Regulation of G protein signaling by G protein alpha subunit phosphorylation
Sarah T. Clement
A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Department of Biochemistry and Biophysics
Chapel Hill 2013
Approved by:
Henrik Dohlman, Ph.D. John Sondek, Ph.D. Jean Cook, Ph.D.
ABSTRACT
SARAH T. CLEMENT: Regulation of G protein signaling by G protein alpha subunit phosphorylation
(Under the direction of Dr. Henrik G. Dohlman)
Cells respond to stimuli by detecting extracellular signals in complex
environments through cell membrane protein receptors. G protein coupled receptors (GPCRs) comprise the largest family of plasma membrane receptors and transduce signals from an array of stimuli including light, odors, hormones and
neurotransmitters. GPCR-mediated pathways are important in many physiological functions and are targeted by numerous pharmaceuticals. Thus a comprehensive understanding of the regulation of GPCR-mediated pathway components is
necessary to achieve full therapeutic effectiveness and discover new drug targets. This work examines how GPCR-mediated signaling pathways are modulated in the context of changes in the cell-cycle and changes in nutrient availability.
In this thesis, we present studies to identify new regulators of G protein signaling. Specifically, we show that the G protein α subunit, Gpa1, is
phosphorylated and degraded in a cell-cycle dependent manner. In addition, we demonstrate that Gpa1 is phosphorylated in a low glucose-dependent manner, which leads to a reduction in signal transduction. These findings reveal new
ACKNOWLEDGEMENTS
The completion of this thesis would not have been possible without the help
of many people. First, I would like to thank my advisor Henrik Dohlman. He has
provided much encouragement throughout my years in the lab and supported my
ideas and pursuits. He has taught me how to become a better scientist and scientific
writer and I greatly appreciate his patience. I would also like to thank the members of
my committee, John Sondek, Pat Brennwald, Jean Cook, and Ken Harden, for
helpful feedback on this work.
I would like to thank Mike Lee for early contributions to this work and his
mentorship when I first joined the lab. Thanks to Matt Torres and Dan Isom for help
on critical experiments in this thesis work and helpful discussions. Thanks to Gauri
Dixit for her contributions to this work using her excellent microscopy skills. Thanks
to Holly Lien for her hard work and dedication to this project. Thanks to Jan Jones,
Steve Cappell, and Jill Hurst for providing helpful feedback with experiments and
writing. Thanks to all members of the Dohlman lab, past and present, for making the
lab a great place to work.
Specifically for the work in Chapter II, the co-contributors and I thank Daniel
Isom for constructive discussions and technical assistance with analysis of G protein
point mutants, Candice Carlile and Jean Cook for the cdc6-1 yeast strain, and
Specifically for the work in Chapter III, the co-contributors and I thank Martin
Schmidt for the Sak1 and Tos3 plasmids used for in vitro kinase assays, Mike Lee
for his early contributions to the analysis of Reg1, Holly Lien for performing the
mating efficiency assays, and Matt Torres for valuable advice and guidance.
Thanks to my friends and family for their constant support. My graduate
career has been a rollercoaster of successes and failures, stress and excitement,
and highs and lows, and I appreciate everyone who’s helped me along the way.
Thanks to my friends in my incoming graduate class – Jon Edwards, Pat Lackey,
Ben Stranges, and Aaron Hobbs. Thanks to my family – Mom, Dad, Melissa, Dan,
Grandpa, Aunt Jeanne and Uncle George, and the Clement family – they never
doubted that I would finish this work. Lastly I would especially like to thank Brian
Clement. He knows how much his support has carried me through and made all of
TABLE OF CONTENTS
LIST OF TABLES...viii
LIST OF FIGURES... ix
LIST OF ABBREVIATIONS AND SYMBOLS... xi
CHAPTERS I. Introduction ...1
Heterotrimeric G proteins ...2
Yeast as a model organism for studying G protein signaling ...4
The pheromone response pathway...5
Energy-sensing AMPK ...9
Cross talk between signaling pathways ...12
Summary ...14
Chapter I References ...15
II. Cell-cycle dependent phosphorylation and ubiquitination of a G protein alpha subunit...21
Introduction...22
Results...24
Discussion ...36
Experimental Procedures ...43
III. Regulation of G protein signaling by Snf1/AMPK kinases ...58
Introduction...59
Results...61
Discussion ...72
Experimental Procedures ...77
Chapter III References ...85
VI. Conclusions and General Discussion ...90
Conditional Gpa1 phosphorylation ...91
Signal transduction in human health and disease...96
Conclusions...97
Experimental Procedures ...99
LIST OF TABLES
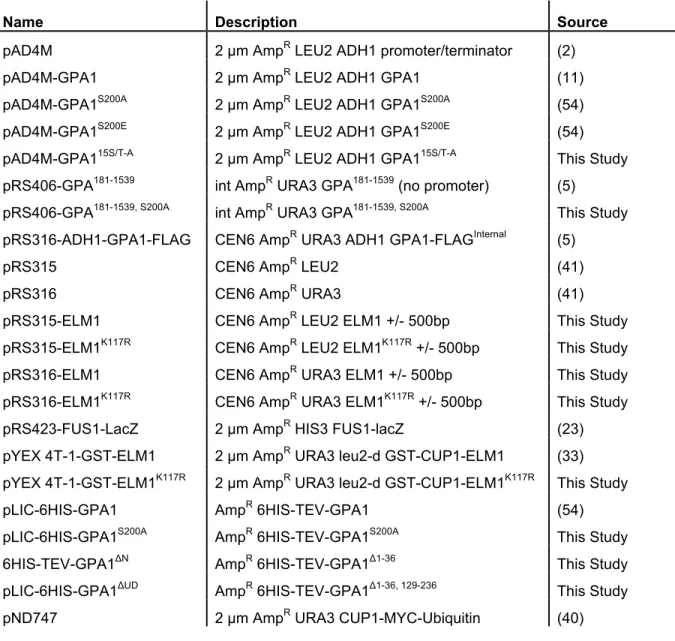
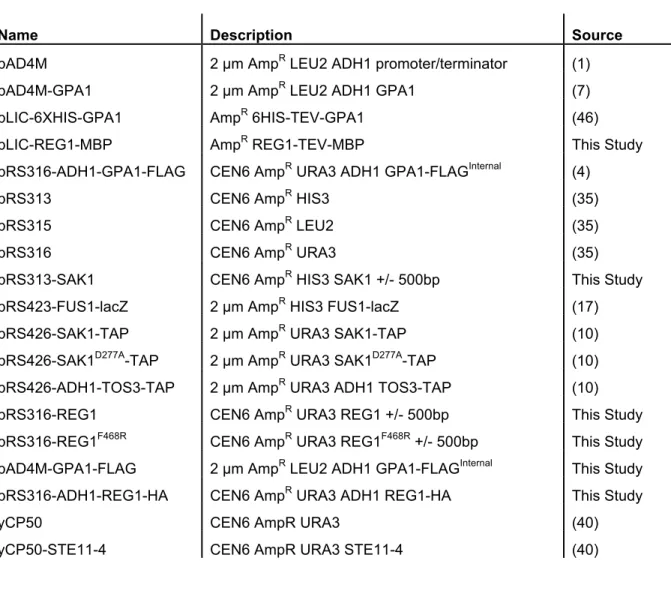
Table 2.1 Plasmids used in Chapter II ...51
Table 2.2 Oligonucleotide primer sequences in Chapter II ...52
Table 3.1 Strains used in Chapter III ...82
Table 3.2 Plasmids used in Chapter III ...83
LIST OF FIGURES
Figure 1.1 Heterotrimeric G proteins signaling cascades ...3
Figure 1.2 The yeast pheromone response pathway...6
Figure 1.3 Regulation of Gpa1 via post-translational modifications and effectors ...8
Figure 1.4 Regulation of the AMP-activated kinase, Snf1...10
Figure 2.1 Phosphorylation induces an electrophoretic mobility shift in Gpa1...25
Figure 2.2 Elm1 is a candidate Gpa1 kinase ...27
Figure 2.3 Yeast kinome screen reveals Elm1 as a Gpa1 kinase...29
Figure 2.4 Elm1 is necessary and sufficient for Gpa1 phosphorylation ...31
Figure 2.5 Gpa1 phosphorylation is cell cycle-dependent...33
Figure 2.6 Elm1 is required for Gpa1 polyubiquitination ...35
Figure 2.7 Gpa1 polyubiquitination by SCFCdc4 is cell cycle-regulated ...37
Figure 2.8 Model of cell cycle Gpa1 regulation ...42
Figure 3.1 Gpa1 is phosphorylated in low glucose conditions ...62
Figure 3.2 Kinases Sak1 and Tos3 and phosphatase Reg1 bind Gpa1 ...65
Figure 3.3 Snf1 kinases prevent early mating response while the Snf1 phosphatase is required for maximal mating response...67
Figure 3.4 Crosstalk between mating and glucose sensing pathways...69
Figure 3.5 Mating transcriptional response and epistasis in low glucose. ...71
Figure 3.6 Shmoo formation and mating are impaired during glucose limitation ....73
Figure 3.7 Gpa1 phosphorylation is not affected by nucleotide binding, but is dampened in the absence of glucose sensors Rgt2 and Snf3...75
Figure 4.2 Gpa1 is differentially phosphorylated under conditions of
LIST OF ABBREVIATIONS AND SYMBOLS
6XHIS Six histidine residues (used as an affinity tag)
A600nm Absorbance measured at 600nm wavelength
Ala or A Alanine
AMP adenosine monophosphate
AMPK AMP-activated protein kinase
APT1 Acyl-Protein Thioesterase
cAMP cyclic adenosine monophosphate
CDC4 Cell Division Cycle
Δ Deletion
DAG Diacyl glycerol
DIC Differential interference contrast
Dox Doxycycline
E. coli Escherichia coli
ELM1 ELongated Morphology
FDG Fluoroscein di-galactoside
FLAG Flag epitope tag (DYKDDDDK)
FUS3 cell FUSion
G protein Guanine nucleotide binding protein
G6PDH Glucose-6-phosphate dehydrogenase
Gα G protein alpha subunit
GAL Galactose
Gβ G protein beta subunit
GDP Guanosine diphosphate
GEF Guanine nucleotide exchange factor
GET3 Guided Entry of Tail-anchored proteins
Gγ G protein gamma subunit
GLC7 GLyCogen
Glu or E Glutamate
GPA1 G Protein alpha subunit
GPCR G protein-coupled receptor
GST Glutathione S-transferase (used as an affinity tag)
