Vol. 21 1, No. 3 Prthtedm U.S.A.
596
0022-3565/79/21 I3-0596$02.0O/O
Tsv JOURNAL OY PHARMACOLOGY AND EXPERIMENTAL THERAPEUTiCS
Copyright © 1979 by The American Society for Pharmacology and Experimental Therapeutics
Discriminative
Stimulus
Effects
of Naltrexone
in the
Morphine-Dependent
Rat1’2
VANCE F. GELLERT and STEPHEN G. HOLTZMAN
Department of Pharmacology, Emory University School of Medicine, Atlanta, Georgia
Accepted for publication August 27, 1979
ABSTRACT
Gellert, Vance F. and Stephen G. Holtzman: Discriminative
stimulus effects of naltrexone in the morphine-dependent rat.
J. Pharmacol. Exp. Ther. 21 1 : 596-605, 1979.
Rats maintained physically dependent upon morphine by
scheduled access to drinking water containing morphine were
trained to discriminate between s.c. injections of saline and 0.1
mg/kg of naltrexone in a discrete trial avoidance procedure in
which a response on one of two choice levers would prevent or
terminate the delivery of mild electric shocks to the floor of the
test chember. Stimulus control of behavior by naltrexone in the
morphine-dependent rat (defined as the reliable completion of
at least 1 8 trials of a 20-trial session on the appropriate choice
lever) had many of the features previously described for the
stimulus control of behavior by morphine in the nondependent
rat: long-term stability and reproducibility, orderly dose- and
time-effect relationships and pharmacologic specificity.
Stim-ulus control by naltrexone was blocked in a dose-related
man-ner by morphine, an effect completely surmounted by a 1 0-fold
increase in the dose of naltrexone suggesting a competitive
antagonism . The naltrexone-induced discriminative stimuli
ap-peared to be related to precipitated morphine withdrawal
phe-nomena: following the abrupt withdrawal of morphine the
amount and time course of naltrexone-appropriate responding
were directly related to the degree of physical dependence;
loss of body weight, a reliable index of morphine withdrawal in
the rat, paralleled changes in naltrexone-appropriate
respond-ing; the maximum level of naltrexone-appropriate responding
produced by a total of eight narcotic antagonists with agonist
activity of differing prominence was a function of the extent of
separation of the agonist and antagonist components of action
of the drugs. Control of behavior by stimuli associated with
morphine withdrawal may afford a specific animal model for
studying factors relevant to the perpetuation of chronic drug
use by human addicts.
Morphine and related narcotic analgesics produce a
distinc-tive syndrome of alterations in thought, mood and perception
such as feelings of relaxation, “coasting,” and a sense of
well-being (i.e., “euphoria”) that is commonly described as pleasant
by individuals experienced in the illicit use of narcotics and
other drugs (Martin and Fraser, 1961; Haertzen, 1974). The
nature of these subjective effects appears to be a principal
factor in the high abuse potential of the narcotic analgesics
(Fraser, 1968; Jasinski, 1973, 1977). The results ofrecent studies
have emphasized the parallelism of the stimulus effects of
morphine and other narcotic analgesics in the rat and squirrel
monkey determined in drug discrimination procedures and the
subjective effects of these drugs in man, and have led to the
suggestion that drug discrimination procedures may afford an
animal model for studying the subjective effects of narcotic
analgesics (Hirschhorn and Rosecrans, 1976; Shannon and
Holtzman, 1976a, 1977; Colpaert, 1977; Lal et at., 1977; Schaefer
and Holtzman, 1977).
Received for publication April 1 1, 1979.
This work was supported in part by U.S. Public Health Service Grant DA00541 from the National Institute on Drug Abuse and Research Scientist
Development Award K02 DA00008 to S.G.H.
2A preliminary report of this work appears in Fed. Proc. (37: 660, 1978).
Prominent subjective changes occur not only when morphine
or other narcotic analgesics are administered acutely, but also
when administration is abruptly terminated following a period
of chronic exposure to the drug. The narcotic withdrawal
syn-drome is a distinctly unpleasant event consisting of symptoms
such as weakness, anxiety, lack of motivation, irritability,
nau-sea, mental depression and dysphoria (Haertzen and Hooks,
1969; Jaffe, 1975). Thus, for the individual who is physically
dependent upon a narcotic analgesic, the desire to prevent the
emergence of withdrawal symptomatology becomes another
important factor contributing to the perpetuation of drug use
(Haertzen and Hooks, 1969; Jaffe, 1975; O’Brien, 1975).
The present study represents an initial effort to develop an
animal model for studying the discriminative stimuli associated
with the narcotic withdrawal syndrome. Rats that were
main-tamed physically dependent upon morphine by the method of
scheduled access to a morphine drinking solution (Gellert and
Holtzman, 1978) were trained to discriminate between
injec-tions of saline and the narcotic antagonist naltrexone (0.1 mg/
kg)
in a two-choice discrete-trial avoidance paradigm (Shannonand Holtzman, 1976a, 1977). A narcotic antagonist precipitates
a withdrawal syndrome in a physically dependent subject that
is qualitatively similar to the syndrome that develops when the
drug of dependence is abruptly withdrawn (Winkler et at., 1953;
at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
Stimulus Eftscts of Naltrexone
597
Martin, 1967; Jaffe and Martin, 1975), but the
antagonist-pre-cipitated withdrawal syndrome has a rapid onset and short
duration, features which are essential for a study of this type.
Naltrexone, like its structural analog naloxone, has little
de-tectable agonist activity (Martin et at., 1973; Blumberg and
Dayton, 1973, 1974), and was selected for use in this study
because of the advantage afforded by the longer duration of its
action compared to that of naloxone (Villarreal and Karbowski,
1974; Dykstra et al., 1974; Holtzman, 1976).
The principal objectives of this study were 1) to demonstrate
the feasibility of establishing stable stimulus control of choice
responding in the rat with stimuli associated with an
antagonist-precipitated morphine withdrawal syndrome, and 2) to begin to
characterize those discriminative stimuli by determining the
extent to which they are generalized to a) the morphine
with-drawal syndrome induced by the abrupt termination of
mor-phine administration, b) other narcotic antagonists having
ago-nist activity of differing prominence and c) discriminable
non-opioid compounds that can produce some of the signs or
symp-toms of morphine withdrawal. The ability of morphine
injec-tions to block the stimulus control of behavior by naltrexone
was also assessed. Loss of body weight is perhaps the most
reliable single index of the morphine withdrawal syndrome in
the rat (Akera and Brody, 1968; Wei and Way, 1975; Gellert
and Holtzman, 1978). Therefore, body weight was monitored
during all of the experiments in order to determine the extent
to which naltrexone-induced weight loss and stimulus control
of behavior would be correlated.
Methods
Subjects. Male, Sprague-Dawley descended CFE rats Weighrng 120
to 140 g were obtained from Charles River Breeding Laboratories
(Wilmington, MA). Upon arrival from the supplier, the rats were
immediately placed into individual cages which were maintained in a
ventilated cabinet designed to permit control over the access of each
animal to its drinking solution (Gellert and Holtzman, 1978). Food was
always present in the home cage. A diurnal lighting cycle was
main-tamed by illuminating the cabinet between 7:00 A.M. and 7:00 P.M.
