Copyright 0 1996 by the Genetics Society of America
Reduced Germline
Mobility of
a mariner Vector Containing
Exogenous DNA
Effect of Size or Site?
Allan
R. Lohe
and
Daniel L.
Hard
Department of Organismic and Evolutionary Biology, Haruard University, Cambrzdge, Massachusetts 021?8
Manuscript received February 14, 1996 Accepted for publication April 18, 1996
ABSTRACT
Germline mobilization of the transposable element mariner is severely inhibited by the insertion of a 4.5- to 11.9-kb fragment of exogenous DNA into a unique Sac1 site approximately in the middle of the 1286bp element. In the presence of transposase driven by the germline-specific hspZGsgs? promoter, mobilization of the MlwB construct (containing a 11.9-kb insertion) is detected at low frequency. Analysis of a mobilized MlwB element indicated that mobilization is mediated by the marinertransposase. However, transposed MlwB elements are also defective in germline mobilization. Rare, transposase-induced germ- line excision events were also recovered for such vectors. The estimated rate of excision is <0.1% per chromosome per generation. Excision appears to be accompanied by gap repair if a suitable template is available. The data imply that the reduced mobility of mariner vectors with exogenous DNA in the SacI site results from disruption of sequences necessary for efficient mobilization. The relative stability may be a valuable property in the uses of mariner-like elements in genetic engineering of insects of economic importance.
M
ARINER is a member of the Tcl-mariner superfam- ily of transposable elements having an ancientevolutionary history (ROBERTSON 1995). The phyloge-
netic distribution is typified by mariner and mariner-like
elements (MLEs), which are present in the genomes of
an unusually wide variety of insects and other organisms
(LIDHOLM et al. 1991; ROBERTSON 1993; ROBERTSON and MACLEOD 1993; GARCIA-FERN~DEZ et ul. 1995; LOHE et
al. 1995a), including some vertebrates (MORGAN 1995).
MLEs appear to persist through evolution by their abil- ity to undergo horizontal transmission into diverse hosts
( M A R U Y A M A and HARTL 1991; LOHE et al. 1995a; ROB-
ERTSON and LAMPE 1995), suggesting that MLEs have
evolved highly efficient mechanisms to invade and
spread within a genome. The principal events in the “life history” of MLEs are as follows: introduction of
an active copy into a susceptible host via horizontal
transmission, increase in copy number of the element within the host genome, vertical inactivation by muta- tion, and, eventually, stochastic loss of multiply mutated
elements (LOHE et al. 1995a).
Mariner is a relatively small transposable element, 1286 bp in length, with 28-bp terminal inverted repeats. The element inserts into the dinucleotide TA, which is
duplicated upon insertion. The element codes for a
single transposase protein of 345 amino acids (JACOS
SON et al. 1986) that is active in both the germline and
the soma. To date, the only known class of functionally
active MLEs is represented by Mosl, an autonomous
Cmesponding author: Daniel L. Hartl, Department of Organismic and Evolutionary Biology, Harvard University, 16 Divinity Ave., Cam- bridge, MA 02138. E-mail: [email protected]
copy of mariner isolated from Drosophila mauritiana
(MEDHORA et al. 1991). Other MLEs that have been
examined are nonfunctional either because of missense
mutations in the transposase open reading frame or
because of small deletions or point mutations that
frameshift the open reading frame. Although Mosl is
not normally present in D. melanogaster, a sibling species
of D. mauritianu, Mosl integrates efficiently into the germline when injected into the pole plasm of embryos
(GARZA et al. 1991). Mosl also functions as a vector for
the introduction of foreign DNA into L). melanoguster
(LIDHOLM et al. 1993). The potential of mariner to serve as a general transformation vector in insects has been
demonstrated by the transformation of M o d into
L).
virilis, a species that last shared a common ancestor
with D. melanogaster >40 million years ago (LOHE and
HARTL 1996).
