das eine E x e m p l a r von Vierkholenskaja gora ( S p ä t w ü r m ? , H o l o z ä n ? ) passen.
V
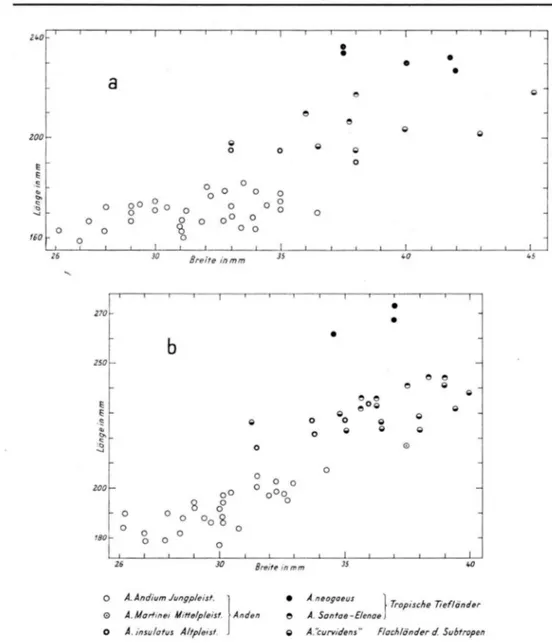
U m die G r u n d l a g e für die zu treffenden Folgerungen zu verbreitern, empfiehlt sich, zum Vergleich noch einen weiteren Formenkreis heranzuziehen. A m geeignetsten ist für diesen Zweck die Untergattung Amerbippus in ihren südamerikanischen Vertretern ( D i a g r a m m e 5a u. b ) . Ü b e r diese G r u p p e liegt eine neuere Beschreibung v o r , die sehr geeignet ist, die verfolgten Absichten zu unterstützen ( H O F S T E T T E R 1952). D i e südamerikanischen Arten von Amerbippus sind A. neogaeus L U N D , A. Santae-Elenae S P I L L M . , A. Martinei
S P I L L M . , A. insulatus C . A M E G H I N O , A. Andium W A G N E R - B R A N C O , A. curvidens O W E N ;
die letztgenannte ist allerdings nach H O F S T E T T E R als taxionomische Einheit „unrein", d a sie ein Formengemenge darstellt. W ä h r e n d die beiden erstgenannten Arten Bewohner v o n tropischen Flachländereien waren, lebten A. insulatus (Altpleistozän), A. Martinei (Mittel-pleistozän) und A. Andium ( J u n g p l e i s t o z ä n ) in den Hochanden. D e r Lebensraum v o n
A. „curvidens" schließlich waren die Ebenen, die sich im Süden an das Verbreitungsgebiet
von A. neogaeus anschlossen.
Aus einem Vergleich der betreffenden D i a g r a m m e ( 2 , 3, 4 u. 5) ergibt sich eine nahe zu vollständige Übereinstimmung, die fast verblüffend genannt werden kann. Dies w i r d nur unter der A n n a h m e verständlicher, die Entwicklung beider Equidengruppen sei von den gleichen Formungskräften gesteuert. Wie bei den nachmindelzeitlichen Formen von
Caballus lassen sich drei verschiedene Größen unterscheiden, w o b e i die durch die v o r
herrschende Mittelgröße gekennzeichnete G r u p p e auch hier ein Formengemenge ist und verschiedene Arten einschließt. D i e Zunahme der L ä n g e und Breite der Metapodien von Größenstufe zu Größenstufe folgt einer positiven Allometrie, aber auch bei Amerbippus sind Tendenzen z u verzeichnen, innerhalb einer A r t die positive in eine negative A l l o metrie umzukehren. Wie dies auch bei Caballus die Regel ist, besitzen die Mtc und M t t innerhalb jeder F o r m die gleichen oder sehr ähnlichen Proportionsverhältnisse. I m g a n z e n gesehen sind die jeweils arteigenen Proportionen bei den Mtt gefestigter als bei den M t c . Unter den andinen Formen ist wieder die geologisch jüngste, A. Andium, die kleinste. Eine Beziehung z u klimatischen F a k t o r e n scheint nur bei der G r o ß f o r m (A. neogaeus, tro pisches Flachland) und bei den Kleinformen (A. Andium, andines H o c h l a n d ) klar erkenn bar. Undeutlicher verhält sich in dieser Beziehung die Mittelgruppe, obwohl sich auch hier feststellen läßt, d a ß unter ihren Angehörigen A. Santae-Elenae als Bewohner tropischer Warmgebiete größenmäßig die S p i t z e hält. In B e z u g auf die P l u m p h e i t der M e t a p o d i e n ( L ä n g e : Breite) bestehen zwischen den größeren und kleineren A r t e n keine ersichtlichen Unterschiede (Allometrie!). Auch A. „curvidens" bildet keine A u s n a h m e , obwohl v o n den Angehörigen dieses Formgemenges noch am ehesten ein Leben in offeneren, ebenen L a n d schaften vorausgesetzt werden darf, wenn nicht wenigstens w ä h r e n d längerer Abschnitte des Pleistozäns auch diese Gebiete waldreicher als in der Gegenwart waren.