Morphine dependence. The method of establishing and
maintain-ing tolerance to and physical dependence upon morphine in the rat by
scheduled access to a morphine drinking solution has been described in
detail elsewhere (Gellert and Holtzman, 1978). This method involves
placing morphine in the rats’ only source of drinking water. Water
bottles are held in a bottle rack above the cages and can be rotated into and out of the animals’ home cage by activating a small motor located on the top of the cabinet in which the cages are kept. The concentration
of the morphine solution was gradually increased from 0.01% on the
first day to 0.05% (base concentration) on the 11th day, and was
maintained at this level for the duration of the study. The rats were
given access to the morphine solution for 10 mm four times daily at
5:00 AM., 11:00 AM., 5:00 P.M. and 11:00 P.M. Under this type of
scheduled access to a morphine drinking solution, rats have been shown
to consume an average of about 50 mg/kg/day of morphine with 20 to
30% of the daily dose being ingested at each of the access periods
(Gellert and Holtzman, 1978). Daily drug intake was not routinely
monitored in the present study. Our previous experiments have shown
that the development of morphine tolerance (as determined by the tail-flick test for analgesia) and physical dependence (as determined by the
intensity of the naloxone-precipitated withdrawal syndrome) reaches a
pleateau after about 3 weeks ofmorphine intake (Gellert and Holtzman, 1978).
Apparatus. A one-lever rat chamber (model 1 1 10-L, Grason-Stadler
Co., Bolton, MA) was modified by mounting two “choice response”
levers 15 cm apart on the wall opposite the original lever which was
designated the “observing response” lever. A clear Plexiglas partition
was mounted on the wall of the chamber between the choice response
levers. The partition protruded 5 cm into the chamber and extended
from the chamber ceiling to 1.0 cm above the grid floor. A scrambled
electric shock could be delivered to the grid floor of the chamber by a
constant current shock generator (model 700, Grason-Stadler Co.). The
test chamber was housed in a ventilated enclosure that was light-proof and sound-attenuating. Schedule contingencies were controlled by stan-dard relay programming equipment.
Discrimination training. After they had been maintained on the
regimen of scheduled access to the morphine solution for at least 1
month, the rats were trained to discriminate between saline and 0.1
mg/kg of naltrexone in a two-choice discrete-trial avoidance paradigm
(Shannon and Holtzman, 1976a, 1977). This dose of naltrexone and the
injection time of 15 min before the session were selected on the basis of
pilot experiments. The rats were trained to complete a two-response
chain in order to terminate a trial and avoid or escape from electric
shocks delivered to the floor of the test chamber. The beginning of a
trial was signalled by the simultaneous illumination of the house light
and the presentation of white noise. Starting 5.0 sec later, the chamber
floor was electrified (1.0 mA) for a 1.0-sec period every 3.0 sec. The
animal was required first to press the observing response lever, then to
press one of the two choice response levers on the opposite side of the
chamber. The first observing response of the trial terminated the white
noise; the appropriate choice response extinguished the house light and
ended the trial. A response on the inappropriate choice lever had no
programmed consequence. A trial was defined as correct if the rat
emitted the response sequence of observing lever-appropriate choice
lever and as incorrect if the rat emitted the response sequence of
observing lever-inappropriate choice lever-appropriate choice lever.
The interval between trials was 50 sec during which the test chamber
was dimly illuminated with red light. A session ended after 21 trials or
30 mm, whichever came first. The first trial of each session was
considered to be a “warm-up” trial and was excluded from the data
analysis.
Daily training sessions were conducted 5 days per week. All sessions began 15 min after the s.c. injection ofsaline or 0.1 mg/kg of naltrexone
which were administered in a sequence of double alternation (i.e.,
saline, saline, naltrexone, naltrexone, saline ...). Half of the rats were trained to press the right choice lever throughout each saline session and the left choice lever throughout each naltrexone session; the other
half of the rats were trained under the opposite conditions. Training
continued until a rat completed four consecutive sessions in which 19
out of 20 trials (after the first trial) were correct. When this criterion was met, the next four sessions (two saline and two naltrexone sessions
in random order) were conducted as test sessions in which both choice
levers were electrically activated so that following the observing
re-sponse a response on either choice lever terminated the trial regardless
of the pretreatment (nondifferential reinforcement of choice respond-ing). The behavior of the rat was considered to be under the stimulus control of saline and naltrexone if the rat completed at least 18 out of 20 trials on the choice lever appropriate for the substance administered
in each of the four test sessions.
Stimulus generalization tests. After stimulus control of behavior was established, training sessions with only the appropriate choice lever
activated continued to be held on Mondays, Wednesdays and
Thurs-days. Saline or 0.1 mg/kg of naltrexone was administered before each
training session according to a double alternation sequence. Tests of
generalization to novel drug conditions were conducted on Tuesdays
and Fridays under nondifferential reinforcement of choice responding
provided the animal had completed at least 18 out of 20 trials on the
appropriate choice lever in the four preceding training sessions. Each
rat was tested first with graded doses of naltrexone. Other drugs were then tested in a nonsystematic order. Within each series of drug tests,
doses were administered in a random sequence that also included saline
and the training dose of naltrexone. All drugs were injected s.c. 15 min before the start of a test session unless otherwise specified. The body
weight of each rat was measured just before administration of the test
at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
4 C,) U) 0 -J I Ui at 12
598
Gellert and Holtzman Vol. 211week of training.
The degree of physical dependence of the rats appeared to
drug and again at the conclusion of the test session. Training and test sessions took place from 12 noon to 4:00 P.M., 1 to 5 hr after a period of access to the morphine drinking solution.
Abrupt morphine withdrawal. Rats with stable discrimination
behavior were abruptly withdrawn from morphine by replacing their
0.05% morphine drinking solution with a 0.01% solution of quinine
sulfate. Quinine, like morphine, has a bitter taste and was used to
minimize the change in taste cues resulting from the removal of
morphine from the drinking water. Test sessions were conducted 15
min after an injection of saline at 0, 6, 12, 18, 24 and 36 hr after the last access to the morphine solution.
In order to induce a greater degree of morphine dependence than
could be achieved solely by the oral intake of morphine, other rats were
implanted subcutaneously with two pellets formulated according to the
method of Gibson and Tingstad (1970), each containing 50 mg of
morphine base. Two additional pellets were implanted 24 hr later.
Three days after the second two pellets were implanted, all of the
pellets were removed while the rats were lightly anesthetized with
ether. The morphine drinking solution was replaced with a 0.01%
quinine solution at this time. Test sessions were conducted 15 mm after
an injection of saline just before the pellets were removed (0 hr) and at 12, 18, 24, 36 and 60 hr after removal of the pellets.
Data analysis. The data for discriminative responding are presented
as the mean number of trials completed on the naltrexone-appropriate choice lever in a 20-trial session. The remaining trials of the session
were always completed on the saline-appropriate choice lever unless
indicated otherwise. Based upon probabilities derived from the
binom-ial theorem (Hays, 1973), as described previously (Shannon and
Holtz-man, 1976a). A test condition was considered to produce stimulus
control ofbehavior comparable to that ofthe training dose of naltrexone if the group completed an average of at least 18 out of 20 trials on the
naltrexone-appropriate choice lever.
Changes in body weight were determined for the interval between
the injection of saline or a test drug and the end of a test session, and
are presented as mean ± S.E.M. percent weight loss. The Spearman
rank correlation coefficient (r,) corrected for tied observations (Siegel,
1956) was computed for percent weight loss and number of trials
completed on the naltrexone-appropriate choice lever. Weight loss and
naltrexone-appropriate responding were considered to be significantly correlated if the P value of r, was less than .05.