In previous transformation experiments, we observed
that the MlwB element, which contains 11.9 kb of exog-
enous DNA in the SacI site, was apparently refractory
to germline mobilization induced by Mosl (LIDHOLM et
al. 1993). Furthermore, MlwB is mobilized inefficiently,
or not at all, in the soma in the presence of Mosl trans-
posase under the control of the dual Mosl and heat-
shock$rotein 70 ( h70) promoters (LOHE et al. 1995b), even with heat-shock induction. However, the previous experiments on germline mobilization were small in scale and therefore not definitive; a low frequency of transposition or excision would have been missed. Fur- thermore, the mechanism of the reduced mobility was not ascertained. On the one hand, it could result from
a size requirement whereby mobilization of mariner ele-
ments much greater than -1.3 kb is inhibited. On the
other hand, it could result from the disruption of se- quences near t h e SacI site that are necessary for efficient recognition and mobilization. In this paper, we present a detailed analysis of the apparent stability, with primary focus o n t h e following questions: (1) Does germline mobilization take place in the presence of mariner trans- posase driven by the germline-specific hsp26-sgs-3 (h26)
promoter? (2) Does the size of exogenous DNA in the construct affect germline mobilization? (3) Is t h e stabil- ity specific t o ( a n d p e r h a p s a result oQ chromosomal integration via t h e mariner transposase? We r e p o r t t h a t in the presence of high levels of transposase the stability of mariner transposons carrying a white' reporter gene
is not absolute. Rare germline events of transposition
and excision have been identified. Furthermore, exami- nation of some of the excision products yields evidence of template-dependent
gap
repair, analogous to thatreported previously for the
P
element (GLOOR et al.1991; NASSIF et al. 1994).
MATERIALS AND METHODS
Genetic crosses: Crosses were carried out at 25" on stan- dard cornmeal-molasses medium. Descriptions of mutants and balancer chromosomes can be found in LINDSLEY and
ZIMM (1992). Three different experiments were carried out
to screen for germline mobilization of mariner transposons that harbor exogenous DNA one experiment with the MlwB transposon that contains w+ as a marker (line M108; MlwB is located on chromosome
3
LIDHOLM et al. 1993) and two experiments with the M789 [mini-white] transposon, one with an autosomal copy (line M256, located on chromosome 3) and one with an X-linked copy (line M325) (LOHE et al.1995b). The sources of transposase were P-element constructs that carry an engineered mariner transposase gene driven by germline-specific or heat-inducible promoters, which have been characterized previously (LOHE et al. 199513). h7@182 refers to a Pelement construct,
Ry',
hsp7O:Mosl], inserted into chromosome 2. In h70-182, transcription of the Mosl open reading frame is under the control of an h70 promoter juxtaposed to the endogenous Mosl promoter. h26-67 refers to a P-element construct, P[ly', hsp26Sgs3:Mosl], also in- serted into chromosome 2. h26-97 is an insertion of the sameP[yr', hsp2&Sgs3:Mosl] construct on chromosome 3. In h26-
67 and h26-97, transcription of Mosl is under control of the dual promoters Sgs-3 and Mosl and is driven by the germline- specific enhancer hsp26. To induce the hsp7Opromoter, larvae in vials were heat shocked at 37" for 1 hr at days 5, 7 and 9 after hatching.
Attempted mobilization of the MlwB transposon: Males of the enotype y w / K h7@182/+; TM3,Sb/+ were crossed with
w1I1F M108 females. F1 progeny were subjected to heat shock for 1 hr at 37" during larval development at days 5, 7 and 9, and Sb flies were crossed to w"". Half of the F1 progeny are of the desired genotype, w; h7@182/+; M108/TM3,Sb. Sibs that lack the h7@182 transposase gene also contribute to the progeny. Tester adults were mass-mated in 120 bottles for males and 24 bottles for females, and a minimum of 500 progeny were scored for each cross, or a total of at least 70,000 flies. Putative jumps from chromosome 3 to another chromosome appear in the F2 progeny as w+ Sb individuals.
Attempted mobilization of the X-linked M789[mini-whitel transposon: Females of the genotype M325/yw; CyO/+ were crossed with yw; h26-67 males and M325/ Y; h26-67/ Cy0 sons
were crossedwithFM7ufemales (note that wand Bare present on FM7u). From this cross, single females of the genotype FM7u/M325; h26-67/+ were crossed with w1'18 males. Sons of the genotype w+B represent potential jumps. The M325 target transposon is present with the transposase source in both the F1 males and F2 females in this scheme. The hsp26 sgs3 promoter is expressed strongly in the female germline
(FRANK et al. 1992), and for this reason, FP females were gener-
ated before scoring for possible mobilization events.