VI
Auf G r u n d der bisherigen Feststellungen k a n n der Versuch unternommen werden, das Bezugssystem Größenentwicklung und erdgeschichtliches Geschehen, deutend zu untersu chen. Auf teils ähnliche, teils auch stark abweichende G e d a n k e n g ä n g e in der bisherigen
L i t e r a t u r ( G R O M O V A 1949, L U N D H O L M 1949, H O F S T E T T E R 1952, L E H M A N N 1954, M O H R
Breite in mm A. Andium Jungpleist. A. Martin ei Mittelpleist. A. insulatus Altpleist Tropische Tiefländer • A. neogaeus in © A. Santae -Elenae I
Q A."curv/dens" Flachländer d. Subtropen
Abb. 5. Südamerikanische Arten der Gattung Amerbippus. a: Metacarpale, b: Metatarsale.
1958, D I E T R I C H 1950, S T E H L I N 1 9 3 3 , H E R R E 1938 usw.) sei v e r w i e s e n , eine ins einzelne
gehende Auseinandersetzung k a n n in diesem R a h m e n aber nicht erfolgen. Vorausgeschickt sei, daß das Phänomen der stammesgeschichtlichen Größenzunahme u n d -abnähme äußerst vielschichtig ist. Zahlreiche F a k t o r e n sind beteiligt: Ernährung, K l i m a , G e l ä n d e , F o r t bewegungsart, Weite und E n g e des Lebensraumes (Isolation!), Evolutionsgeschehen ( R a diation, Aussterben), B a s t a r d i e r u n g , Inzucht ( I s o l a t i o n ) , vielleicht sogar Großschwankun gen der kosmischen Strahlung usw. Bei einer Rechnung mit so vielen Unbekannten haftet jeder Deutung eines bestimmten stammesgeschichtlichen Geschehens Unsicherheit u n d U n -vollkommenheit an.
Innerhalb der Untergattung Caballus sind im jüngeren A l t q u a r t ä r E u r o p a s bisher nur große Pferde v o m T y p u s Mosbach bekannt geworden. K l e i n p f e r d e fehlen offensichtlich. Umgekehrt fehlen jene zumindest seit dem späteren Würm in E u r o p a und Asien. D e r
Pleistozäne Pferde der Caballus-Gruppe in Europa und Nordasien 117
Körbisdorf er T y p u s tritt in seiner Groß Variante (E. steinheimensis) zum ersten M a l zwi schen Mindel- und Rißkaltzeit in Erscheinung. Ihm gehören die Mehrzahl der riß- und frühwürmzeitlichen Pferde sowohl in E u r o p a als auch in N o r d a s i e n an. K l e i n p f e r d e vom T y p u s F r e y b u r g b fehlen bisher mit geologisch gesicherten Belegen, die älter als das spätere Würm sind. Zunächst scheinen diese Tatsachen dafür zu sprechen, daß in allen Stammes linien, die m a n sich vielleicht als engbenachbart vorzustellen hätte, eine gleichmäßige Größenabnahme stattgefunden hätte, wofür v o r allem die nicht z u übersehende, gewisser maßen stetige Größenabnahme innerhalb des T y p u s K ö r b i s d o r f v o m Beginn des Riß an bis zum End würm bzw. A l t h o l o z ä n spricht ( v g l . auch Amerbippus). Dieser Größenschwund kann aber, besteht diese A n n a h m e zu recht, nur von einem T e i l der S t ä m m e mitgemacht worden sein, denn Großpferde v o m Mosbacher T y p u s fehlen in E u r o p a u n d offenbar selbst in N o r d a s i e n noch im E e m , ja vielleicht sogar noch im frühen Würm, zumindest in den südlidten Gebieten, nicht. Ebenso vergesellschaftet sich der Körbisdorfer T y p u s mit dem Freyburger b im W ü r m und A l t h o l o z ä n . Ein Teil der durch Mittelgröße gekenn zeichneten Populationen unterlag also ebenfalls nicht der V e r z w e r g u n g . Es ist mithin auch die Annahme, bereits im P r ä m i n d e l hätte es Vertreter aller drei Größenkategorien, nur nicht die gleichen Regionen bewohnend, gegeben, vorläufig nicht widerlegbar, ja nicht einmal g a n z unwahrscheinlich. Sie wären d a n n nur etappenweise ausgestorben, wobei die Großpferde die ersten Opfer waren. D a s wirkliche Geschehen w i r d sich aber in der Form eines „sowohl — als auch" vollzogen haben, indem neben großen auch kleinere Pferde, aber wohl nicht Zwerge, schon frühzeitig vorhanden waren, eine echte