Drugs. d-Amphetamine sulfate, morphine base, morphine sulfate,
sodium pentobarbital and physostigmine sulfate were purchased from
commercial sources. Diprenorphine hydrochloride, naloxone
hydro-chloride and naltrexone hydrochloride were obtained from the National
Institute on Drug Abuse. The following drugs were generously donated:
levallorphan tartrate (Roche Laboratories, Division of Hoffmann-La
Roche, Inc., Nutley, NJ), cyclazocine base and
pentazocine
base(Ster-ling-Winthrop Research Institute, Rensselaer, NY), nalorphine
hydro-chloride (Merck and Company, Inc., Chemical Division, Rahway, NJ),
and oxiorphan tartrate (Bristol Laboratories, Division ofBristol-Myers
Co., Syracuse, NY). Oxiorphan was dissolved in distilled water, and
cyclazocine and pentazocine were dissolved in a vehicle of 8.5% lactic
acid and 1.0 N sodium hydroxide in a 3:2 ratio; all other drugs were
dissolved in 0.9% saline. Drug solutions and saline were injected s.c. in
a volume of 1.0 mi/kg b.w. Drug doses are expressed in terms of the
free base or acid.
Results
Stimulus control of behavior by naltrexone.
Twenty-one rats were rendered physically dependent on morphine for
use in this study. Of those, 15 met the criteria for stimulus
control of behavior after 7 to 14 weeks of training (35-70
sessions), with most of the animals falling within the upper half
of this range. The other six rats were discarded after the 15th
remain constant during the period of approximately 9 months
that drug tests were conducted, as reflected by the consistency
of the weight loss data. Figure 1 shows the average percent loss
of body weight experienced by rats from the time of injection
of saline or 0.1 mg/kg of naltrexone to the end of the test
sessions which were inserted randomly among the various test
drug series described in this report. The interval of time over
which the measurements were made averaged 38 min (range:
35-45 mm). The mean loss of body weight ranged from 1.0 ±
0.1 to 1.4 ± 0.3% after an injection of saline, and from 4.0 ± 0.3
to
5.2
± 0.4% after an injection of 0.1 mg/kg of naltrexone. Each1.0% represents approximately 4.0 g.
Tests of stimulus generalization to graded doses of naltrexone
(0.003-0.1 mg/kg) were conducted twice in nine of the rats:
once immediately following the establishment of stimulus
con-trol of behavior by saline and naltrexone (0.1 mg/kg), and again
5 to 9 months later upon the completion of most of the
experi-ments described in this report. In both instances, 0.003 mg/kg
produced responding appropriate for saline, whereas higher
doses resulted in a progressive increase in the number of trials
completed on the naltrexone-appropriate choice lever (fig. 2).
Naltrexone (0.003-0.1 mg/kg) also produced a dose-related
increase in weight loss (fig. 2) which, for each of the two
determinations, correlated significantly with drug-appropriate
choice responding (curve 1: r5 = 0.57, P < .001; curve 2: r5 =
0.68, P < .001). That the first and second determinations of the
dose-response
curves
yielded almost identical results for thetwo pairs of curves is further indication of the long-term
stabil-ity of the preparation.
Abrupt morphine withdrawal. In order to determine how
closely the discriminative stimuli associated with
antagonist-precipitated withdrawal resemble those associated with
spon-taneous withdrawal, test sessions were conducted at
various
intervals after quinine was substituted for morphine in the
drinking solution of the rats. Figure 3 shows that mean
naltrex-one-appropriate choice responding increased gradually
fol-lowing withdrawal from morphine, reaching a peak of 7.2
re-spouses at 24 hr (based upon individual responses of 0, 0, 6, 13
5/\+#ThA
3 . NALTREXONE 0 SALINE 2 0’ I I I I I I I I 4 5 6 7 8 9 10 IlMONTHS OF MORPHINE ADMINISTRATION
Fig. I. Percent loss of body weight during monthly test sessions with
saline and naltrexone (0.1 mg/kg) in morphine-dependent rats trained
to discriminate between saline and 0.1 mg/kg of naltrexone. Change
in body weight was determined over an interval that averaged 38 mm.
Each point is a mean ± 1 S.E.M. based upon one observation in each of nine rats; the S.E.M. is not shown where it is less than the radius of
the point. The rats were stabilized on the morphine maintenance
regimen during month 1 of the study and were trained in the
discrimi-nation procedure during the following 2 to 3 months. Data are not
available for saline test sessions during month 12.
at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
20 -Ui > Ui -I Ui z 0 x Ui a: z U) -I a: I-U) U) 0 -J I-I Ui #{149}0 8 6 4 2 0 8 6 4’ 2 o 5 4 3 2 0 I I I
leled the changes in naltrexone-appropriate responding (fig. 3).
The total loss of body weight after 36 hr ofwithdrawal averaged
10.3 ± 1.7% for the rats that had been implanted with morphine
pellets and 7.2 ± 1.3% for the group that only had access to the
morphine drinking solution.
Stimulus generalization tests with narcotic
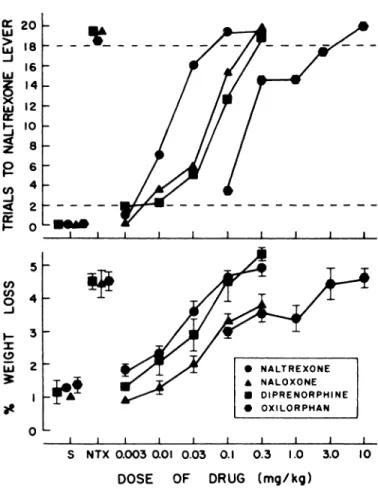
antago-fists. Seven drugs having narcotic antagonist activity in
addi-tion to naltrexone were tested for stimulus generalization. Three
of these, naloxone (0.003-0.3 mg/kg), diprenorphine (0.003-0.3
mg/kg) and oxiorphan (0.1-10 mg/kg), produced dose-related
increases in the number of trials completed on the
naltrexone-appropriate lever, and, at the highest dose, stimulus control of
behavior comparable to that produced by the training dose of
naltrexone (fig. 4). In this latter respect, naltrexone was 3 times
more potent than naloxone and diprenorphine, and 100 times
more potent than oxilorphan. These drugs also produced a
dose-related loss of body weight (fig. 4) which correlated with
naltrexone-appropriate responding: naltrexone-rM = 0.74, P <
.001; naloxone-r = 0.55; P < .01; diprenorphine-rM = 0.72, P < .001; oxiorphan-r = 0.42, P < .05. a: 20 Ui > 18 Ui -J l6 LU z l4 0 )( Io Ui
!:
10z
34
a: 0.5 :D 0.3 0 -J I-. 0.2 0 ORAL MORPHINE I ORAL MORPHINE + PELLETS I I I I I I1979 Stimulus Effects of Naltrexone
599
.
. CURVE I 0 CURVE 2 I I I I I S 0.003 0.01 0.03 0.1 DOSE OF NALTREXONE (mg/kg)Fig. 2. Dose-related discriminative stimulus effects of naltrexone (upper
panel), loss of body weight induced by naltrexone (lower panel) and the reproducibility of these eftects, in morphine-dependent rats trained to discriminate between saline and 0. 1 mg/kg of naltrexone. Each
point in the upper panel is the mean number of trials completed on the
naltrexone-appropriate choice lever in a 20-trial session; the remaining
trials were completed on the saline-appropriate lever. The upper and
lower horizontal dashed lines indicate the minimum levels of
discrimi-native responding at which the performance of the animals was
main-tamed with naltrexone (0. 1 mg/kg) and saline, respectively. Each point inthe lower panel is the mean ± 1 S.E.M. percent loss of body weight of the rats during the test sessions; the S.E.M. is not shown where it is less than the radius of the point. Points above S indicate the results of
saline test sessions. Means are based upon one observation in each of
nine rats. Curve 2 was determined 5 to 9 months after curve 1 in the
same animals.
and 17), and returning toward saline levels at 36 hr. The average
number of trials completed on the naltrexone-appropriate lever
at 24 hr of morphine withdrawal is approximately equivalent to
that observed after the administration ofO.01 mg/kg of
naltrex-one to nonwithdrawn rats (fig. 2). In contrast, morphine
with-drawal in rats that had been implanted with morphine pellets
in addition to having had access to the morphine drinking
solution resulted in a relatively rapid increase in
naltrexone-appropriate responding, and a return to saline-appropriate
re-sponding that was still incomplete at 60 hr (fig. 3). The
maxi-mum number of trials completed on the naltrexone-appropriate
lever, 15.6 (based upon individual responses of 8, 14, 18, 18 and
20), again occurred at 24 hr, and is approximately equivalent to
the value obtained after the administration of 0.03 mg/kg of
naltrexone to nonwithdrawn rats (fig. 2).