Attempted mobilization of the autosomal M789[mini-white] transposon: Females of the genotype yw; M256/ TM3,Sb were crossed with yw/K h26-67/+; TM3,Sb/+ males. F1 females of the genotype yw/yw; h26-67/+; M256/TM3,Sb were crossed with w1I1* males and w"Sb progeny were scored as jumps of the transposon from chromosome 3 to another chromosome. The genotype of the F1 mothers with respect to h26-67 was verified by PCR with primers to Mosl sequences.
Lines homozygous for transposon and transposase source:
Stocks were constructed in which either the MlwB or M789[mini-white] elements were homozygous on one pair of major autosomes, and either the hsp70- or hsp26-sgs3-Mosl transposase sources were homozygous on the other pair of autosomes. For example, to construct lines homozygous for the h26-67 transposase source on chromosome 2 and the MlwB or M789[mini-white] transposons on chromosome 3, flies of the genotype w; h26-67/+; +/TM3,Sb were crossed with w;
+/
CyO; MlwB/+. Single pair matings of the genotype h26-67/ CyO; MlwB/ TM3,Sb were established and balancer chromosomes were removed in the following generations. To verify that h7@182or h26-67were present on chromosome 2, or h26-97 on chromosome 3, the parents were sacrificed for PCR when larvae were visible in the vial. PCR primers to Mosl sequences spanned the SacI site into which the exogenous DNA had been inserted, and consequently, only sequences from the mariner transposase source, rather than from the MlwB or M789[mini-white] transposons, were amplified. Mul- tiple lines were established for most combinations of geno- types, although in some of the lines homozygosity resulted in lethality or sterility and it was not possible to remove the balancer chromosome or chromosomes.DNA manipulations: Details of methods for DNA isolation, PCR and DNA sequencing can be found in LOHE et al. (1995a) and LOHE and HARTL (1996). The pPM789[mini-white] plas- mid was kindly provided by D.-A. LIDHOLM and was con- structed by digestion of the pM789 plasmid (LOHE et al.
199513) with Not1 and XhoI. A 5.8-kb fragment, containing the hsp70mini-whitegene bounded by inverted repeats from Mosl, was purified and the ends were made blunt with Klenow poly- merase. The fragment was ligated into the HpuI site of the P-
element transformation vector Carnegie 20 (RUBIN and SPRADLING 1983) to generate pPM789[mini-white].
The 5' genomic sequences flanking the MlwB transposons in lines M108 and M159 (LIDHOLM et al. 1993), and the 5' and 3' genomic sequences flanking the MlwB transposon in line M159J were cloned by inverse PCR with primers near the 5' and 3' ends of Mosl, as described (LOHE and HARTL 1996). PCR primers used to amplify the "empty" genomic DNA se- quence before integration of MlwB in line M108 and two
variant 3' primers that distinguish the donor and target geno- mic DNA sequences are illustrated in Figure 2. The common upstream 5' primer in the latter experiment was 5"GACATT- AATCAACAAGCCGAGJ'. Primers used to PCR amplify the flanking empty genomic DNA sequences before integration of MlwB in line M159 were 5'-TTGCGCCTTTCCCCGAAG CAG3' and 5'-CAATAGCGATTACAACTCCCG3'. PCR prod- ucts were cloned using a TA cloning kit (Invitrogen).
mariner Germline Stability 1301
gether with the wingsclipped helper P element at 150 pg/
ml, as described (LOHE et al. 1995b). Seven independent G1
transformants were recovered from a total of 184 GO adults.
RESULTS
Germline
stability
of the MlwB mariner transpo-son: The MlwB transposon carries an 11.9-kb insertion
of foreign DNA, including the w+ gene, in the SacI site at position 789 of the Mosl transposable element. In
a small experiment to mobilize MlwB with Mosl, no
germline transpositions were detected (LIDHOLM et al.
1993). One reason for the apparent germline stability of MlwB could be that Mosl is expressed too weakly in the germline for effective transposase recognition by
the terminal inverted repeats of MlwB, which are 13.2
kb apart. The h7U promoter is expressed in the germ- line at levels sufficient to promote a high frequency of
transposition of most P-element constructs, and the h7U-
182 transposase efficiently excises the nonautonomous
mariner element peach (LOHE et al. 1995b). Therefore,
the experiment to mobilize MlwB was repeated on a significantly larger scale, using h7U-182 transposase un- der heat shock conditions.