GTP Guanosine triphosphate
HA Human influenza hemagglutinin (used as an affinity tag)
HU hydroxyurea (arrests cells in S phase)
IB Immunoblot
IP Immunoprecipitate
IP3 Inositol 1,4,5-trisphosphate
kDa kilo-Dalton (unit of mass)
KSS1 Kinase suppressor of Sst2 mutations
NMT1 N-Myristoyl Transferase
MAPK Mitogen-activated protein kinase
MAPKK Mitogen-activated protein kinase kinase
MAPKKK Mitogen-activated protein kinase kinase kinase
NZ nocodazole (arrests cells in G2/M phase)
PCR Polymerase chain reaction
PIP Phosphatidylinositol 4,5-bisphosphate
PMA1 Plasma Membrane ATPase
PP1 Protein Phosphatase Type 1
REG1 REsistance to Glucose repression
RGS Regulator of G protein signaling
RSP5 Reverses Spt- Phenotype
SAK1 Snf1 Activating Complex
S. cerevisiae Saccharomyces cerevisiae
SCF Skp, Cullin, F-box
SDS-PAGE Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
Ser or S Serine
SIR2 Silent Information Regulator, Sirtuin
SNARE SNAP (Soluble NSF Attachment Protein) REceptor
SNF1 Sucrose Non-Fermenting
SST2 Supersensitive to pheromone
STE STErile
SWI/SNF SWitch/Sucrose Non-Fermenting)
TAP Tandem affinity purification (used as an affinity tag)
TCA Trichloroacetic acid
TetO Tetracycline-repressible promoter
TOS3 Target Of SBF transcription complex
Ub Ubiquitin
CHAPTER I
INTRODUCTION*
Cells respond to stimuli by detecting their extracellular environment through cell membrane protein receptors. One such family of receptors, the G protein
coupled receptors (GPCRs), is responsible for the transduction of numerous signals including light, odors, hormones and neurotransmitters. GPCRs are important
mediators of signaling pathways in numerous tissues in the body, including the heart and brain, and often aberrant signaling can lead to disease. GPCRs are commonly used drug targets, making up a third of drugs on the market (15, 50, 62). These drugs aimed at modulating signaling pathways include depressants,
anti-histamines, and beta-blockers for the treatment of many diseases and ailments such as heart disease, schizophrenia, and allergies. As research leads to a more
comprehensive understanding of GPCR-mediated protein signaling pathways, it is expected to lead to the identification of new drug targets and the generation of new pharmaceuticals and alternative drug regimens.
Cells often encounter multiple extracellular cues, which can often be in conflict with each other. While there is much known about how extracellular signals activate GPCRs and downstream G protein signaling, less is known about how GPCR-mediated pathways are altered when multiple extracellular signals are
received and concurrent pathways are activated. This thesis work examines the convergence of two signaling pathways long thought to act separately: a GPCR-mediated pathway and the metabolism-regulating AMP-activated protein kinase (AMPK) pathway. More broadly, this work demonstrates the importance of crosstalk between signaling pathways to coordinate and elicit an appropriate cellular response when cells encounter complex extracellular environments.
Heterotrimeric G proteins
GPCRs are ubiquitously found in yeast, plants and animals. GPCRs are coupled to heterotrimeric G proteins, which consist of a G protein α subunit and a G protein βγ obligate dimer. Once an agonist binds to the GPCR, the receptor
becomes activated and acts as a guanine nucleotide exchange factor (GEF) leading to the exchange of GDP for GTP on the Gα subunit. The activation of Gα leads to dissociation from Gβγ, and both are free to bind downstream effectors leading to consequences such as the production of second messengers and the activation of protein kinase cascades. Signaling stops when GTP is hydrolyzed to GDP, which is often accelerated through GTPase activating proteins (GAPs), and the subunits rebind to form the heterotrimeric complex (53).
!"#$ !"#$%&'()(
!"#$%&'()(%%&'(')*()+,')+-%!%.)*('+/%0+1/23+/1%-20-24'05%6'7'/%()2/0,',8)2/'%!% .)*('+/%-*9.3'4%)'-'.(*)0%2)'%-*9.3'4%(*%:'(')*()+,')+-%!%.)*('+/05%!%.)*('+/%23.:2% 0989/+(0%8'3*/1%(*%.)*('+/%;2,+3<%098(<.'0%2/4%,'4+2('%4+;;')'/(%-'33932)%
)'0.*/0'05%!%.)*('+/0%()2/0,+(%0+1/230%(*%4*=/0()'2,%.2(:=2<%-*,.*/'/(0%8<% .)*49-+/1%0,233%,*3'-93'%0'-*/4%,'00'/1')0%09-:%20%->?"@%4+2-<313<-')*3%AB>!C% 2/4%+/*0+(*3%()+.:*0.:2('%AD"EC%*)%2-(+72(+/1%.)*('+/0%09-:%20%?>"%F+/20'0%2/4% 0,233%!G"20'0%3+F'%$:*5%B*=/0()'2,%0+1/23+/1%'7'/(0%3'24%(*%()2/0-)+.(+*/23% '7'/(0%(*%,'4+2('%2/%2..)*.)+2('%-'33932)%)'0.*/0'5
! "
?>"H >#
>G"
->?"
"I#
"D"J
D"E B>!
$:*!KL $:* !#+
! "
!#0
! "
!#M
! "
phospholipase C and the production of diacylglycerol (DAG) and inositol
triphosphate (IP3) second messengers. Finally, the Gα12/13 family members can regulate small GTPase Rho activity via activation of RhoGEF proteins. While it is well-known that extracellular GPCRs are good targets for pathway modulation, the intracellular Gα subunits are poised to be good targets for regulation as the first components in G protein signaling cascades.
Yeast as a Model Organism for Studying G Protein Signaling
Saccharomyces cerevisiae has proven to be a very useful model organism for studying conserved signaling pathways. In 1996, Saccharomyces cerevisiae
became the first eukaryotic genome to be sequenced (22), thus facilitating genome-based screens and studies. The founding members of many important proteins have been first found independently in yeast including the master metabolic regulator AMP-activated kinase (Snf1) (8), regulator of G protein signaling protein (Sst2) (13), aging-related sirtuin protein (Sir2) (33), p21-activated kinase (Ste20) (37), vesicle-fusion SNARE proteins (48), cell division cycle proteins (Cdc) (24), and the
nucleosome-remodeling SWI/SNF complex (61). Yeast is tractable to numerous genetic techniques, including complete gene deletions and targeted point mutations of endogenous proteins. In 2000, the Saccharomyces Genome Deletion Project released the Yeast Knock-Out deletion collection, which included almost 82% of all yeast genes (20, 61).
16 Gα subunits, yeast contain 3 GPCRs and 2 Gα subunits (4, 27). Thus yeast contains a simplified system for studying evolutionarily similar proteins and their signaling responses. Diploid and haploid yeast can proliferate stably, which easily allow the study of recessive gene mutations. Haploid yeast exist in two mating types, type a cells (MATa) and type alpha cells (MATα). Small peptide pheromones are released when cells of opposite mating type are in proximity to one another. MATa releases a-factor pheromone and MATα releases α-factor pheromone. The
pheromone ligands can bind to GPCRs on opposite cell mating types and activate G protein signaling cascades to prepare the cells for fusion to form an a/α diploid cell (3).
The Pheromone Response Pathway
The pheromone response or mating pathway in the yeast Saccharomyces cerevisiae is a prototypical GPCR-mediated pathway. The pheromone response pathway has been used as a model to study signaling events, as many proteins in this pathway are functionally conserved in higher organisms. The response to pheromone ligand α-factor binding the GPCR Ste2 on a MATa cell, includes
activation of a Gαi protein Gpa1 and a protein kinase cascade comprised of Ste20, Ste11, Ste7 and two partially redundant MAP kinases Fus3 and Kss1. The activation of the MAP kinases results in upregulated mating gene transcription, morphological changes, and growth arrest for the haploid cells to prepare for fusion and form a diploid cell (14) (Fig. 1.2).
!