Changes in body weight were monitored at every time point
only for the rats that had been implanted with morphine pellets.
The rate at which the animals lost weight between two
succes-sive test sessions (percent weight loss per hour) closely
paral-I
I I I
0 6 12 824 36 60
HOURS OF WITHDRAWAL
Fig. 3. Time course of naltrexone-appropriate responding (upper panel)
and rate of loss of body weight (lower panel) following the abrupt
withdrawal of morphine in morphine-dependent rats trained to
discrim-nate between saline and 0.1 mg/kg of naltrexone. Each point in the
lower panel is the mean ± 1 S.E.M. percent loss of body weight of the
animals calculated as change per hour between two successive test
sessions. Means are based upon one observation in each of five rats.
One group of rats had received morphine only in the drinking water;
the other group had received morphine in the drinking water and had
also been implanted s.c. with morphine pellets which were removed
immediately after the test session at 0 hr (see Methods). Other details as in the legend to figure 2.
at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
L..;:-
I I 20 > Ui -I 16 l4 Ui 12 lO z C/) -J <2 I-0 5 C/) U) -i z 0 I I I I I ::.
. NALTREXONE (0.1) #{163}NALOXONE (0.3) . DIPRENORPHINE (0.3) a: 20 Ui 18 -I 16 Ui z 14 0 12 a: 48 z 06 I-U) -I 2 4 a: o I-0 30 60 90 20 50 80600 Gellert and Holtzman Vol. 2 1 1
1 I I L I 1 .J.. .1.. .1. 1.
S NTX 0.003 0.01 0.03 0.1 0.3 I .0 3.0 10
DOSE OF DRUG (mg/kg)
Fig. 4. Dose-related discriminative stimulus effects (upper panel) and
loss of body weight (lower panel) induced by narcotic antagonists that
produce stimulus control of behavior comparable to that of 0.1 mg/kg
of naltrexone in morphine-dependent rats trained to discriminate
be-tween saline and 0.1 mg/kg of naltrexone. Points above S and NTX
indicate the results of saline and naltrexone (0.1 mg/kg) test sessions,
respectively, that were interspersed among the various dose-response
curves. Each point is a mean based upon one observation in each of
15 (naltrexone), six (naloxone) or five rats. Other details as in the
legend to figure 2.
The time course of the discriminative stimulus effects of
doses of three of the drugs that produced comparable stimulus
control ofbehavior-naltrexone (0.1 mg/kg), naloxone (0.3 mg/
kg) and diprenorphine (0.3 mg/kg)-is presented in figure 5. An
average of at least 90% of the trials was completed on the
naltrexone-appropriate choice lever in sessions which began up
to 105 min after the administration of naltrexone, 45 miii after
the administration of naloxone and 15 mm after the
adminis-tration of diprenorphine. Drug-appropriate responding had
re-turned essentially to saline levels in sessions which began 180
mm after the administration of naltrexone and naloxone.
Di-prenorphine clearly had the shortest duration of action of the
three drugs; responding appropriate for saline was in evidence
in sessions which started 105 miii after diprenorphine was
injected. The decrease in body weight induced by the three
drugs ranged between 4.0 ± 0.4 and 5.5 ± 0.6% at each of the
time points.
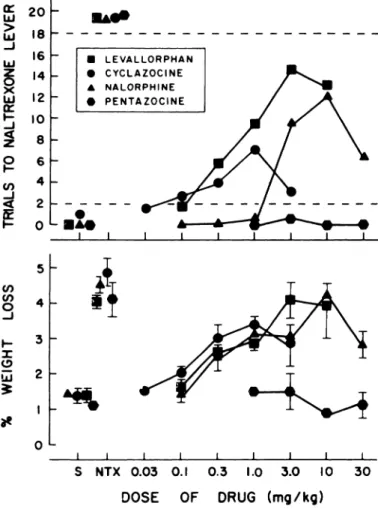
Four drugs with mixed agonist and narcotic antagonist
prop-erties-levallorphan (0.1-10 mg/kg), nalorphine (0.1-30 mg/
kg), cyclazocine (0.3-3.0 mg/kg) and pentazocine (1.0-30 mg/
kg)-failed to produce stimulus control of behavior comparable
to that produced by 0.1 mg/kg ofnaltrexone (fig. 6). Pentazocine
produced only responding appropriate for saline. However, the
other three drugs engendered a biphasic pattern of
naltrexone-- - appropriate responding characterized by a dose-related increase
in the number oftrials completed on the naltrexone-appropriate
lever up to a maximum, and then, at the highest dose, a decline
from this maximum (fig. 6). The maximum number of trials
completed on the naltrexone-appropriate lever averaged 14.5 at
3.0 mg/kg of levallorphan (based upon individual responses of
9, 1 1, 18 and 20), 12.0 at 10 mg/kg of nalorphine (based upon
individual responses of 4, 9, 14, 14 and 19), and 7.0 at 1.0 mg/kg
of cyclazocine (based upon individual responses of 0, 3, 6, 8 and
18). At the next highest dose of each drug, at least half of the
- - - - animals completed fewer trials than these on the
naltrexone-appropriate lever. Although interanimal variability tended to
be high at doses that resulted in intermediate levels of
respond-ing on the naltrexone-appropriate lever, dose-response
relation-ships were generally orderly within each subject. For example,
the number of trials completed on the naltrexone-appropriate
lever by each of the four rats tested with levallorphan at doses
ofO.1, 0.3, 1.0, 3.0 and 10 mg/kg was: 0, 5, 13, 18 and 10 (rat E3);
4, 11, 17, 20 and 16 (rat Gi); 1, 6, 6, and 8 (rat G9); and 1, 0, 2,
11 and 18 (rat G13). From an inspection of figure 6, it is also
apparent that the patterns of weight loss induced by the four
drugs resembled the patterns of naltrexone-appropriate
re-sponding. However, these two variables did not correlate
sig-nificantly for any of the drugs.
Stimulus generalization tests with nonopioid drugs. In
order to assess further the stimulus effects of naltrexone, tests
of stimulus generalization were performed with three
behav-iorally active nonopioid drugs: d-amphetamine (0.1-3.0 mg/kg),
pentobarbital (1.0-17.5 mg/kg) and physostigmine (0.03-1.0
mg/kg). Neither d-amphetamine nor pentobarbital engendered
appreciable naltrexone-appropriate responding when tested up
to the highest dose at which the animals could still complete
MINUTES AFTER INJECTION
Fig. 5. Time course of the discriminative stimulus eftects of naltrexone
(0.1 mg/kg), naloxone (0.3 mg/kg) and diprenorphine (0.3 mg/kg) in
morphine-dependent rats trained to discriminate between saline and
0.1 mg/kg of naltrexone. The animals were injected with the
appropri-ate drug and returned to their home cage until the interval indicated on
the abscissa had elapsed, at which time they were placed in the
experimental chamber and the test session was started. The open
points indicate the results of saline test sessions (1 5-mm pretreatment)
that were interspersed among the time course curves. Each point is a
mean based upon one observation in each of five rats. Each time point
was determined on a separate day. Other details as in the legend to
figure 2.
at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
L EVA L LO R P HA N C YC L A ZOC IN E NALORPHINE PENTAZOCINE S NTX 0.03 0.1 I I I 20 8 16 Ui 14 )c Ui I 2 ‘: 10 8 6 -- - - -I. I #{149} I #{163} I
.