Line M108 carries an insertion of the MlwB element
on chromosome 3. Adults of the genotype h7U-182/+; M108/TM3,Sb were heat shocked and crossed to w- flies. Jumps of M108 from chromosome 3 to another chromosome would be detected as w+, Sbprogeny. Only two independent events of possible mobilization were recovered, but in both cases the w+ marker was linked
to Sb, demonstrating that the MlwB element was still
located on chromosome 3. It is unclear whether the two events represent jumps of MlwB between homologous chromosomes or instability in the TM3,Sb balancer chromosome. Activity of the h7U-182 transposase gene
was verified by somatic mosaicism of the wPCh allele
(LOHE et al. 1995b). In analogous crosses to mobilize
the P element, an individual produces on average one
jump in the F1 progeny (ROBERTSON et al. 1988). Conse-
quently, at least one jump per bottle, or -140 jumps in total, could have been expected in a P-element mobi-
lization experiment of similar size. Therefore, the
MlwB element is extremely stable in the germline in the presence of the h7U-182 transposase source.
Germline
stability
of M789[mini-white] transposons:Inefficient mobilization of Mosl constructs could result
from excessive size of the transposon, which, in the case
of the 13.2-kb MlwB element, is >10 times larger than
Mosl itself. Therefore, the germline behavior of the
M789[mini-white] transposon was examined in the pres- ence of transposase. At 5.8 kb in length, the M789[mini- white] transposon is less than half the size of MlwB and
the DNA is inserted into the same SacI site of the Mosl
vector, providing an appropriate basis for comparison.
TWO independent lines of the M789[mini-white] trans-
formants, M325 and M256 (LOHE et al. 1995b), were
tested for germline mobility. The source of transposase
was h26-67, which is highly active in the female germline
(FRANK et al. 1992; LOHE et al. 199513).
The M325 line has an X-linked insertion of the M789 [mini-white] transposon, which is somatically sta-
ble in the presence of h7U-182 (LOHE et al. 1995b).
Mothers of the genotype M325/FM7a; h26-67/+ were
crossed to FM7a males, and F1 sons were examined for
mobilization of M325 to the autosomes. Mobilization from the Xchromosome would be detected in sons that inherit the FM7a chromosome [marked with the Bar (B) mutation], together with the w" marker. In single crosses, 99 mothers were tested and 40 additional moth- ers were pooled in four bottles. From >5000 male prog- eny, no w+B sons were recovered, confirming that the M789[mini-white] transposon is stable in the presence of transposase.
The autosomal insertion of the M789 [mini-white] transposon in line M256 was also tested for germline mobility. In a small-scale experiment, 15 females of the genotype h26-67/+; M256/TM3,Sb were crossed indi- vidually to w- males. Jumps of the M256 transposon
from chromosome 3 to another chromosome should
be recovered as wfSb progeny. All progeny ( n = 20-
60) from each of the 15 crosses were either w+Sb+ or
w-Sb in phenotype, as expected from the segregation of
dominant markers on chromosome 3. In a comparable experiment with P-element mobilization, the expecta- tion is roughly one jump per "dysgenic" parent, or 15 jumps in total.
The resistance of the M789 [ mini-white] transposon
to germline mobilization parallels the germline stability
described above for the larger MlwB transposon in line
M108. We conclude that mariner vectors carrying 4.5- 11.9 kb of exogenous DNA in the SacI site are stable in the germline in the presence of mariner transposase, at least under the genetic and molecular conditions examined in these experiments. It remains a formal possibility that insertion of <4.5 kb of exogenous DNA
into Mosl does not disrupt transposon excision or trans-
position.
Rescue of 5' ends of MlwB transformants: The in-
tegrity of inverted repeats of transposable elements is essential for excision and reintegration of the element
elsewhere in the genome. For the two MlwB trans-
formants M108 and M159 (LIDHOLM et al. 1993), the
sequences of the 3"terminal repeats are intact. Never- theless, germline stability could still result from muta- tions within the 5"terminal repeats. Alternatively, the genomic sequences that flank the insertions may be unusual in organization or base composition.
The 5' regions of the MlwB element in lines M108
and M159 were cloned by inverse PCR (OCHMAN et al.