"
#
!"#$%&'
&()''
&()*
$+,-./#000
./#00
./#0
&()12
&()'1
$34'
&()5
!#67
!"#$%&'()*
!83'
!9#&()1
#:;3<(=4>8?)4=@=A)6B<C1
.3(DAE>83(?F3G>=+(<=@)
%84)E+H3()B>E)A)>(43A,<4D8(D=A
6)HH>8=H34DI3(D=A
!4=F(?>344),(>DA>!'>8?3,)
!"#$%&'()*>>9?)>G)3,(>8?)4=@=A)>4),8=A,)>83(?F3GJ>K38H=DB>G)3,(>4)H)3,)>,@3HH>
8)8(DB)>8?)4=@=A),L>F?D<?>MDAB>(=>=88=,D()>@3(DAE>(G8)>!>84=()DA:<=+8H)B>4)<)8: (=4,>3(>(?)>8H3,@3>@)@M43A)J>N?)A>8?)4=@=A)>MDAB,>(=>(?)>4)<)8(=4>(?)>!#> 84=()DA>,+M+AD(>4)H)3,),>!"#>3AB>MDAB,>(=>!9#L><3+,DAE>(?)>?)()4=(4D@)4>(=>BD,,=: <D3()>DA(=>;4))>!#>O!83'P>3AB>!!">BD@)4>O&()CQ&()'RPJ>$4))>!!">3<(DS3(),>(?)> ./#0><3,<3B)><+H@DA3(DAE>DA>(?)>8?=,8?=4GH3(D=A>=;>./#0>$+,-J>$+,->3<(DS3(),> B=FA,(4)3@>);;)<(=4,>DA<H+BDAE>&()'1>3AB>$34'J>&()'1>D,>3>(43A,<4D8(D=A>;3<(=4>(?3(> 4)E+H3(),>@3(DAE:,8)<D;D<>E)A),>DA<H+BDAE>$+,'J>$34'>DA?DMD(,>3><G<HDA:B)8)AB)A(> TDA3,)>(=>@)BD3()><)HH><G<H)>344),(>DA>(?)>!'>8?3,)J
&()C
&()'R
known for years, questions still remain about how sensitivities, amplitudes, and temporal aspects of adaptive responses are controlled. Of particular interest is when concurrent extracellular signals received by the cell have potentially contradictory outcomes.
This work focuses on the regulation and modification of the G protein α subunit in the mating pathway as a means of pathway regulation. The list of G protein-binding partners and post-translational modifications has expanded in recent years. It is well known that Gpa1 binds to a pheromone receptor Ste2 (5), the Gβγ subunit pair Ste4/Ste18 (11), and a GTPase accelerating protein Sst2 (2). Other identified binding partners for Gpa1 include a non-receptor exchange factor (Get3) and two subunits of the phosphatidylinositol 3-kinase (Vps15 and Vps34).
Additionally, binding partners of Gpa1 include the enzymes responsible for the addition or removal of post-translational modifications. These include a
N-myristoyltransferase Nmt1 (17), a palmitoyltransferase (32), a de-palmitoylating acyl-protein thioesterase Apt1 (16), a ubiquitin ligase Rsp5 (Gpa1 monoubiquitination) (57), and a ubiquitin ligase SCF Cdc4 (Gpa1 polyubiquitination) (7) (Fig. 1.3). These modifications are often dynamic and directly contribute to the ability of Gpa1 to maintain signaling fidelity during mating response.
!"#$
!"#$
!"#$
!"#$%&'()*
!"#$
%&'(
!"#$%&'()*))*'+,-#&./0)/1)!"#$)2.#)"/3&4&5#03-#&./0#-)6/7.1.8#&./03)#07)'11'8&/539)
!"#$):.073)+,#0/3.0')0,8-'/&.7'3;)<=.8=)7.5'8&->)#11'8&)!"#$?'11'8&/5).0&'5#8&./03) #07)-/8#-.@#&./09)!AB4:/,07)!"#$)"5'1'5'0&.#-->):.073)&=')!)"5/&'.0):'&#?+#66#) C%&'D?%&'$EF)7.6'5)#07)G"3$H9)!IB4:/,07)!"#$)"5'1'5'0&.#-->):.073)G"3JD9)!"#$) &5#03.&./0)3&#&'):.073)&/)!IB#3'4#8&.2#&.0+)"5/&'.0)C%3&(F9)K,8-'/&.7'415'')!"#$) "5'1'5'0&.#-->):.073)&=')!BL*)C%&'(F)#07)0/045'8'"&/5)+,#0.0')0,8-'/&.7')
'M8=#0+')1#8&/5)!'&J9)N0)#77.&./0;)!"#$).3)6/7.1.'7):>)0,6'5/,3)"/3&4&5#03-#&./0#-) 6/7.1.8#&./03;)<=.8=)/1&'0)#11'8&)!"#$)-/8#-.@#&./09
O:)O:) O:)O:)
B)
%&'D)%&'$E) !'&J)!'&J)
!"#$
!"#$
!"#$
!"#$
!IB
O:
6/0/4,:.P,.&.0#&./0 "/->4,:.P,.&.0#&./0
0,8-'/&.7'4:.07.0+
%3&(
%LQ
L78D
*3"H
-.".7)6/7.1.8#&./0
K6&$
R-6$
G"3$H
G"3JD
:.07.0+)"#5&0'53
interaction with RGS (21), loss of guanine nucleotide binding (47), receptor desensitization (10, 52), and loss of interaction with downstream effectors (41).
Energy-sensing AMPK
All cells and organisms must monitor and maintain their energy levels for survival. AMP-activated kinases (AMPKs) are major metabolic regulators in
eukaryotes. These kinases serve as molecular switches to turn on ATP-generating catabolic pathways while suppressing ATP-consuming anabolic pathways in energy-poor or other stress conditions (55). AMPK consists of a protein complex encoded by three subunits, a serine-threonine kinase catalytic subunit (α) and two regulatory subunits (β, γ). When AMPK signaling is disrupted, cells can eventually undergo cell death due to lack of signal reprogramming in starvation conditions. Signaling
networks initiated by AMPKs are implicated in a variety of human diseases including diabetes, lipid disorders, and cardiovascular disease (59).
!"#$%&'()*
!"#$%&'()*!!"#$%&'()*+!*,!(-#!./01'2()3'(#4!5)+'6#7!8+,9:!;+!-)$-!$&%2*6#!2*+4)1 ()*+67!8+,9!)6!)+'2()3#!<#2'%6#!(-#!='(#!*,!4#>-*6>-*=?&'()*+!<?!>-*6>-'('6#! @&2AB"#$9!#C2##46!(-#!='(#!*,!>-*6>-*=?&'()*+!<?!5)+'6#6!D&E97!8'597!'+4!F*6G:! ;+!&*H!$&%2*6#!2*+4)()*+67!(-#!='()*!*,!.I0J.F0!)+2=#'6#6!'+4!.I0!<)+46!(-#!8+,K! =#$%&'(*=?!6%<%+)(!(*!>=*(#2(!8+,9!,=*E!4#>-*6>-*=?&'()*+:!.2()3'()*+!*,!8+,9!&#'46! (*!'2()3'()*+!*=!4'E>#+)+$!*,!E#('<*&)2!>'(-H'?6!(*!*>()E)L#!#+#=$?!%6'$#:
!
@'&MGB8)>9B8)>N!
8+,9!
D&E9!
8'59!
F*6G!
"#$9!
@&2A!
8+,K!
0!
+,-'.+/0,12' 34.3'.+/0,12'
5)+'6#6!
>-*6>-'('6#!
activate Snf1 by phosphorylation (28). Notably, in support of yeast as a valuable model system, the identification of the Snf1-activating kinases in yeast (28, 56) quickly led to the discovery of the mammalian AMPK kinases (25, 30, 63).
The protein phosphatase complex Glc7/Reg1 mediates dephosphorylation of Snf1 (40, 44, 51). Glc7 is the catalytic subunit of the phosphatase complex whereas Reg1 is the regulatory subunit that targets Glc7 to substrates. Glc7 has numerous other substrates, which are targeted by a variety of regulatory subunits (19, 34, 36, 39). Yeast are inviable without Glc7, so the deletion of REG1 is used to uncouple Glc7 phosphatase activity towards Snf1. In the absence of Reg1, the Snf1 complex is constitutively active resulting in loss of glucose repression (29, 44, 58).
The phosphorylation (activation) status of the kinase Snf1 is a balance between upstream kinases/phosphatase activity and active site availability. In
energy-rich conditions, Snf1 exists in an unphosphorylated state because the rate of phosphate removal by Glc7/Reg1 exceeds the rate of phosphate addition by Elm1, Sak1, and Tos3 (49). In glucose stress conditions, Snf1 is phosphorylated because the regulatory Snf4 subunit binds to two molecules of ADP and protects Snf1 from dephosphorylation(42). Consequently, Snf1 persists in an activated state until ATP levels are restored. This mechanism of regulation is conserved in mammals (1).
An ongoing question in the field is why there exists three ‘redundant’
upstream kinases for Snf1. Indeed, deletion of all three kinases is needed to abolish Snf1 activity and result in a snf1Δ phenotype (28). There are some subtle
findings that Sak1 is the only upstream kinase to be found in a stable complex with Snf1 (18). In addition, cell lysates purified from strains expressing each upstream kinase individually showed Snf1 had the highest activity toward a peptide substrate in the strain containing only Sak1 (26, 28). While Sak1 and Tos3 have only been shown to function as Snf1 kinases, Elm1 has been shown to have other functions besides Snf1 metabolism regulation. Elm1 also has regulatory functions involving cell morphology, filamentous invasive growth, and control of bud growth and cytokinesis (6, 54). Studies have shown that the differences between the Elm1, Sak1, and Tos3 kinases are dependent on the type of carbon source stress and the different isoforms of the Snf1 kinase complex (43). It is likely that novel functions and substrates exist that will, if identified, further define the differences between these kinases.