Iphine could be surmounted by increasing the dose of naltrexone.
Figure 9 shows two pairs of stimulus generalization curves for
naltrexone.Thepairontheleft(solidsymbols)wasdetermined
with naltrexone administered in combination with saline: saline
was injected 45 mm before a session and naltrexone (0.003-0.1
mg/kg) was given 30 mm later, or naltrexone (0.01-0.1 mg/kg)
was injected 45 min before a session and saline was injected 15
mm later. The pair of curves on the right (open symbols) was
determined in a similar fashion except that 56 mg/kg of
mor-phine was administered instead of saline. In the presence of 56
mg/kg of morphine given either before or after naltrexone,
(I) increasing the dose of naltrexone to 1.0 mg/kg fully restored
-I
4 2 - - - drug-appropriate responding to the level produced by 0. 1 mg/
0 A#{149}
.
kg of naltrexone. in combination with saline (fig. 9). In contrast,increasing the dose of naltrexone to 1.0 mg/kg in the presence
of 56 mg/kg of morphine did not reinstate maximum weight
5 loss (fig. 9). Percent weight loss and naltrexone-appropriate
(I) 4
I
p- 3L
II
(I
2-I
IL
a1
0 , , , , 0.3 I:
3:0 10 DOSE OF DRUG (mg I kg)Fig. 6. Discriminative stimulus effects (upper panel) and loss of body
weight (lower panel) induced by narcotic antagonists that did not
produce stimulus control of behavior cOmparable to that of 0.1 mg/kg
of naltrexone in morphine-dependent rats trained to discriminate
be-tween saline and 0.1 mg/kg of naltrexone. Points above S and NTX
indicate the results of saline and naltrexone (O1 mg/kg) test sessions, respectively, that were interspersed among the various dose-response
curves. Each point is a mean based upon one observation in each of
four to five rats. Other details as in the legend to figure 2.
the 20-trial session (fig. 7). An average of 6.0 trials was
com-pleted on the naltrexone-appropriate lever after the
administra-tion of 1.0 mg/kg of physostigmine (fig. 7), but, due to the
prominent ataxia associated with this dose, only two out of four
animals completed the session. None of the three drugs induced
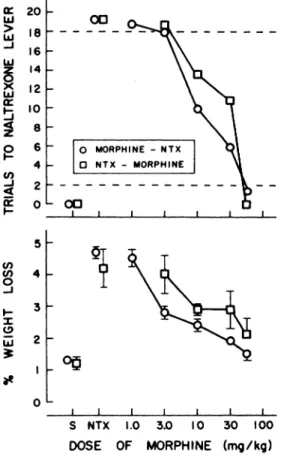
Blockade of the stimulus effects of naltrexone by
mor-significant changes in body weight.
phine. Stimulus control of behavior by 0.1 mg/kg of naltrexone
could be blocked by the administration of morphine either
before or after naltrexone had been given. In the former case,
morphine (1.0-56 mg/kg) was administered 45 mm before the
start of a session and naltrexone (0.1 mg/kg) was given 30 mm
later; in the latter instance, naltrexone (0.1 mg/kg) was injected
45 mm before a session followed 15 mm later by an injection of
morphine (3.0-56 mg/kg). Morphine produced similar
dose-related decreases in the number of trials completed on the
naltrexone-appropriate choice lever under the two treatment
regimens (fig. 8); responding appropriate for saline occurred at
56 mg/kg of morphine in combination with naltrexone.
Mor-phine also attenuated the naltrexone-induced loss of body
weight in a dose-dependent manner (fig. 8).
The blockade of the stimulus effects of naltrexone by
mor-responding were significantly correlated for the
saline-naltrex-one curves (r8 = 0.59, P < .02) but not for the
morphine-naltrexone curves.
Discussion
We have demonstrated that naltrexone, a narcotic antagonist
that is essentially devoid of agonist activity (Martin et al., 1973;
Blumberg and Dayton, 1974), can serve as a discriminative
stimulus in the morphine-dependent rat. The degree of stimulus
control of behavior achieved with saline and 0.1 mg/kg of
naltrexone, as determined by the percentage oftrials completed
on the appropriate choice lever, was comparable to that
de-scnbed for saline and morphine (3.0 mg/kg), the prototypical
narcotic agonist, in this same behavioral paradigm (Shannon
and Holtzman, 1976a, 1977). Moreover, once established, the
E 20 Ui > I 8 w J I 6
1 #{149}
PHYSOSTIGMl 14 #{163}d-AMPHETAMINEL.
PENTOBARBI TAL X I2 Ui Io
-i 4 8 z 46 cn -I 2 --<o
#{149}
I I I I I I I I I S NTX 0.03 0.1 0.3 I .0 3.0 I 0 30DOSE
OF
DRUG
(mg
Ikg)
Fig. 7. Failure of nonopioid psychoactive drugs to produce stimulus
control of behavior comparable to that of 0.1 mg/kg of naltrexone in
morphine-iependent rats trained to discriminate between saline and
o.i mg/kg of naltrexone. Points above S and NTX indicate the results
of saline and naltrexone (0.1 mg/kg) test sessions, respectively, that
were intersper5ed among the various dose-response curves. Each
point is a mean based upon one observation in each of four rats with
the exception that only two out of four rats could be tested after
receiving i.o mg/kg of physostigmine. Other details as in the legend
to figure 2.
1979 Stimulus Effects of Naltrexone 601
at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
I4- l2I-6 4 0 MORPHINE - NTX 0 NIX - MORPHINE 2r- -0L 0 I I I I I I I S NTX 1.0 3.0 10 30 100 20 Ui l8 -I 16 l0 U) I -J 4 5 U) (I) 4 0 -J I-. I (2 at I 0 DOSE OF MORPHINE (mg/kg)
Fig. 8. Dose-related antagonism by morphine of the discriminative
stimulus effects (upper panel) and loss of body weight (lower panel)
induced by 0. 1 mg/kg of naltrexone in morphine-dependent rats
trained to discriminate between saline and 0.1 mg/kg of naltrexone. In
one experiment (morphine-NTX), morphine was administered 45 mm
before a test session and 0. 1 mg/kg of naltrexone was administered
15 mm before the session; in another experiment (NTX-morphine), 0.1
mg/kg of naltrexone was administered 45 mm before a test session
and morphine was administered 30 mm before the session. Points
above S and NTX, indicate the results of saline + saline and saline +
naltrexone (0.1 mg/kg) test sessions, respectively. that were
inter-spersed among the dose-response curves. Each point is a mean based
upon one observation in each of five rats. Other details as in the legend to figure 2.
stimulus control of behavior by saline and naltrexone remained
stable and reproducible over an extended period of time making
it amenable to systematic investigation.