~ 1 0 8 :
. . .
TCACTTGCTCGCGTCCTGTACTTTAGGGATTAGACGATATATTATAT~TTACAC~GT~ . . .M159:
. . .
AATTGCCCCGCCGAGTTGTTTTGTTGTTTTTATGGCGCGCCTTAT~TCTCGATTGTT~GGCA . . .~ 1 5 9 ~ :
. . .
ATGTATTCAAAGATGATGCACGCATACTTTTACTTTCGAGCTT~CATCTG~CC~GATA . . .FIGURE 1.-Nucleotide sequences flanking three independent insertions of the MlwB transposon. The TA dinucleotide target
that is duplicated upon inseition is indicate2 in bold.
otide was present in the 5”genomic DNA adjacent to the insertion site. The genomic DNA sequences ap-
peared to be unique sequence DNA of normal base
composition for >lo0 bp flanking each insertion (Fig-
ure 1). The nucleotide sequences surrounding the in-
sertion site of the MlwB transposon in line M159J (see
below) are also presented in the figure. These results
suggest that the stability of the MlwB transposon is not
caused by mutational alterations within the inverted repeats or by unusual genomic sequences flanking the insertions.
Pelement-mediated transformation of a PM789[mini-
white] transposon:We also considered the possibility that mrinmmediated transformation results in molecular al- terations to DNA sequences at regions of the transposon that are essential for transposition but that are physically distinct from the inverted repeats. Therefore, the entire M789 [mini-white] transposon described above was in-
serted into the Pelement transformation vector Carnegie
20 using restriction enzyme sites distal to the mariner in-
verted repeats, and the construct was transformed into
the strain using the wingsclipped helper P element.
Seven independent
$
transformants were obtained.When the
n ~ + ,
M789(mini-white)] transformants werecrossed with h7@182, no mosaicism was observed in the
eyes of the F1 progeny, confirming the absence of somatic
excision of the M789(mini-white) mariner transposon
within the larger Ptransposon. To investigate the possibil-
ity that sequences intrinsic to the mariner construct are
generally inhibitory to transposon-mediated excision, the
transformants were crossed individually to the Pelement
transposase source A2-3(99B) (ROBERTSON et al. 1988).
Extreme somatic mosaicism was observed in the eyes for each of the crosses, confirming that the transposon, as defined by Pelement inverted repeats, responds normally to Pelement transposase.
Although not strictly comparable, an indication of the efficiencies of germline transformation can never-
theless be obtained for the mariner-mediated us. P-ele-
ment-mediated experiments. With mariner-mediated transformation, the efficiency for germline integration of the 5.8-kb M789[mini-white] construct was 0.6% (2/
320) (LOHE et al. 199513) but with P-element transforma-
tion of the 16.6-kb PM789[mini-white] construct, the
efficiency was 3.8% (7/184), more than six times
greater despite the significantly larger size of the P-
element construct.
These results suggest that germline stability of mariner
constructs are not due to molecular alterations that
occur to the plasmid before or during mariner-mediated transformation, or to “poison” sequences intrinsic to the M789[mini-white] transposon that are generally in- hibitory to transposon-mediated excision, but that sta- bility is a property of the mariner transposon itself. This
property could be associated with the presence of exog-
enous DNA in the vector, with the position of exoge-
nous DNA in the vector, or a combination of these
factors.
Rare events of MlwB germline mobilization: The ge-
netic screen for mobilization of the MlwB or
M789[mini-white] elements detects mobilization in a
single generation and utilizes a single copy of a transpo- sase source. To improve the chance for mobilization, MlwB/Mosl stocks were constructed that combined a homozygous target element with a homozygous trans- posase source. The target element is therefore under constant exposure to transposase. Mobilization of the transposon can be monitored over many generations, either phenotypically by changes in eye color, or geneti- cally by segregation of dominant markers. A low level of mobilization of MlwB was demonstrated by both methods.
Flies of the genotype M159/M159; h26-97/h26-97
normally have a bright red, nonmosaic eye color, but a single female with an orange (nonmosaic) eye color was discovered in one of the lines. The orange eye color
mapped to chromosome 3 rather than to chromosome
2, the location of w+ in the parental M159 line. The
transposon in line M159J, derived from the exceptional
chromosome, was tested for mobility over many genera-
tions in a genetic background that was homozygous
for h26-67. Although further mobilization or excision should have been detected easily as a different eye color from the parents, the eye color phenotype was stable for over 12 months. The absence of further mobility of MlwB in line M159J does not support the hypothesis that the chromosomal environment of the MlwB ele-
ment in line M159 is inhibitory to mobilization.