Cross talk between signaling pathways
between pathways provides a means to tune and prioritize signaling responses. Cross talk can mediate responses to attenuate or strengthen two signaling pathways. For example, previous work by Nagiec et al. examined cross talk between the mating pathway and the hyperosmotic stress pathway in yeast (46). Cells were treated simultaneously with pheromone and salt, and monitored for pheromone response. Under these circumstances, Nagiec et al. observed an attenuation of the mating pathway, as determined by quantitative mating assays, microscopic
assessment of mating projection morphology, dampened mating transcriptional response and dampened MAPK activation. In this case the mating pathway and hyperosmotic stress pathway share signaling components. Thus, the mechanism by which cross talk dampens the mating pathway is complex and acts on multiple pathway components, where some are shared and some are not. Another example where one pathway antagonizes another is in the Protein Kinase A (PKA) and MAPK pathways in human arterial smooth muscle cells (23). Cells were treated with
communication between signaling cascades. In this age where patients are taking numerous drugs for multiple ailments, it is crucial to investigate cross talk between pharmaceutically-targeted signaling pathways.
Summary
The remainder of this thesis is divided into 3 chapters. In Chapter II, “Cell cycle-dependent phosphorylation and ubiquitination of a G protein α subunit,” I present a screen of a kinase gene deletion library to determine the Gpa1 kinase. Also in chapter II, I show how we determined that Gpa1 phosphorylation by Elm1 occurs in a cell cycle-dependent manner and leads to polyubiquitination and destruction of Gpa1. In Chapter III, “Regulation of a G protein by AMPK/Snf1 kinases,” I examine how Gpa1 is phosphorylated in response to glucose limitation and causes dampened GPCR-mediated signaling. Finally, in Chapter IV,
References
1. Adams J, et al (2004) Intrasteric control of AMPK via the gamma1 subunit AMP allosteric regulatory site. Protein Sci 13: 155-165.
2. Apanovitch DM, Slep KC, Sigler PB & Dohlman HG (1998) Sst2 is a GTPase-activating protein for Gpa1: Purification and characterization of a cognate RGS-galpha protein pair in yeast. Biochemistry 37: 4815-4822.
3. Bardwell L (2005) A walk-through of the yeast mating pheromone response pathway. Peptides 26: 339-350.
4. Bjarnadóttir TK, et al (2006) Comprehensive repertoire and phylogenetic analysis of the G protein-coupled receptors in human and mouse. Genomics 88: 263-273.
5. Blumer KJ & Thorner J (1990) Beta and gamma subunits of a yeast guanine nucleotide-binding protein are not essential for membrane association of the alpha subunit but are required for receptor coupling. Proc Natl Acad Sci USA 87: 4363-4367.
6. Bouquin N, et al (2000) Regulation of cytokinesis by the Elm1 protein kinase in Saccharomyces cerevisiae. J Cell Sci 113: 1435-1446.
7. Cappell SD, Baker R, Skowyra D & Dohlman HG (2010) Systematic analysis of essential genes reveals important regulators of G protein signaling. Mol Cell 38: 746-757.
8. Carlson M, Osmond BC & Botstein D (1981) Mutants of yeast defective in sucrose utilization. Genetics 98: 25-40.
9. Celenza JL & Carlson M (1989) Mutational analysis of the Saccharomyces cerevisiae SNF1 protein kinase and evidence for functional interaction with the SNF4 protein. Mol Cell Biol 9: 5034-5044.
10. Chu J, Zheng H, Zhang Y, Loh HH & Law PY (2010) Agonist-dependent mu-opioid receptor signaling can lead to heterologous desensitization. Cell Signal 22: 684-696.
11. Clark KL, Dignard D, Thomas DY & Whiteway M (1993) Interactions among the subunits of the G protein involved in Saccharomyces cerevisiae mating. Mol Cell Biol 13: 1-8.
13. Dohlman HG, Apaniesk D, Chen Y, Song J & and Nusskern D (1995) Inhibition of G-protein signaling by dominant gain-of-function mutation in Sst2p, a
pheromone desensitization factor in Saccharomyces cerevisiae. Mol Cell Biol 15: 3635-3643.
14. Dohlman HG & Thorner JW (2001) Regulation of G protein-initiated signal transduction in yeast: Paradigms and principles. Annu Rev Biochem 70: 703-754.
15. Drews J (2000) Drug discovery: A historical perspective. Science 287: 1960-1964.
16. Duncan JA & Gilman AG (2002) Characterization of Saccharomyces cerevisiae acyl-protein thioesterase 1, the enzyme responsible for G protein alpha subunit deacylation in vivo. J Biol Chem 277: 31740-31752.
17. Duronio RJ, Towler DA, Heuckeroth RO & Gordon JI (1989) Disruption of the yeast N-myristoyl transferase gene causes recessive lethality. Science 243: 796-800.
18. Elbing K, McCartney RR & Schmidt MC (2006) Purification and characterization of the three Snf1-activating kinases of Saccharomyces cerevisiae. Biochem J 393: 797-805.
19. Frederick DL & Tatchell K (1996) The REG2 gene of Saccharomyces
cerevisiae encodes a type 1 protein phosphatase-binding protein that functions with Reg1p and the Snf1 protein kinase to regulate growth. Mol Cell Biol 16: 2922-2931.
20. Giaever G, et al (2002) Functional profiling of the Saccharomyces cerevisiae genome. Nature 418: 387-391.
21. Glick JL, Meigs TE, Miron A & Casey PJ (1998) RGSZ1, a gz-selective
regulator of G protein signaling whose action is sensitive to the phosphorylation state of gzalpha. J Biol Chem 273: 26008-26013.
22. Goffeau A, et al (1996) Life with 6000 genes. Science 274: 563-567.
23. Graves LM, et al (1993) Protein kinase A antagonizes platelet-derived growth factor-induced signaling by mitogen-activated protein kinase in human arterial smooth muscle cells. Proc Natl Acad Sci USA 90: 10300-10304.
25. Hawley SA, et al (2005) Calmodulin-dependent protein kinase kinase-beta is an alternative upstream kinase for AMP-activated protein kinase. Cell Metab 2: 9-19.
26. Hedbacker K, Hong SP & Carlson M (2004) Pak1 protein kinase regulates activation and nuclear localization of Snf1-Gal83 protein kinase. Mol Cell Biol 24: 8255-8263.
27. Hildebrandt JD (1997) Role of subunit diversity in signaling by heterotrimeric G proteins. Biochem Pharmacol 54: 325-339.
28. Hong SP, Leiper FC, Woods A, Carling D & Carlson M (2003) Activation of yeast Snf1 and mammalian AMP-activated protein kinase by upstream kinases. Proc Natl Acad Sci USA 100: 8839-8843.
29. Huang D, Chun KT, Goebl MG & Roach PJ (1996) Genetic interactions
between REG1/HEX2 and GLC7, the gene encoding the protein phosphatase type 1 catalytic subunit in Saccharomyces cerevisiae. Genetics 143: 119-127. 30. Hurley RL, et al (2005) The Ca2+/calmodulin-dependent protein kinase kinases
are AMP-activated protein kinase kinases. J Biol Chem 280: 29060-29066. 31. Jiang R & Carlson M (1997) The Snf1 protein kinase and its activating subunit,
Snf4, interact with distinct domains of the Sip1/Sip2/Gal83 component in the kinase complex. Mol Cell Biol 17: 2099-2106.
32. Kao LR, Peterson J, Ji R, Bender L & Bender A (1996) Interactions between the ankyrin repeat-containing protein Akr1p and the pheromone response pathway in Saccharomyces cerevisiae. Mol Cell Biol 16: 168-178.
33. Kennedy BK, Austriaco NR, Jr., Zhang J & Guarente L (1995) Mutation in the silencing gene SIR4 can delay aging in S. cerevisiae. Cell 80: 485-496. 34. Knaus M, et al (2005) The Bud14p-Glc7p complex functions as a cortical
regulator of dynein in budding yeast. EMBO J 24: 3000-3011.
35. Kozasa T & Gilman AG (1996) Protein kinase C phosphorylates G alpha 12 and inhibits its interaction with G beta gamma. J Biol Chem 271: 12562-12567. 36. Kozubowski L, et al (2003) A Bni4-Glc7 phosphatase complex that recruits
chitin synthase to the site of bud emergence. Mol Biol Cell 14: 26-39.
38. Lesage P, Yang X & Carlson M (1996) Yeast SNF1 protein kinase interacts with SIP4, a C6 zinc cluster transcriptional activator: A new role for SNF1 in the glucose response. Mol Cell Biol 16: 1921-1928.
39. Lin JT & Lis JT (1999) Glycogen synthase phosphatase interacts with heat shock factor to activate CUP1 gene transcription in Saccharomyces cerevisiae. Mol Cell Biol 19: 3237-3245.
40. Ludin K, Jiang R & Carlson M (1998) Glucose-regulated interaction of a regulatory subunit of protein phosphatase 1 with the Snf1 protein kinase in Saccharomyces cerevisiae. Proc Natl Acad Sci USA 95: 6245-6250.