Several lines of evidence suggest that the discriminative
stimuli engendered by naltrexone are related to withdrawal
phenomena. The first, and most compelling, is that following
the abrupt withdrawal of morphine there was an increase in
the number of trials completed on the naltrexone-appropriate
choice lever, the magnitude and time course of which were
directly related to the degree of physical dependence of the
animals. Withdrawal from the morphine drinking solution
re-suited in a relatively modest increase in naltrexone-appropriate
responding concordant with other indications, such as the
al-most complete absence of an observable withdrawal syndrome,
that this regimen of drug administration does not result in a
high degree of physical dependence (Gellert and Holtzman,
1978). However, when orally administered morphine was
sup-plemented by the implantation of morphine pellets, upon the
abrupt withdrawal of morphine all of the rats manifested
in-creases in naltrexone-appropriate responding, and three out of
five completed at least 90% of the trials on the drug-appropriate
lever at the 24-hr time point giving a group mean comparable
to that produced by the administration of 0.03 mg/kg of
nal-trexone to nonwithdrawn morphine-dependent rats. The time
course of changes in naltrexone-appropriate responding
follow-ing the abrupt withdrawal of morphine corresponds well with
the time course of other signs of abrupt morphine withdrawal
in the rat (Akera and Brody, 1968; Khazan and Colasanti, 1971;
Grumbach et al., 1974; Gianutsos et a!., 1975). It is conceivable
that under conditions that result in the development of a
greater degree of physical dependence on morphine than
achieved in this study (e.g., Bl#{228}siget al., 1973; Bhargava, 1977),
naltrexone-appropriate responding following abrupt withdrawal
would reach the same level as that produced by the training
dose of naltrexone in nonwithdrawn morphine-dependent rats.
The second line of evidence is the relationship between the
number of trials completed on the naltrexone-appropriate
choice lever and the loss of body weight by the rats. These two
variables were significantly correlated in every determination
of the dose-response curve for naltrexone (Figs. 2, 4 and 9).
Following the abrupt withdrawal of morphine in rats that had
been implanted with morphine pellets, the hourly rate of weight
loss covaried with the changes in naltrexone-appropriate
re-sponding. Loss of body weight by the morphine-dependent rat
201-> Ui 8 -J 16 Ui l4 Ui 12 a: l0 4 z 8 4 U) 4 5 U) -I I- 3 I (2 at 0 I I I I I I _I S 0.003 0.01 0.03 0. I 0.3 1.0 DOSE OF NALTREXONE (mg/kg)
Fig. 9. Effects of 56 mg/kg of morphine on the discriminative stimulus
effects (upper panel) and loss of body weight (lower panel) induced by
graded doses of naltrexone in morphine-dependent rats trained to
discriminate between saline and 0.1 mg/kg of naltrexone. In one
experiment, either saline (saline-NTX) or 56 mg/kg of morphine
(mor-phine-NTX) was administered 45 mm before a test session and
naltrex-one was administered 1 5 mm before the session; in another
experi-ment, naltrexone was administered 45 mm before a test session and
saline (NTX-saline) or 56 mg/kg of morphine (NTX-morphine) was
administered 30 mm before the session. Points above S indicate the
results of saline + saline test sessions. Each point is a mean based
upon one observation in each of five rats. Other details as in the legend to figure 2.
602
Gellert and Holtzman Vol. 211at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
1979 Stimulus Effects of Naltrexone
603
has consistently been shown to provide a reliable index of the
intensity of the abrupt and antagonist-precipitated withdrawal
syndromes (Akera and Brody, 1968; Wei et al., 1973; Tieger,
1974; Stolerman et al., 1975; Wei and Way, 1975; Gellert and
Sparber, 1977; Geilert and Holtzman, 1978). Thus, the changes
observed in this physiological variable are a source of
indepen-dent corroboration of the validity of the behavioral data.
Our results support the view that the withdrawal syndrome
precipitated by a narcotic antagonist is qualitatively similar to
the syndrome that emerges following abrupt withdrawal of the
drug of dependence except for time course and intensity (Wilder
et al., 1953; Martin, 1967; Jaffe and Martin, 1975). This
rela-tionship between the states of abrupt and precipitated
with-drawal has been difficult to confirm in animals because of the
lack of a specific model for testing the hypothesis.
There is some controversy as to whether or not the
with-drawal syndrome precipitated by a narcotic antagonist can be
suppressed by the administration of additional agonist (Jaffe,
1975). Wilder et al. (1953) reported that the withdrawal
syn-drome precipitated by nalorphine in morphine-dependent
vol-unteers could not be attenuated by the subsequent
administra-tion of large doses of morphine, and Cheney et al. (1972) found
that the administration of levorphanol to
levorphanol-depen-dent mice actually enhanced the ability of naloxone to
precip-itate withdrawal as measured by a decrease in the ED5O for
naloxone-induced withdrawal jumping. In contrast, others have
reported that withdrawal jumping precipitated by naloxone in
morphine-dependent mice could be suppressed by pretreatment
of the mice with morphine or related narcotic analgesics (Iorio
et al., 1975; Brase et al., 1976). In the present study, the
discriminative stimulus effects of the training dose of naltrexone
were blocked in a dose-related manner by morphine
adminis-tered either before or after the naltrexone. The decrease in
drug-appropriate responding was not the consequence of a
nonspecific depression of ongoing behavior by the relatively
high doses of morphine that were used; the behavioral
perform-ance of the rats was unimpaired, with the number of trials
completed on the saline-appropriate lever increasing as the
number of trials completed on the naltrexone-appropriate lever
declined. Stimulus control of behavior by naltrexone in the
presence of morphine was fully restored by increasing the dose
of naltrexone to 10 times the training dose. Thus, the effects of
morphine on the stimulus control of behavior by naltrexone in
the morphine-dependent rat are entirely analogous to the
ef-fects of naltrexone and naloxone on the stimulus control of
behavior by morphine in nondependent subjects (Shannon and
Holtzman, 1976a,b; Schaefer and Holtzman, 1977; J#{228}rbeand
Rollenhagen, 1978) and are suggestive of a competitive type of
antagonism. This finding provides further support for the view
that the discriminative stimuli engendered by the
administra-tion of naltrexone are associated with the displacement of
morphine from its site(s) of action in the body.
In contrast to stimulus control of behavior, maximal loss of
body weight was not re-established when 1.0 mg/kg of
naltrex-one was administered either before or after the administration
of 56 mg/kg of morphine. The dissociation of these two
van-ables-weight loss and naltrexone-appropriate
responding-in-dicate that the loss of body weight per se is not the
discrimi-native stimulus for naltrexone. Whereas loss of body weight is
a useful index of the intensity of the abrupt and
antagonist-precipitated withdrawal syndromes, it becomes a less reliable
measure where additional drugs have been administered for the
purpose of suppressing withdrawal. With appropriate
pharma-cological intervention, it is possible to achieve an almost
corn-plete separation of weight loss and naltrexone-appropriate
re-sponding (unpublished observations).
In tests of stimulus generalization, three other narcotic
an-tagonists produced stimulus control of behavior comparable to
that produced by the training dose of naltrexone. Two of these,
naloxone and diprenorphine, are, like naltrexone, essentially
devoid of agonist activity (Jasinski et al., 1967; Blumberg and
Dayton, 1973; Cowan, 1974). The relative potencies of
naltrex-one, naloxone and diphrenorphine for producing stimulus
con-trot of behavior are consistent with their relative potencies for
inducing morphine withdrawal jumping in the mouse (Cowan,
1976). The shorter duration of action of naloxone relative to
naltrexone in producing stimulus control of behavior is also
consistent with the time course of these drugs for the
precipi-tation of withdrawal in man (Jasinski et al., 1967; Martin et al.,
1973) and in the rhesus monkey (Villarreal and Karbowski,
1974). On the other hand, diprenorphine has usually been found
to have a longer duration of action than naloxone as a morphine
antagonist (Dykstra et al., 1974; Villarreal and Kabowski,
1974), in contrast to the very short duration of stimulus control
of behavior produced by the drug in this study. However,
appropriate quantitative comparisons in rodents are not
avail-able, and seeming discrepancies in relative durations of action
may be related to species differences.