Genetic results also suggested that the MlwB element is capable of mobilization at a low level. M159 heterozy-
gous females have a dull red eye color, in contrast to
the bright red exhibited by M159 homozygotes. Males from independent homozygous lines of M159; h26-97
described above were crossed to w- females, and daugh-
ters were scored for eye color. M159/+ females with a bright eye color were recovered at a low frequency and
were crossed to w- males. Two lines in which the w+
mariner Germline Stability 1303
other M159; h26-97 line, in which the h26-97 chromo- some was balanced over TM3,Sb due to the presence of
a lethal, the wi marker had become linked to the
TM3,Sb balancer chromosome. (In generating the ho-
mozygous MlwB/h26-Mosl lines, it was not always possi-
ble to remove one or both balancer chromosomes be- cause some tester chromosomes were lethal when
homozygous.) These results suggest that either MlwB
has jumped to chromosome 3 or a chromosomal frag-
ment containing MlwB has become associated with
chromosome 3. There was no evidence for “local hop-
ping” of MlwB, which also would have been detected in the eye color screen.
Mobilization is mediated by mariner transposase: If
mobilization of the MlwB element to chromosome 3
in line M159J were mediated by mariner transposase, a TA dinucleotide should flank the site of insertion at both ends. The genomic DNA sequences should also differ from those at the M159 insertion site. Therefore, the 5’ and 3’ ends of the M159J insertion were isolated by inverse PCR and cloned. Sequencing verified that a TA dinucleotide pair is present at both ends of the
insertion, and that the genomic DNA sequences sur-
rounding the M159 and M159J insertion sites bear no
resemblance (Figure 1). The inverted repeats at both
the 5’ and 3‘ ends of the element were also intact.
These results demonstrate that the MlwB jump from
chromosomes 2 to 3 in line M159J was a genuine mari-
ner-mediated event and that transposase can recognize
inverted repeats that are >13 kb apart.
Rare events of MlwB and M789[mini-white] germline
excision: Transposase-mediated excision events of the
MlwB or M789[mini-white] transposons can be detected
as w- individuals that are either homozygous for the
excision chromosome (if excision does not generate a
lethal), or as w-/Balancer heterozygotes in stocks that carry a balancer chromosome.
In MlwB or M789[mini-white] control lines that lack a mariner transposase source, the w” eye color pheno- type is stable. However, in 6/17 lines of the genotype MlwB/h26-Mosl, at least one w- individual appeared after several generations. Eight lines were recovered from independent excision events of the MlwB element in lines M159 and M108. Seven of the excision chromo- somes were homozygous viable and one was homozy-
gous lethal. An estimate of the excision frequency was
provided by scoring progeny from the cross h26-67/
CyQ M108/TM3,Sb X w- males. A single w-Sb+ individ-
ual was obtained from 1379 progeny, suggesting an exci- sion frequency of MlwB of <0.1% per generation.
Rare excision events of the M789[mini-white] element were also observed in the presence of h26”osl. In the screen for mobilization of the X-linked M789[mini-
white] element in line M325 described above, two fertile
w-B+ sons were recovered from -5000 male progeny
(frequency of 0.04%). The fertility of these males dem-
onstrates that the origin of their X chromosome was
the maternal M325 chromosome, in contrast to other exceptional w-@ sons that were sterile. (The sterile
sons were presumably X 0 in genotype and arose from
sex chromosome nondisjunction.)
Molecular analysis of MlwB excision events: The w-
phenotype described above could result either from precise excision of the element at the inverted repeat termini, or from imprecise excision events that resulted in deletions in the w” reporter gene. In the Pelement
gap repair model, excision of a P element results in
repair of DNA in the regions surrounding excision of the element, usually from a P-element template located on the sister chromatid or on the homologous chromo-
some (GLOOR et al. 1991). If the same gap repair mecha-
nism functions in chromosome repair following exci- sion of an MlwB element, the template for gap repair in MlwB; Mosl stocks should also be an MlwB element. However, the appearance of w- flies in these lines sug-
gests that if gap repair occurs after MlwB excision, then
excision is not always accompanied by complete repair
from the homologous MlwB element, which carries the
w” marker.