41. Manganello JM, Huang JS, Kozasa T, Voyno-Yasenetskaya TA & Le Breton GC (2003) Protein kinase A-mediated phosphorylation of the G alpha 13 switch region alters the G alpha/beta/gamma-GPCR complex and inhibits rho
activation. J Biol Chem 278: 124-130.
42. Mayer FV, et al (2011) ADP regulates SNF1, the Saccharomyces cerevisiae homolog of AMP-activated protein kinase. Cell Metab 14: 707-714.
43. McCartney RR, Rubenstein EM & Schmidt MC (2005) Snf1 kinase complexes with different beta subunits display stress-dependent preferences for the three Snf1-activating kinases. Curr Genet 47: 335-344.
44. McCartney RR & Schmidt MC (2001) Regulation of the Snf1 kinase. Activation requires phosphorylation of threonine 210 by an upstream kinase as well as a distinct step mediated by the Snf4 subunit. J Biol Chem 276: 36460-36466. 45. McCudden CR, Hains MD, Kimple RJ, Siderovski DP & Willard FS (2005)
G-protein signaling: Back to the future. Cell Mol Life Sci 62: 551-577. 46. Nagiec MJ & Dohlman HG (2012) Checkpoints in a yeast differentiation
pathway coordinate signaling during hyperosmotic stress<br />. PLoS Genet 8: e1002437.
47. Navarro L, et al (2007) Identification of a molecular target for the yersinia protein kinase A. Mol Cell 26: 465-477.
48. Novick P, Field C & Schekman R (1980) Identification of 23 complementation groups required for post-translational events in the yeast secretory pathway. Cell 21: 205-215.
49. Rubenstein EM, et al (2008) Access denied: Snf1 activation loop
50. Russ AP & Lampel S (2005) The druggable genome: An update. Drug Discov Today 10: 1607-1610.
51. Sanz P, Alms GR, Haystead TA & Carlson M (2000) Regulatory interactions between the Reg1-Glc7 protein phosphatase and Snf1 protein kinase. Mol Cell Biol 20: 1321-1328.
52. Shi J, Zemaitaitis B & Muma NA (2007) Phosphorylation of a G alpha 11 protein contributes to agonist induced densensitization of 5-HT 2A receptor signaling. Mol Pharmaco 71: 303-313.
53. Sprang SR (1997) G protein mechanisms: Insights from structural analysis. Annu Rev Biochem 66: 639-678.
54. Sreenivasan A & Kellogg D (1999) The elm1 kinase functions in a mitotic signaling network in budding yeast. Mol Cell Biol 19: 7983-7994.
55. Steinberg GR & Kemp BE (2009) AMPK in health and disease. Physiol Rev 89: 1025-1078.
56. Sutherland CM, et al (2003) Elm1p is one of the three upstream kinases for the Saccharomyces cerevisiae SNF1 complex. Curr Biol 13: 1299-1305.
57. Torres MP, et al (2009) G protein mono-ubiquitination by the Rsp5 ubiquitin ligase. J Biol Chem 284: 8940-8950.
58. Tu J & Carlson M (1995) REG1 binds to protein phosphatase type 1 and regulates glucose repression in Saccharomyces cerevisiae. EMBO J 14: 5939-5946.
59. Viollet B & Andreelli F (2011) AMP-activated protein kinase and metabolic control. Handb Exp Pharmacol 203: 303-330.
60. Wilson WA, Hawley SA & Hardie DG (1996) Glucose repression/derepression in budding yeast: SNF1 protein kinase is activated by phosphorylation under derepressing conditions, and this correlates with a high AMP:ATP ratio. Curr Biol 6: 1426-1434.
61. Winzeler EA, et al (1999) Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science 285: 901-906. 62. Wise A, Gearing K & Rees S (2002) Target validation of G-protein coupled
receptors. Drug Discov Today 7: 235-246.
64. Woods A, et al (1994) Yeast SNF1 is functionally related to mammalian AMP-activated protein kinase and regulates acetyl-CoA carboxylase in vivo. J Biol Chem 269: 19509-19515.
65. Yang X, Jiang R & Carlson M (1994) A family of proteins containing a
CHAPTER II
CELL CYCLE-DEPENDENT PHOSPHORYLATION AND UBIQUITINATION OF A G
PROTEIN α SUBUNIT*Φ
A diverse array of external stimuli, including most hormones and
neurotransmitters, bind to cell surface receptors that activate G proteins. Mating
pheromones in yeast Saccharomyces cerevisiae activate G protein-coupled
receptors and initiate events leading to cell cycle arrest in G1 phase. Here, we show
that the Gα subunit (Gpa1) is phosphorylated and ubiquitinated in response to
changes in the cell cycle. We systematically screened 109 gene deletion strains
representing the non-essential yeast kinome and identified a single kinase gene,
ELM1, as necessary and sufficient for Gpa1 phosphorylation. Elm1 is expressed in a
cell cycle-dependent manner, primarily at S and G2/M. Accordingly, phosphorylation
of Gpa1 in G2/M phase leads to polyubiquitination in G1 phase. These findings
demonstrate that Gpa1 is dynamically regulated. More broadly, they reveal how G
proteins can simultaneously regulate, and become regulated by, progression
through the cell cycle.
* Elements of the work referenced in this chapter have been published in: Torres MP, Clement ST,
Cappell SD, and Dohlman HG. (2011) Cell cycle-dependent phosphorylation and ubiquitination of a G protein α subunit. J Biol Chem. 286, 20208-16.
Φ I am including this work in my thesis as I am a co-author on this publication. I performed the initial kinome screen, which lead to the examination of Elm1 as a potential kinase for Gpa1. I also
Introduction
G protein coupled receptors and heterotrimeric G proteins are the
predominant components through which cells receive and transduce extracellular
signals. G protein signal transduction is highly conserved throughout eukaryotes,
including the yeast Saccharomyces cerevisiae. In yeast, haploid a-type cells secrete
an a-factor pheromone that binds to receptors on the surface of α-type cells,
whereas α-type cells secrete an α-factor that acts exclusively on a-type cells (13).
Consequently, the haploid cells fuse to form an a/α diploid cell.
As with other G protein systems, activation of the yeast pheromone receptor
stimulates exchange of GDP for GTP on the G protein α subunit (Gpa1), which
promotes its dissociation from the Gβγ (Ste4/18) heterodimer (13, 44). In yeast, Gβγ
is primarily responsible for transmission and amplification of the signal to effector
proteins, whereas the Gα serves primarily to regulate the levels of free Gβγ. As a
consequence, cells are highly sensitive to small changes in the stoichiometry of Gα
and Gβγ (5, 7, 43). The signal is terminated by hydrolysis of GTP to GDP on the Gα
subunit, which promotes reassociation of the heterotrimeric G protein complex.
Further regulation is imposed by accelerating the GTPase activity of Gα via
regulators of G protein signaling proteins (21).
Propagation of the G protein signal requires components of a MAPK cascade
(Ste20, Ste11, Ste7, Fus3 or Kss1), a MAPK scaffold (Ste5), as well as a
transcription factor (Ste12) (13). Consequently, the pheromone initiates changes in
gene expression and cell morphology that prepare the cell to undergo cell-cell and
which ensures that each haploid cell contains one copy of every chromosome before
fusion (61). Pheromone-induced G1 arrest necessarily prevents fusion during DNA
replication (S phase) or before mitosis (M phase), during which nuclear fusion could
lead to missegregation of genetic material, aneuploidy, and other proliferative
disadvantages (51, 52). Consequently, yeast mating and cell cycle progression must
be highly coordinated processes.
The mating pathway is well known to regulate the cell cycle by stimulating the
expression of Far1, a cyclin-dependent kinase inhibitor that arrests cells at START in
late G1 phase (34, 36, 38, 39, 55). Reciprocally, Far1 is itself regulated in a cell-cycle
dependent manner and is degraded after cells pass through START and exit G1
phase (34, 38). Degradation of Far1 is initiated in G1 by cyclin-dependent
kinase-mediated phosphorylation, which promotes its ubiquitination by the SCF
(Skp1/Cullin/F-box) ubiquitin ligase (3, 20, 36).
Emerging evidence suggests that multiple mating pathway components are
regulated by ubiquitination. Targets of ubiquitin-mediated degradation include the
regulator of G protein signaling protein Sst2 (18), the MAPK kinase Ste7 (56), the
MAPK scaffold protein Ste5 (16), and the G protein α subunit Gpa1 (31, 32, 54, 57).
Of these, the SCF has been shown to be necessary and sufficient for ubiquitination
of Gpa1 (5). While SCF substrates are generally phosphorylated prior to
ubiquitination (10, 60), as yet the kinase that phosphorylates Gpa1 has not been
identified.
Here we identify a novel G protein kinase, Elm1. We show that the G protein
with Elm1 expression during the cell cycle. G protein phosphorylation in G2/M phase
leads to ubiquitination during the following G1 phase. In the absence of Elm1, the G
protein is neither phosphorylated nor ubiquitinated. Taken together, these findings
show how G proteins can simultaneously regulate, and become regulated by,
progression through the cell cycle.
Results
Gpa1 Is A Phosphoprotein -Gpa1 undergoes a variety of post-translational
modifications including myristoylation, palmitoylation, and ubiquitination (32, 58).