The rats also generalized completely to oxiorphan, a narcotic
antagonist that has weak but detectable agonist activity (Pircio
and Gylys, 1975). However, four other drugs with mixed agonist
and narcotic antagonist properties-levallorphan, cyclazocine,
nalorphine and pentazocine-failed to produce stimulus control
of behavior comparable to that produced by the training dose
of naltrexone, although all except pentazocine engendered some
responding on the naltrexone-appropriate choice lever. The
data in table 1 suggest that the level of naltrexone-appropriate
responding produced by a narcotic antagonist is primarily a
function of the degree of separation of the agonist and
antago-nist
components of action of the drug. If agonist activity isexpressed as the ED5O to inhibit the phenylquinone-induced
writhing response in the mouse, and antagonist activity as the
ED5O to precipitate the morphine withdrawaljumping response,
then the agonist/antagonist potency ratio of the drugs is highly
TABLE 1
Correlation between agonist/antagonist potency ratio of narcotic
antagonists and naltrexone-appropriate choice responding
ED Agonist! MaxImum
Drug Agonist Antagst Anta9onist Naltrexone
ActMty’ Activity” Lever
mg/kg mg/kg
Naltrexone >80c 0.01 “ >1 00 >18
Diprenorphine >1 00’ O.02d >1 >i s
Naloxone >80c O.03d >100 >18
Oxilorphan 12.& O.97c 13.2 >18
Levallorphan 0.29’ O.09d 3.22 14.5
Nalorphine O.77 0.57” 1 .35 12
cyclazocine O.05c 0. 1 3” 0.38 7
Pentazocine 37C 0.37 <2
&ED to inhibit phenylguinone-induced writhing in the mouse.
bED,,, to induce morphine withdrawal jumping in the mouse.
C
Pircio and Gylys (197S).dCowan (1976).
. Cowan (1974). ,Leimgruber et al.(1974).
at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
604
Gellert and Holtzman Vol. 211correlated with the maximum number of trials completed on
the naltrexone-appropriate choice lever (r = 0.98; P < .001).
Many of the signs of the morphine withdrawal syndrome
reflect hyperactivity of the autonomic nervous system (Jaffe,
1975), and the accompanying subjective disturbances bear a
resemblance to the subjective effects produced by pentobarbital
(Haertzen and Hooks, 1969). However, the stimulus effects of
naltrexone were not mimicked by either d-amphetamine, which
produces sympathetic predominence, physostigmine, which
produces parasympathetic predominance, or by pentobarbital.
Thus, stimulus control of behavior by naltrexone in
morphine-dependent subjects, like that of other drugs in nondependent
subjects, appears to be relatively specific with respect to
phar-macologic class, and is not duplicated by drugs that produce
only some of the effects which may compose the discriminative
stimulus complex of the training drug.
Study of antagonist-precipitated narcotic withdrawal
phe-nomena has been hampered by the lack of appropriate animal
models. In the rat, the signs of morphine withdrawal that are
manifested following administration of a narcotic antagonist
can vary considerably depending upon such factors as the
degree of physical dependence and the dose of the antagonist
(Bl#{228}siget al., 1973; Gellert and Holtzman, 1978). Even those
signs that are reliably associated with morphine withdrawal, for
example, loss of body weight, are by no means unique to the
state of morphine withdrawal, and are subject to nonspecific
and often unpredictable modification by drugs. Our initial
find-ings indicate that the stimulus control ofbehavior by naltrexone
in the morphine-dependent rat has many of the characteristics
previously demonstrated for the stimulus control of behavior
by morphine in the nondependent rat (Shannon and Holtzman,
1976a, 1977): long-term stability and reproducibility, orderly
dose- and time-effect relationships, surmountable blockade by
an “antagonist,” and pharmacologic specificity. Accordingly,
this drug discrimination paradigm may afford a specific animal
model for studying a component of the morphine withdrawal
syndrome that is directly relevant to the perpetuation of chronic
drug use in physically dependent human addicts.
References
AKERA, T. AND BRODY, T. M.: The addiction cycle to narcotics in the rat and its relation to catecholamines. Biochem. Pharmacol. 17: 675-688, 1968. Bj..Asic, J., HERZ, A., REINHOLD, K. AND ZIEGLGANSBERGER, S.: Development of
physical dependence on morphine in respect to time and dosage and quantifi-cation of the precipitated withdrawal syndrome in rats. Psychopharmacologia 33: 19-38, 1973.
BHARGAVA, H. N.: Rapid induction and quantitation of morphine dependence in the rat by pellet implantation. Psychopharmacology 52: 55-62, 1977.
BLUMBERG, H. AND DAYTON, H. B.: Naloxone and related compounds. In Agonist
and Antagonist Actions of Narcotic Analgesic Drugs, ed. by H. W. Kosterlitz, H. 0. J. Collier and J. E. Villarreal, pp. 110-119, University Park Press, Baltimore, 1973.
BLUMBEEG, H. AND DAYTON, H. B.: Naloxone, naltrexone, and related
noroxy-morphines. In Narcotic Antagonists, ed. by M. C. Braude, L. S. Harris, E. L May, J. P. Smith and J. E. Villarreal, pp. 33-43, Raven Press, New York, 1974. BRASE, D. A., IWAMOTO, E. T., LoH, H. H. AND WAY, E. L: Reinitiation of sensitivity to naloxone by single narcotic injection in post-addicted mice. J. Pharmacol. Exp. Ther. 197: 317-325, 1976.
CHENEY, D. L, JUDSON, B. A.AND GOLDSTEIN, A.: Failure ofan opiate to protect
mice against naloxone-precipitated withdrawal. J. Pharmacol. Exp. Ther. 182: 189-194, 1972.
COLPAERT, F. C.: Narcotic cue and narcotic state. Life Sci. 20: 1097-1108, 1977.
COWAN, A.: Evaluation in nonhuman primates: Evaluation of the physical de-pendence capacities of oripavine.thebaine partial agonists in pates monkeys. In Narcotic Antagonists, ed. by M. C. Braude, L S. Harris, E. L May, J. P. Smith and J. E. Villarreal, pp. 427-438, Raven Press, New York. 1974.
COWAN, A.: Use of the mouse jumping test for estimating antagonist potencies of
morphine antagonists. J. Pharm. Pharmacol. 28: 177-183, 1976.
Dyicgrs, L A., McMILLAN, D. E. AND HARRIS, L. S.: Antagonism of morphine by long acting narcotic antagonists. Psychopharmacologia 39: 151-162, 1974. FRASER, H. F.: Methods for assessing the addiction liability of opioids and opioid
antagonists in man. In The Addictive States, ed. by A. Wikier, pp. 176-187, Williams and Wilkins, Baltimore, 1968.
GELLERT, V. F. AND HOLTZMAN, S. G.: Development and maintenance of mor-phine tolerance and dependence in the rat by scheduled access to morphine drinking solutions. J. Pharmacol. Exp. Ther. 205: 536-546, 1978.
GELLERT, V. F. AND SPARBER, S. B.: A comparison of the effects of naloxone upon body weight loss and suppression of fixed-ratio operant behavior in morphine-dependent rats. J. Pharmacol. Exp. Ther. 201: 44-54, 1977.
GIANuTsos, G., DRAWBAUGH, R., HYNES, M. AND LAL, H.: The narcotic with.
drawal syndrome in the rat. In Methods in Narcotic Research, eli by S.
Ehrenpreis and A. Neidle, pp. 293-309, Marcel Dekker, New York, 1975.