The molecular basis for the w- phenotype was exam- ined by PCR with primers to genomic DNA on the 5’
and 3’ sides of the MlwB insertion. PCR of DNA from
four independent, homozygous w- individuals from
M159; Mosl lines gave no product, although DNA from
control (wild-type) flies gave the expected 314bp band.
One interpretation of this result is that excision of the MlwB transposon is accompanied by deletions of geno- mic DNA that extend past one or both primers. Since
another w- chromosome from the M159; Mosl lines
was homozygous lethal, deletions may sometimes ex- tend into neighboring genes. Alternatively, the w- phe- notype could arise from w’ deletions internal to the
MlwB transposon, but the DNA remaining in the
transposon may be too large to be amplified by the flanking primers in PCR reactions.
Three individuals with a w- phenotype, representing
putative excision events of the MlwB transposon, were
also recovered in homozygous M108; Mosl lines. Each individual was examined by PCR using primers to ge-
nomic DNA 5’ and 3’ to the MlwB insertion. In con-
trast to the results with the M159 w- lines described above, a PCR product was obtained from each of the homozygous M108 w- lines, and the product was simi- lar in size to the expected 159-bp band observed with wild-type DNA. Synthesis of a PCR product of roughly the correct length from DNA of M108 w- individuals
is surprising, because the MlwB transposon is homozy-
gous and the only available template for repair after excision of one MlwB copy should be the remaining MlwB copy. Repair from an MlwB template should re- store the w+ phenotype. Several lines of evidence con- firm that the PCR product from w- individuals derives
from the genomic region at the site formerly occupied
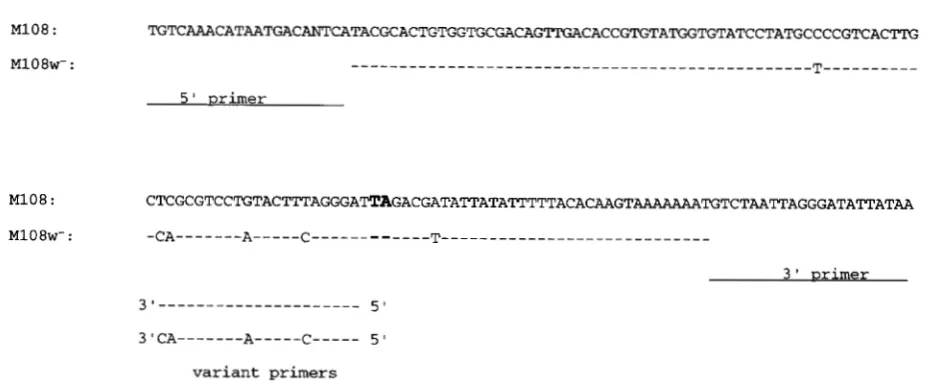
M108w-: -CA---A---C---T---
3 wrimer
3'""""""""""- 5 '
3'CA---A---C--- 50
variant primers
FIGURE 2.-Comparison of sequences surrounding the insertion site of the MlwB transposon in line M108 with sequences
recovered following excision of the element. The TA target site that is duplicated upon insertion of the transposon is shown in
bold, and the nucleotide sequence following excision (identical in three independent excision events) is shown beneath, with
nucleotide alterations indicated. The 21-bp primers used for PCR amplification of the region flanking the MlwB insertion are shown at either end of the M108 sequence. The 21-bp region from which two variant primers were synthesized, one with the
upper sequence and one with the lower sequence, is also indicated.
genome. First, no PCR product was obtained from flies
homozygous for the MlwB insertion in line M108, pre- sumably because under the conditions employed, the primers are too far apart to synthesize a 13.2-kb prod-
uct. However, a 159-bp band was obtained using DNA
from either a wild-type strain or from the M159 line, which differs from line M108 only in the genomic loca-
tion of the MlwB element. Second, a 159-bp PCR prod-
uct was obtained when DNA from M108 w- heterozy-
gotes was used as template, demonstrating that the
appearance of the PCR product is dependent upon
the w- chromosome.