The myristoylation state of Gpa1 can be distinguished by an electrophoretic mobility
shift following SDS-PAGE and immunoblotting. Because only a fraction of Gpa1 has
shifted mobility under these conditions, the prevailing view has been that Gpa1
exists in both myristoylated and non-myristoylated states (46). However, upon close
inspection of the overexpressed protein, we found that even non-myristoylated
Gpa1G2A exhibits differential mobility shift by immunoblotting, indicating the presence
of another modification (Fig. 2.1 A). Given that phosphorylation can likewise alter the
electrophoretic mobility of proteins, we asked whether phosphorylation rather than
myristoylation might account for the second form of Gpa1. Consistent with this
hypothesis, phosphatase treatment of whole cell extracts (Fig. 2.1 B) or of purified
Gpa1 (Fig. 2.1 C) resulted in a complete loss of the slower migrating form of Gpa1.
!"#$%&'()*
!"#$%&'()*''!"#$%"#&'()*+#,-+,./01$-),-1(10*&#%"#&1*+0-2#3+(+*'-$"+4*-+,-5%)67'
822/,3(#*-),)('$+$-#4-5%)6-/$+,9-5%)6:$%10+4+0-),*+3#.+1$7-;!<-=1(($-#>1&1?%&1$$:
+,9-@+(.:*'%1-5%)6-;%ABCD:5%)6<-;"#<-#&-*"1-2'&+$*#'()*+#,:$+*1-2/*),*-5%)65EA
-;$%!
<-),)('F1.-3'-+22/,#3(#**+,97-G"1-)&&#@-2)&H$-*"1-%#$+*+#,-#4-%&1$/2%*+>1-%"#$%"#&'()*1.-4#&2-#4-5%)67-;&<-I#(/3(1-%&#*1+,-1?*&)0*$-4-01(($-#>1&1?%&1$$:
+,9-@+(.:*'%1-5%)6-@1&1-$%(+*-+,*#-*@#-$1%)&)*1-)(+J/#*$K-),.-#,1-")(4-@)$-*&1)*1.-;!<-@+*"-":%&#*1+,-%"#$%")*)$1-;''()*<7-'+$'(,K-%"#$%"#&'()*1.-5%)67-;-<-5%)6:
LMA5-%/&+4+1.-4-,#0#.)F#(1:)&&1$*1.-01(($-),.-*&1)*1.-;N<-@+*"-":%&#*1+,-%"#$:
%")*)$1-;''()*<7-;.<-O+(.:*'%1-01(($-#>1&1?%&1$$+,9-@+(.:*'%1-5%)6-;"#<-#&-%"#$:
%"#&'()*+#,-$+*1-2/*),*$-;IEPPAK-!/!-#&-IEPPQK-$/0<7
phosphorylation site by mass spectrometry (28), altered the mobility of Gpa1.
Substitution of Ser-200 with alanine (S200A) or glutamate (S200E) replicated the
mobility of dephosphorylated and phosphorylated Gpa1, respectively (Fig. 2.1 D).
We conclude that Gpa1 is a phosphorylated protein.
A Yeast Kinome Screen Reveals Elm1 as a G Protein Kinase - To identify the
Gpa1 kinase, we monitored the phosphorylation-dependent mobility shift of
endogenously expressed Gpa1 in gene deletion strains representing the majority of
the yeast kinome. Of the 109 strains tested, deletion of ELM1 alone resulted in a
significant observable loss of phosphorylated Gpa1 (Fig. 2.2). Phosphorylation of
Gpa1 was restored by plasmid-borne expression of ELM1 (Fig. 2.3 A). We observed
no such differences in the absence of kinases that act downstream of Elm1 or that
are functionally similar to Elm1 (see discussion, Fig. 2.2) (1, 45). We conclude that
Elm1 phosphorylates Gpa1 in vivo.
Elm1 Is Required for Maximal Pheromone-induced Gene Transcription - Elm1
is best known as a regulator of cell morphology during G2/M phase of the cell cycle.
During G2/M, Elm1 phosphorylates proteins required for the morphogenesis
checkpoint that coordinates bud emergence and mitosis (8, 49) as well as
organization of septins during cytokinesis (1, 4, 45). Yeast harboring elm1 mutations
exhibit morphologically distinct growth pattern in which cells delay cytokinesis and
undergo elongated bud growth, a process that also occurs under conditions or
nitrogen starvation and filamentous growth (26). Elm1 has been proposed to inhibit
the filamentous growth response that includes multiple signaling branches, including
!"
#$%&'$#
!"#$
!
%&!'
!
!()$
!
)*+$
!
,*"-
!
#./-
!
0!*$
!
1./-
!
!/2$
!
"#
!"#$%&
'
#
!"#$%&'()(
well in the pheromone response pathway. Accordingly, we found that Kss1 was
more highly expressed and activated in elm1Δ cells compared to wild-type cells (Fig.
2.3 B). In addition, deletion of KSS1 in elm1Δ cells reduced filamentous-like
phenotype (elongated buds and flocculation) typical of elm1Δ cells (Fig. 2.3 C). In
contrast, the other pheromone-responsive MAP kinase Fus3 was largely unaffected
by the absence of Elm1 (Fig. 2.3 B).
We next determined whether Elm1 regulates pheromone-dependent gene
transcription using a β-galactosidase reporter fused with the promoter of the
mating-specific gene FUS1. We found that elm1Δ cells exhibit a significantly reduced
maximum level of pheromone-induced gene transcription. Signaling was restored
upon plasmid-borne expression of wild-type Elm1 but not kinase-inactive Elm1K177R
(Fig. 2.3 D). Thus, Elm1 represses activation of the MAP kinase branch of the
filamentous growth response, including Kss1, and is required for maximal response
to the mating pheromone.
Gpa1 is phosphorylated directly by Elm1 - Elm1 clearly plays a role in multiple
signaling pathways, including the filamentous growth and pheromone response
pathways, any of which could be indirectly responsible for the diminished Gpa1
phosphorylation in elm1Δ cells. Therefore, we asked whether Elm1 acts directly on
Gpa1. We established that purified Elm1 could bind to and phosphorylate
recombinant Gpa1 in vitro (Fig. 2.4). Elm1 is also capable of autophosphorylation
(26), and we show that this activity is exhibited in GST-Elm1 (Fig. 2.4 A-C). Both
Elm1 autophosphorylation and Gpa1 transphosphorylation were blocked when
!"#$%&'()* !
"
#
$$"%&&' $$"()&*+, !"#$∆
!"#$∆
%&&$∆
" # " # " #
α"(
%&&'
()&*
!"#$∆
'!($∆ %&&$∆
" #
$$"%&&' $$"()&* -!./ $00102 -!./%&&' -!./()&* 34 +, !"#$∆
" # " # " # " #
)*+$,-..+
5$6'
α"(
"
" "
"
"
" # #
/01$ $"5$6' !"#$∆ #/ /01$ !"#$∆ #/ !"#$2$$34 +, !"#$∆ %&&$∆ %&&$∆ !"#$∆ 78 '8"9 '8"; '8": '8"< '8"= '8"0
8 = '8 '= 28 +,/#/>? !"#$∆/#/>? !"#$∆/#//01$ !"#$∆/#/!"#$2$$34 β "@6A /4BCDEDCF/GH(IJ 7-3 K'888
L!"(MN/O
!"#$%&'()*//PQ6&C/RDSTUQ/&BVQQS/VQEQ6A&/>AU'/6&/6/5$6'/RDS6&QW/,XQ/6Y)SZ6SBQ/ T[/5$6'/\6&/ZQCQVUDSQZ/YF/DUU)STYATCCDS@/'89/ZD[[QVQSC/RDS6&Q/@QSQ/ZQAQCDTS/ &CV6DS&W/4/&DS@AQ/RDS6&QN/>AU'N/D&/VQ])DVQZ/[TV/ZQCQBCDTS/T[/$XT&$XTVFA6CQZ/5$6'W/ G+J/?6ADZ6CDTS/T[/RDSTUQ/&BVQQS/VQ&)AC&W/+DAZ"CF$Q/BQAA&/TV/BQAA&/QK$VQ&&DS@/ 5$6'^2884/GYF/@QSQ/VQ$A6BQUQSCJ/TV/!"#$"/BQAA&/X6VYTVDS@/QU$CF/EQBCTV/TV/
$A6&UDZ"YTVSQ//01$W/G5J/4S6AF&D&/T[/$XQVTUTSQ"ZQ$QSZQSC/G#J/O4_/RDS6&Q/6BCD" E6CDTS/G'67N/ZD[[QVQSC/QK$T&)VQ&J/6SZ/CTC6A/QK$VQ&&DTS/G86''6#J/DS/+,N/!"#$"N/6SZ/ !"#$"%&&$"/BQAA&W/95N/DUU)STYATCW/G:J/7D[[QVQSCD6A/DSCQV[QVQSBQ/BTSCV6&C/G;9:J/ DU6@Q&/T[/FQ6&C/&CV6DS&/)&QZ/DS/CXQ/$XQVTUTSQ/&D@S6ADS@/6&&6F&/&XT\S/DS/G5J/6SZ/ G;JW/G;J/4S6AF&D&/T[/$XQVTUTSQ"ZQ$QSZQSC/@QSQ/CV6S&BVD$CDTS/DS/!"#$"/BQAA&/\DCX/ CXQ/DSZDB6CQZ/$A6&UDZ/BTSC6DSDS@//01$N/!"#$2$$34N/TV/6S/QU$CF/EQBCTVW/-SZDB6CQZ/6VQ/
FQ6&C/&CV6DS&/QK$VQ&&DS@/6/<=,$>0?(@/VQ$TVCQV/CVQ6CQZ/\CDX/CXQ/DSZDB6CQZ/BTSBQS" CV6CDTS&/T[/U6CDS@/$XQVTUTSQW/HQ&)AC&/&XT\/CXQ/UQ6S/`/^>O/GSa0JN/Q6BX/
A). Consistent with our findings in vivo, Ser-200 is required for maximum
phosphorylation of Gpa1 (Fig. 2.4 B). Similar results were observed upon mutation
of all 15 Ser and Thr residues in the ubiquitination domain (data not shown). Finally,
Elm1 was able to bind to, but not effectively phosphorylate, Gpa1ΔUD, a mutant that
lacks the ubiquitination domain of the protein (residues 129-236) (Fig. 2.4 C and D).