GIBSON, R. D. AND TINGSTAD, J. E.: Formulation of a morphine implantation
pellet suitable for tolerance-physical dependence studies in mice. J. Pharm. Sci.
59: 426-427, 1970.
GRUMBAcH, L., SHELOFSKY, M. AND BosToN, J. E.: The kinetics of physical
dependence to morphine in the rat. Pharmacology (Basel) 11: 38-51, 1974. HAERTZEN, C. A.: Subjective effects of narcotic antagonists. In Narcotic
Antag-onists, ed. by M. C. Braude, L S. Harris, E. L. May, J. P. Smith and J. E.
Villarreal, pp. 383-398, Raven Press, New York, 1974.
HAERTZEN, C. A. AND HOOKS, N. T.: Changes in personality and subjective experience associated with the chronic administration and withdrawal of opi-ates. J. Nerv. Ment. Dis. 148: 606-614, 1969.
HAYS, W. L.: Statistics for the Social Sciences, Holt, Rinehart and Winston, New York, 1973.
HIRSCHHORN, I. .AND ROSECRANS, J. A.: Generalization of morphine and lysergic
acid diethylamide (LSD) stimulus properties to narcotic analgesics. Psycho-pharmacology 47: 65-69, 1976.
HOLTZMAN, S. G.: Effects of morphine and narcotic antagonists on avoidance behavior of the squirrel monkey. J. Pharmacol. Exp. Ther. 196: 145-155, 1976.
loiuo, L. C., DEACON, M. A. AND RYAN, E. A.: Blockade by narcotic drugs of naloxone-precipit.ated jumping in morphine-dependent mice. J. Pharmacol. Exp. Ther. 192: 56-63, 1975.
JA.FFE, J. H.: Drug addiction and drug abuse. In The Pharmacological Basis of
Therapeutics, 5th ed., ed. by L. S. Goodman and A. Gilman, pp. 284-324, The Macmillan Company, New York, 1975.
JAFFE, J. H. AND MARTIN, W. R.: Narcotic analgesics and antagonists. In The Pharmacological Basis of Therapeutics, 5th ed., ed. by L. S. Goodman and A. Gilman, pp. 245-283, The Macmillan Company, New York, 1975.
JARBE, T. U. C. AND ROLLENHAGEN, C.: Morphine as a discriminative cue in gerbils: Drug generalization and antagonism. Psychopharmacology 58: 271-275, 1978.
JASINSKI, D. R.: Effects in man of partial morphine agonists. In Agonist and
Antagonist Actions of Narcotic Analgesic Drugs, ed. by H. W. Kosterlitz, H. 0. J. Collier, and J. E. Villarreal, pp. 94-103, University Park Press, Baltimore, 1973.
JASINSKI, D. R.: Assessment of the abuse potentiality of morphine-like drugs (methods used in man). In Handbook of Experimental Pharmacology, vol. 45, Drug Addiction, ed. by W. R. Martin, pp. 197-258, Springer-Verlag, Berlin, 1977.
JASINSKI, D. R, MARTIN, W. R. AND HAERTZEN, C. A.: The human pharmacology
and abuse potential of N-allylnoroxymorphone (naloxone). J. Pharmacol. Exp. Ther. 157: 420-426, 1967.
KHAZAN, N. AND COLASANTI, B.: Decline in the mean integrated
electroenceph-alogram voltage during morphine abstinence in the rat. J. Pharmacol. Exp. Ther. 177: 491-499, 1971.
LAL, H., GIANUTSOS, G. AND MIKSIC, S.: Discriminable stimuli produced by
analgesics. In Discriminative Stimulus Properties of Drugs, ed. by H. Lal, pp. 23-45, Plenum Press, New York, 1977.
LEIMGRUBER, W., M0HAc5I, E., BARUTH, H. AND RANDALL, L. 0.: Levallorphan and related compounds. In Narcotic Antagonists, ed. by M. C. Braude, L. S.
Harris, E. L May, J. P. Smith and J. E. Villarreal, pp. 45-50, Raven Press,
New York, 1974.
MARTIN, W. R.: Opioid antagonists. Pharmacol. Rev. 19: 463-521, 1967.
MARTIN, W. R. AND FRASER, H. F.: A comparative study of physiological and subjective effects of heroin and morphine administered intravenously in post-addicts. J. Pharmacol. Exp. Ther. 133: 388-399, 1961.
MARTIN, W. R., JA5INSKI, D. R. AND MANSKY, P. A.: Naltrexone, an antagonist
for the treatment of heroin dependence. Arch. Gen. Psychiatry 28: 784-791, 1973.
O’BRIEN, C. P.: Experimental analysis of conditioning factors in human narcotic addiction. Pharinacol. Rev. 27: 533-543, 1975.
PIRCIO, A. W. AND GYLYS, J. A.: Oxiorphan
(l-N-cyclopropylmethyl-3,14-dihy-droxymorphinan): A new synthetic narcotic antagonist. J. Pharmacol. Exp.
Ther. 193: 23-34, 1975.
SCHAEFER, G. J. AND HOLTZMAN, S. G.: Discriminative effects of morphine in the squirrel monkey. J. Pharmacol. Exp. Ther. 201: 67-75, 1977.
SHANNON, H. E. AND HOLTZMAN, S. G.: Evaluation of the discriminative effects of morphine in the rat. J. Pharxnacol. Exp. Ther. 198: 54-65, l976a.
SHANNON, H. E. AND HOLTZMAN, S. G.: Blockade of the discriminative effects of morphine in the rat by naltrexone and naloxone. Psychopharmacologia 50:
119-124, 1976b.
SHANNON, H. E. AND HOLTZMAN, S. G.: Further evaluation of the discriminative effects of morphine in the rat. J. Pharmacol. Exp. Ther. 201: 55-66, 1977.
SIEGEL, S.: Nonparametric Statistics for the Behavioral Sciences, McGraw-Hill Book Company, New York, 1956.
at ASPET Journals on May 11, 2016
jpet.aspetjournals.org
1979 Stimulus Effects of Naltrexone 605
STOLERMAN, I. P., JOHNSON, C. A., BUNKER, P. AND JARVIK, M. E.: Weight loss and shock-elicited aggression as indices of morphine abstinence in rats. Psy-chopharmacologia 45: 157-161, 1975.
TEICER, D. G.: Induction of physical dependence on morphine, codeine and meperidine in the rat by continuous infusion. J. Pharmacol. Exp. Ther. 190: 408-415, 1974.
VILLARREAL, J. E. AND KARBOwSKI, M. G.: The actions of narcotic antagonists in morphine-dependent rhesus monkeys. In Narcotic Antagonists, ed. by M. C.
Braude, L. S. Harris, E. L. May, J. P. Smith and J. E. Vilarreal, pp. 273-289, _________________________________________________________ Raven Press, New York, 1974.
WE!, E., LOH, H. H. AND WAY, E. L.: Quantitative aspects of precipitated reprint requests to: Stephen G. Holtzman, Ph.D., Department of
Phar-abstinence in morphine-dependent rats. J. Pharmacol. Exp. Ther. 184: 398- macology, Emory University School of Medicine, Atlanta, GA 30322. 403, 1973.
WEI, E. AND WAY, E. L.: Application of the pellet implantation technique for the assessment of tolerance and physical dependence in the rodent. In Methods in
Narcotic Research, ed. by S. Ehrenpreis and A. Neidle, pp. 243-259, Marcel Dekker, New York, 1975.
WIKLER, A., FRASER, H. F.AND LSBELL, H.: N-allylnormorphine: Effects ofSingle
doses and precipitation of acute “abstinence syndromes” during addiction to
morphine methadone or heroin in man (post-addicts). J. Pharmacol. Exp. Ther. 109: 8-20, 1953.