Größenabnahme wenigstens in bestimmten Zweigen aber zweifellos stattgefunden hat. In diesem Z u s a m menhang sei d a r a n erinnert, d a ß die Pferde g a n z allgemein leicht und rasch zur Verzwer gung neigen ( „ P o n y s " , Inselpferde, vgl. L U N D H O L M 1 9 4 8 ) . W e n n die verschiedene Größe durch innere Faktoren allein bestimmt w ä r e , würden alle Überlegungen, inwieweit U m weltverhältnisse steuernd oder sogar verursachend eingegriffen hätten, müßig sein. Groß pferde treten aber in Mittel- und Westeuropa sowie in N o r d a s i e n allem Anschein nach nur in den wärmeren Klimaphasen auf (Cromer, Holstein, E e m ) , Kleinstpferde scheinen in ihrem Auftreten an ausgesprochene K a l t z e i t e n gebunden z u sein. Die kleinsten, bisher überhaupt bekanntgewordenen Pferde lebten in Nordsibirien an der Wende Pleistozän-H o l o z ä n . D e m könnte entgegengehalten werden, daß im A l t q u a r t ä r sehr verschieden große Pferde lebten, wie vermutet w i r d , wenigstens z u m Teil zu gleicher Zeit ( V I R E T 1 9 5 4 ) . Zu wenig ist aber bisher über d a s klimatische Geschehen, besonders im älteren A l t q u a r t ä r be kannt, zu unsicher ist dessen Chronologie und zu unklar die taxionomische Stellung der allem Anschein nach recht zahlreichen Formen (Caballus div. spec. ?, Allohippus d i v . s p e c ,
Asinus s p e c ? ) um die für die Caballus-Gruppe gemachten Feststellungen z u entwerten.
Auf Grund allgemeiner E r w ä g u n g e n darf überdies für die altquartären Pferde insgesamt ein ökologisch größerer S p i e l r a u m (Eurythermie!) anzunehmen sein. Erst im späteren Pleistozän werden sie, wenigstens teilweise, stenotherm g e w o r d e n sein. Die mittelgroßen Formen dagegen scheinen, wie auch nicht anders zu erwarten, in ihren ökologischen A b hängigkeiten unbestimmter. Bestehen solche Beziehungen zur T e m p e r a t u r tatsächlich, so folgt d a r a u s , daß für die P f e r d e entgegen der Ansicht GROMOVA'S ( 1 9 4 9 ) nicht die B E R G -MANN'sche R e g e l gilt. Auch bei der U n t e r g a t t u n g Amerbippus bewohnten die größeren Arten w ä r m e r e Gebiete. Bei anderen Pflanzenfressern können dafür gleichfalls Beispiele gefunden werden (Unterarten und Rassen der Gattung Rangifer, Elefanten der Gattung
Mammonteus). A u f der anderen Seite darf nicht übersehen werden, daß jene P f e r d e der
Gegenwart, die Bewohner heißtemperierter Regionen sind, s a m t und sonders, verglichen mit den F o r m e n der Untergattung Caballus kleiner sind, die weiter nördlich lebenden aber etwas größer. Diese Verbreitung der rezenten und pleistozänen Equiden nach ihrer Größe legt den Gedanken nahe, daß gemäßigte Klimabedingungen eine bedeutende K ö r pergröße begünstigen, mit anderen Worten, die Equiden folgen in der geographischen
An-Ordnung zunächst der BERGMANN'schen Regel; von einem gewissen kritischen Punkte an
schlägt die Größenentwicklung aber in ihr Gegenteil um, d a dann offenbar in den betref
fenden Populationen kleinere T i e r e durch Auslese b e v o r z u g t werden ( N a h r u n g , N a h
rungsaufnahme, U n t e r g r u n d ? ) . G r ö ß e n a b n a h m e ist ebenfalls, wie ihr Gegenteil, ein ent wicklungsgeschichtlicher V o r g a n g . Sic in jedem Fall nur als degenerativen Prozeß a n z u sehen, ist unzulässig. D i e Kleinpferde des Tungpleistozäns etwa nur als arktische K ü m m e r formen anzusprechen, ist daher a b w e g i g . Ist die einmal erreichte G r ö ß e aber durch A b n a h m e der V a r i a b i l i t ä t und einen d a m i t verbundenen Plastizitätsschwund (Stenothermie!) nach Abklingen der R a d i a t i o n s p h a s e erblich fixiert w o r d e n , dann können die betreffenden F o r m e n eintretenden T e m p e r a t u r ä n d e r u n g e n nur mit