We conclude that Elm1 phosphorylates Gpa1 directly at serine 200 and other
multiple other sites throughout the protein.
Gpa1 Phosphorylation is Cell Cycle-dependent – We next determined how
phosphorylation of Gpa1 is regulated. Elm1 is expressed primarily during S and
G2/M phases of the cell cycle (4, 26, 37). Further, longstanding evidence indicates
that the mobility-shifted (phosphorylated) form of Gpa1 is significantly reduced upon
pheromone stimulation (12), which induces cell cycle arrest in G1 phase. Thus, we
considered whether Gpa1 phosphorylation is regulated during the cell cycle. We
directly compared Gpa1 from cells arrested in G1 phase with α-factor (α-F), in S
phase with hydroxyurea (HU), or in G2/M phase with nocodazole (Nz). Hydroxyurea
inhibits deoxyribonucleotide synthesis, inducing a DNA replication checkpoint arrest
in S phase (59). Nocodazole inhibits microtubule polymerization and induces a
checkpoint arrest at the metaphase/anaphase transition of mitosis (25). We found
that phosphorylated Gpa1 is most abundant in cells arrested in S and G2/M, but
lowest in cells arrested in G1 phase (Fig. 2.5 A). To determine the dynamics of
phosphorylation during the cell cycle, we monitored Gpa1 during arrest and release
in cells treated with nocodazole or hydroxyurea. To validate the arrest and release
! " # !"#$%&'()* $%&'()*+, )'-(./0, 1 )'-(./0,2,,34 $%&'()*+,
( 1 1 1
( ( ( ( ( (( 5- 2,,34 )'-(./0, &"67)'-&8*9: ;<=(>8?@A*@A+:>@8 )'-(./0,
5-!"#$%&'()*'./0,7>B78C?CBB+AD7+8E7B9FF>?>C8:7F@A7)*+,7*G@B*G@AD/+:>@8H7I!J7"#$%K7&'(
)&%*+7L>8+BC7+BB+D7M>:G7*9A>F>CE7)'-(./0,7@A7?+:+/D:>?+//D7>8+?:>NC7./0,2,3347+8E7
*9A>F>CE7AC?@0O>8+8:7)*+,7>87:GC7*ACBC8?C7@F7P!;<=Q#-=H7,&-.%/7)'-(./0,7>8*9:7
EC:C?:CE7OD7>0098@O/@:7M>:G7+8:>()'-H7I0J71'()&%*+7L>8+BC7+BB+D7M>:G7)'-(./0,7
+8E7*9A>F>CE7AC?@0O>8+8:7)*+,7I23JK7)*+,'<RR#7I4566!J7@A7)*+,",(;$K",<S(<;$7I"78J7
/+?L>8T7:GC79O>U9>:>8+:CE7B9OE@0+>8H7'G@M87>B7:GC(>8?@A*@A+:>@87@F7A+E>@+?:>NC7 *G@B*G+:C7+8E7)*+,7>8*9:7EC:C?:CE7OD7!@@0+BB>C7B:+>8>8TH7I9J7"#$%K7&'()&%*+7 L>8+BC7+BB+D7M>:G7)'-(./0,7+8E7*9A>F>CE7AC?@0O>8+8:7)*+,7I23JK7)*+,",(;$7I":J7
@A7)*+,",(;$K",<S(<;$7I"78J7/+?L>8T7:GC79O>U9>:>8+:CE7B9OE@0+>8H7'G@M87>B7:GC(>8?@A(
*@A+:>@87@A7A+E>@+?:>NC7*G@B*G+:C7+8E7)*+,7>8*9:7EC:C?:CE7OD7!@@0+BB>C7B:+>8( >8TH7I8J71'()&%*+7+FF>8>:D7*9A>F>?+:>@879B>8T78>?LC/7+FF>8>:D70+:A>V7+8E7>0098@EC:C?:>@87 @F7O@98E7)'-(./0,7:@7*9A>F>CE7AC?@0O>8+8:7)*+,K7)*+,",(;$7I":J7@A7)*+,",(;$K",<S( <;$7I"78JH7
)'-(./0, )'-(./0, $%&'()*+,
(
&"67)*+, &"67)'-&=67$%&' >8*9: $%&'()*+, ∆W ∆XY1 1 1
I,RRZJ
'
)'-(./0,
$%&'()*+,
5- ∆XY ( $%&'()*+,
)'-(./0, 1 1 1
;<=(>8?@A*@A+:>@8 &8*9:
5-(
∆W
1
5-∆W∆XY
phosphorylated Gpa1 accumulates in a coordinated fashion with Clb2 as cells arrest
in either G2 /M or S phase (Fig. 2.5, B and D). Upon release from nocodazole,
phosphorylated Gpa1 and Clb2 rapidly decrease in abundance and then increase
concomitantly as cells reenter the cell cycle (Fig. 2.5 C). Similar coordination
between phosphorylated Gpa1 and Clb2 was evident in cells released from
hydroxyurea induced cell cycle arrest (Fig. 2.5 E). Finally, we observed
temperature-dependent accumulation of phosphorylated Gpa1 in cdc6–1ts cells, which are
incapable of DNA replication licensing, resulting in a post-START arrest in late
G1/early S phase (Fig. 2.5 A) (29, 30). We conclude that Gpa1 phosphorylation is
Elm1-dependent and cell cycle-regulated. Although phosphorylated Gpa1
accumulates throughout the S and G2/M phases, it is rapidly eliminated from cells
during G1 phase.
Elm1 Is Required for Gpa1 Polyubiquitination - We have shown previously
that Gpa1 is polyubiquitinated by the SCFCdc4 ubiquitin ligase (5) and that
ubiquitination occurs primarily at lysine 165 within the ubiquitinated subdomain of
Gpa1 (32). Typically, SCF recruits phosphorylated proteins as substrates for
ubiquitination (10, 60). Therefore, we asked whether Elm1 phosphorylation promotes
Gpa1 polyubiquitination. We compared Gpa1 polyubiquitination in the presence and
absence of ELM1 using a proteasome-deficient yeast strain, cim3–1 (54, 57). For
these experiments, Gpa1 was overexpressed to allow detection of the minor
ubiquitinated species. Growth of cim3–1 cells at the restrictive temperature
inactivates Cim3, an essential protein component of the 26 S proteasome (15),
!"#$%&'()*
!"#$%&'()*'!"#$%"&'("&')*+#,-'.%-(%/0++%/*/+0120"0.20.,3%40++(%,)0#,02%5')%637%&%
8-,&%!15#/,')%"&0)'9'.0%:!!";%#))0(,%/0++(%-.%!$%"&#(0<;%&*2)'=*>)0#%:#$;%#))0(,(%
/0++(%-.%?%"&#(0<%')%.'/'2#@'+0%:%&;%#))0(,(%/0++(%-.%!6AB%"&#(0<%5'++'802%C*%
)0+0#(0%-.,'%5)0(&%902->9%8&0)0%-.2-/#,023%?&'8.%#)0%#+-D>',(%,#E0.%#,%-.2-/#,02% ,-90%"'-.,(%5'++'802%C*%-99>.'C+',,-.F3%G&0%#C>.2#./0%'5%"&'("&')*+#,02%!"#$%
:'()*+,)--./01)2<%#.2%4+C6%80)0%D>#.,-5-02%C*%20.(-,'90,)*%5)'9%,&0%-99>.'C+',%
(&'8.%#C'H03%3$;%#)C-,)#)*%>.-,(I%34;%#(*./&)'.'>(%/0++(I%5.)2;%!JKLM%+'#2-.F%
/'.,)'+3%:3<%L-)0/,%/'9"#)-('.%'5%!"#$%5)'9%#))0(,02%/0++(3%40++(%80)0%#+('%
#))0(,02%-.%+#,0%!$A0#)+*%?%"&#(0%C*%#%,09"0)#,>)0%(&-5,%'5%*2*6!7,/0++(3%:8<%
.'/'2#@'+0%#))0(,3%:4<%.'/'2#@'+0%)0+0#(03%:9<%&*2)'=*>)0#%#))0(,3%::<%&*2)'=*1
immunoblotting. Consistent with the hypothesis, Gpa1 polyubiquitination is
considerably diminished in the absence of Elm1 or in the presence of plasmid-borne
Elm1K117R (Fig. 2.6). To validate the observations made in cim3–1 cells, we also
monitored Gpa1 polyubiquitination in wild type or elm1Δ cells expressing
myc-ubiquitin (proteins conjugated to myc-myc-ubiquitin are degraded slowly) (14, 22). Once
again Gpa1 polyubiquitination was diminished in the absence of Elm1. We conclude
that Elm1 is required for ubiquitination, as well as for phosphorylation, of Gpa1.
Gpa1 Ubiquitination Is Regulated during the Cell Cycle - Phosphorylation is a
well-established precursor to ubiquitination by the SCF (60) and serves as a signal
for recruitment of target substrates by F-box proteins (42). Our previous findings
indicate that Gpa1 is ubiquitinated by SCFCdc4. The data presented above indicate
that Gpa1 is phosphorylated by Elm1 and that Elm1 is required for ubiquitination.
Therefore, we postulated that phosphorylation by Elm1 precedes ubiquitination by
SCF. To test this hypothesis, we compared the relative proportion of phosphorylated
Gpa1 in cells lacking functional Cdc4 (F-box protein) or Cdc34 (ubiquitin-conjugating
enzyme), which we previously identified as responsible for polyubiquitination of
Gpa1 (5). Accordingly, we monitored Gpa1 in temperature-sensitive cdc4-1 and
cdc34-2 mutants (27). Consistent with the hypothesis, we found the proportion of
phosphorylated Gpa1 to be higher in asynchronous cells lacking active forms of
either Cdc4 or Cdc34 but comparatively lower in identically treated wild-type cells
(Fig. 2.7 A, left panel). Similar results were obtained using Tet-repressible versions
of CDC4 and CDC34 (data not shown). Thus, phosphorylated Gpa1 accumulates in
!"#$%&'()*
!"#$%&'()*''!"#$%&'%()*+&(),%-.(%/01$%0."2+3&*+&4&514&.56%7)#0)(14+()8')5'&4&9)%% 0(.4)1'.#)%,)-&:&)54%;!"#$%&%.(%!"#$%&<'(#&!=%.(%&'.>)5&:%?&",8420)%:)""'%.9)()@8 0()''&5>%4A)%&5,&:14),%-.(#%.-%0"1'#&,83.(5)%)*+&%;0BCDE8/FB$G%)*+&=%>(.?5% 14%4A)%()'4(&:4&9)%4)#0)(14+()6%;+=%B::+#+"14&.5%.-%/01$%0."2+3&*+&4&514&.5%&5%
ubiquitination, in addition to phosphorylation, might be regulated by the cell cycle. In
support of this model, accumulation of phosphorylated Gpa1 in SCF-deficient cells is
comparable with that observed in G2/M arrested cells (Fig. 2.7 A, right panel).
Moreover, we found that Gpa1 polyubiquitination is significantly higher in cells
arrested in G1 phase withα-factor mating pheromone or by temperature inactivation
of cdc28-1 when compared with cells arrested in S phase or in G2/M phase (Fig. 2.7,
B and C). Although treatment with α-factor results in Far1-mediated G1 arrest, cells
expressing cdc28-1 undergo G1 arrest independent of pheromone pathway
activation at the restrictive temperature (35). In either case, Gpa1 polyubiquitination
is highest in cells arrested in G1 phase. Taken together, the data presented here
reveal that Gpa1 is dynamically regulated. Phosphorylation and ubiquitination are
independent of the pheromone stimulus and therefore not the result of feedback
regulation. Rather, these modifications occur in conjunction with the cell cycle. Thus,
the G protein is simultaneously a regulator of, and regulated by, cell cycle
progression.
Discussion
It is now well established that the G protein subunit Gpa1 is ubiquitinated.
Here we have begun to discern how this ubiquitination event is regulated. Our
investigation began by showing that Gpa1 is phosphorylated and that
phosphorylation results in an electrophoretic mobility shift of the protein. This
property allowed us to determine that a single kinase (Elm1) is necessary for proper
!"#$%&'()*!!"#$%!#&'()*+,)+-+.$-+&.!*(!/010234!+5!36''!3(3'67869)'$-62:!"#$%!#;&57
#;&8('$-+&.!+.!/01!<)-$.-!5-8$+.5!&8!"#$%!#&'()*+,)+-+.$+-+&.!).268!2+==686.-!36''! 3(3'6!$8865-62!3&.2+-+&.5:!>!?!@33)<)'$-+&.!&=!#;&5#;&8('$-62!"#$%!>"#$"%&?!+.! 36''5!'$3A+.9!=).3-+&.$'!/010234:!"#$%!+<<).&*'&-!$.$'(5+5!&=!B;&'6!36''!6C-8$3-5!
=8&<!-6<#68$-)86756.5+-+D6!'(')#&!&8!'('*)#+!<)-$.-!5-8$+.5E!&8!-;6!+5&96.+3!B+'27 -(#6!5-8$+.:!F;6!,%-./-%"0!86#8656.-5!-;6!8$-+&!&=!#;&5#;&8('$-62!"#$%!>12".,%3(?! -&!-;6!5)<!-&-$'!&=!"#$%!>12".%3(.,21124.,%3(5?!6C#865562!$5!$!#6836.-$96!>6781.
"%376?:!/;&B.!+5!-;6!5$<6!$.$'(5+5!3&.2)3-62!=&8!"#$%!=8&<!$5(.3;8&.&)5!>!9?!
&8!.&3&2$G&'67$8865-62!>:;?!B+'27-(#6!36''5!>-</01."%376=:!H65)'-5!$86!-;6!<6$.!I! /:J:!=&8!-;866!+.26#6.26.-!6C#68+<6.-5!$.$'(G62!+.!-8+#'+3$-6:!>>?!@33)<)'$-+&.!&=! "#$%!#&'()*+,)+-+.$-+&.!>=8&<!#'$5<+2!#@K4LE!-&#!#$.6'?!&8!-&-$'!L(37)*+,)+-+.! >*&--&<!#$.6'?!+.!B+'2!-(#6!MN4O4%!36''5!;$8*&8+.9!3&##687+.2)3+*'6!<(37)*+,)+-+.! $.2!$8865-62!+.!"%!#;$56!B+-;!!7=$3-&8!>!#??E!+.!/!#;$56!B+-;!;(28&C()86$!>@A?E!&8
sufficient for Gpa1 phosphorylation in vitro. Elm1 is expressed primarily in S and
G2/M phases of the cell cycle. Correspondingly, we found that the G protein is
phosphorylated in a cell cycle-dependent manner. Gpa1 is polyubiquitinated by SCF,
and phosphorylation is typically required for SCF-mediated ubiquitination. Thus we
investigated whether phosphorylation leads to ubiquitination of Gpa1, and whether
ubiquitination also occurs in a cell cycle-dependent manner. Indeed, whereas Gpa1
phosphorylation peaks in G2/M phase, ubiquitination occurs in the subsequent G1
phase. These findings establish that G proteins can be regulated by progression
through the cell cycle. More broadly, they raise the possibility that other pathway
components may also be subject to cell cycle regulation.
The participation of Elm1 in G protein signaling was unexpected. Elm1 is best
known as a protein kinase that coordinates events leading to cell division, including
bud emergence, mitosis and cytokinesis (1, 4, 8, 26, 45, 49). In this capacity, Elm1
phosphorylates protein kinases involved in septin organization and cytokinesis (Gin4
and Cla4) (1, 4, 45), another kinase that phosphorylates and de-activates the
morphogenesis checkpoint protein Swe1 (Hsl1) (8, 49), and a fourth kinase that
inhibits the mitotic exit network when the spindle position checkpoint is activated
(Kin4) (6). It is unlikely that Gpa1 is regulated by any of these Elm1 substrates, given
that deletion of those genes has no effect on Gpa1 phosphorylation or pheromone
responsiveness. Thus Gpa1 represents a new target of Elm1.
Whereas elm1Δ was the only mutant that exhibited a loss of Gpa1
phosphorylation, some residual phosphorylation could still be detected in vivo (Fig.
Possibilities include kinases that are absent from the gene deletion array, kinases
essential for cell viability, and kinases that have functions related to Elm1. Elm1 is
one of three closely related kinases, including Tos3 and Sak1. All three proteins are
known to phosphorylate and activate Snf1 (the yeast homolog of human AMPK
(AMP-activated protein kinase), primarily under conditions of glucose starvation (19).
Under the normal growth conditions used to assess mating pheromone responses
however, deletion of TOS3, SAK1, or SNF1 does not appear to affect Gpa1
phosphorylation or pheromone signaling (Fig 2.2). Taken together, our findings
indicate that Elm1 is largely responsible for Gpa1 phosphorylation, is uniquely able
to regulate the pheromone response pathway, and does so in a cell cycle-dependent
manner.
Other lines of evidence support the model that Elm1 phosphorylates Gpa1.
First, Gpa1 is neither phosphorylated nor ubiquitinated when Elm1 is absent. In
contrast, we did not observe any such differences in the absence of 108 other
kinases. Second, we did not observe any differences in the absence of 31 different
protein phosphatases. Thus it is unlikely that Elm1 acts indirectly by inhibiting the
function of a Gpa1-phosphatase. Third, we detected a substantial accumulation of
phosphorylated Gpa1 in the absence of the SCF function. Finally, we detected a
substantial enrichment of Gpa1 polyubiquitination in G1-arrested cells as compared
with S or G2/M-arrested cell cultures. These data confirm that phosphorylation and
ubiquitination occur in a cell cycle-dependent manner and that phosphorylation
precedes ubiquitination.