Arealverschiebungen begegnen; ist dies aus äußeren oder inneren G r ü n d e n nicht möglich, so wird das Aussterben unvermeid lich (Großpferde im L a u f e des W ü r m , Kleinpferde in der Nacheiszeit, T a r p a n ! ) , wobei die Großformen allem Anschein nach zuerst gefährdet sind. Die Aussichten für das Ü b e r leben würden für einen derartigen S t a m m nur dort gegeben sein, w o eine Änderung der U m w e l t nicht oder nur in geringerem M a ß e stattfand. N u r in solchen R ä u m e n konnten sich die Equiden bis in die G e g e n w a r t erhalten, w ä h r e n d sie an der klimatischen K a m p f front ausstarben. D a s Schicksal auch anderer S ä u g e r g r u p p e n im pleistozänen Eurasien dürfte sich ähnlich gestaltet haben, a m vollkommensten scheint die Entsprechung bei den Proboscidiern zu sein: H ö h e p u n k t der Größenentwicklung bei Archidiskodon,
Palaeolo-xodon und Mammonteus im jüngeren Altquartär, H e r a u s b i l d u n g kleinerer Rassen bei M. trogontherii b z w . M. primigenius im N o r d e n , Persistieren der G r o ß f o r m e n als K o n
servativstämme im Süden (P. antiquus, M. trogontherii) und Ausweitung ihres Areals in den klimatisch günstigeren Phasen, Aussterben im L a u f e des Würms auch in Südeuropa, Weiterleben der nördlichen kleinen Primigeniusrassen bis ins beginnende Postglazial, E r leben der G e g e n w a r t nur in den klimatisch stabilen R ä u m e n Indiens und Afrikas
(Euele-phas, Loxodonta).
Ein anderes Problem stellt sich mit der Frage der Bedingtheit der Proportionsverhält nisse, also von Schlankheit und P l u m p h e i t der M e t a p o d i e n bei den verschiedenen Formen („leichte" und „schwere" Pferde). D e r G r a d der Plumpheit wird durch das Verhältnis L ä n g e : Breite bestimmt („Schlankheitsindex"). N a c h den Untersuchungen H O F S T E T T E R ' S ( 1 9 5 2 ) sind außerdem bei den „schweren" Equiden die Metapodien, verglichen mit den anderen Gliedmaßenabschnitten v e r k ü r z t , mit anderen Worten, die Plumpheit wird v o r allem durch die relative K ü r z e v o n Mittelhand und -fuß hervorgerufen. W i e aber H O F S T E T T E R betont, greift die V e r k ü r z u n g (bei etwa gleichbleibender S t ä r k e ) auch auf die proximalen Gliedmaßenteile über. In der Literatur w i r d überwiegend die Auffassung ver treten, in den Großpferden des Pleistozäns seien W a l d - , in den mittelgroßen und beson ders den kleinen hingegen T u n d r a b e w o h n e r zu sehen. Sie stützt sich dabei auf die B i n d u n g an warmzeitliche Phasen auf der einen, an kaltzeitliche auf der anderen Seite. D i e W a l d p f e r d e sollen dabei besonders „schwer", d. h. p l u m p sein. Bei den pleistozänen Pfer den der Caballus-Gruppe — ob groß, ob klein — gibt es im wesentlichen aber keine, die schlankwüchsiger oder plumper sind als andere. Sie w a r e n ausnahmslos schwer und hatten verkürzte Metapodien. M a n könnte höchstens die d a und dort erkennbaren Ansätze zu einer negativen Allometrie innerhalb eines T y p u s b z w . einer P o p u l a t i o n als Ausdruck d a f ü r werten, daß der V e r k ü r z u n g s v o r g a n g als solcher nicht abgeschlossen war.
Extremitäten mit verkürzten M e t a p o d i e n besitzen so gut wie sämtliche Pleistozän-formen, ausgeprägter die Angehörigen der Caballus-Gruppe, undeutlicher und a b g e schwächt die große F o r m des E. sanmeniensis und vermutlich auch die Großformen des
Allohippus-'K.relses. Schlankwüchsig hingegen sind die Esel, Halbesel und Zebras, aber
auch das Przewalski-Pferd ist verhältnismäßig schlank. Plump sind hingegen wieder die südamerikanischen Vertreter v o n Amerbippus. G a n z allgemein ist die Schlankheit der Extremitäten ein Kennzeichen leichter, rascher und nachhaltiger Beweglichkeit, während