Sequencing the cloned PCR products gave an unex-
pected result. Although identical to each other, the
sequences of the 159-bp PCR products from the three
homozygous w- M108 lines differed at six positions out
of 117 bp from the "empty" sequence before MlwB
insertion (Figure 2). There were five nucleotide alter-
ations in 34 bp on the 5' side of the TA insertion point
and one alteration on the 3' side. The wild-type DNA
sequence at the site of the MlwB insertion was deter-
mined in two ways, either by PCR using wild-type DNA
as template or from inverse PCR of the MlwB element
(Figure 1). Both experiments yielded the same se-
quence. The nucleotide differences before and after
excision of MlwB are unlikely to result from PCR or
cloning artifacts because the sequences were identical
in clones from independent PCR reactions using the
same template and in clones from three independent
w- excision lines.
A possible explanation for the result is that the M108
w- lines arise from mariner-mediated complete excision
of the MlwB element, and that the gap is subsequently repaired from a donor template that is almost identical
in nucleotide sequence to the flanking DNA sequence.
Evidence for such a donor sequence was obtained from
additional PCR experiments. One region on the 5' side
of the MlwB insertion site differs at four positions be-
tween the sequences from the wild-type and M108 w-
lines. Two primers were synthesized from the same 21- bp region, one using the wild-type sequence and the
other incorporating the four nucleotide differences, in-
cluding two at the 3' terminus (Figure 2). Since the
two 3' nucleotides of the variant primers are different,
they are unable to amplify the same region of genomic DNA. In PCR amplifications of wild-type DNA, a prod-
uct of the expected length of -200 bp was obtained
when either of the variant primers was tested with a common primer further upstream of the region. The result was identical with a different upstream primer and each of the variant primers. These results confirm
that the region of the genome represented by the MlwB
insertion in line M108 is duplicated, and that one re- gion can serve as a donor sequence for gap repair fol- lowing excision of the MlwB element from the other region.
The target and donor sequences differ at six positions
over 117 bp, and it is likely that additional sequence
variations are present between target and donor DNAs
in the region 5' and 3' to the MlwB insertion, immedi-
ately outside of the 11 7-bp region that was used to syn-
thesize the original PCR primers (Figure 2). Recall that
no PCR product was obtained from DNA of MlwB ho-
mozygotes in line M108, using primers 5' and 3' to
the MlwB insertion. Although such a product was not expected to originate from the site of the large MlwB insertion, a product may have been expected from the
donor sequence elsewhere in the genome. The absence
of such a PCR product suggests that the donor sequence
mariner Germline Stability 1305
a sequence is amplified in PCR reactions with DNA
from the M108 w- lines with the same primer pairs and
this sequence derives from the donor template. The
paradox is simply resolved if repair from the donor
sequence did not extend into one or both regions used
to synthesize the 5' and 3' primer sequences. We con-
clude that both the MlwB and "789[mini-white] ele-
ments can, on occasion, excise in response to mariner transposase, and that in the absence of a template for repair, excision is accompanied by a deletion at the site of excision. However, if a template is available elsewhere in the genome, the deletion is repaired from that tem-
plate according to the gap-repair model (NASSIF and
ENGELS 1993; NASSIF et al. 1994).
DISCUSSION
The transposable elements P, hobo, mariner, Minos
and Hemes can serve as vectors for the introduction of
exogenous DNA into D. melanogaster (RUBIN and
SPRADLING 1982; BLACKMAN et al. 1989; LIDHOLM et al.
1993; LOUKERIS et al. 1995; O'BROCHTA et al. 1996).
With the exception of the mariner-based constructs dis- cussed in this paper, any of these transposons with in- serts of exogenous DNA are mobilized efficiently in the germline and soma in the presence of their respective transposase sources. In the mariner constructs, which have either 4.5 or 11.9 kb of exogenous DNA inserted
into the SacI site of Mosl, somatic excision of the
transposon is severely inhibited. These constructs also exhibit great, but not complete, stability in the germ- line, even in the presence of a germline-specific source of transposase. Relative to the stability, one can rule out the hypothesis that insertion via the marinertransposase somehow interferes with the ability of the transposon to be mobilized subsequently, because the M789 [mini- white] transposon, which was inserted via P-element transformation, was also stable in the soma in the pres- ence of mariner transposase.
Because excessive size of the insert in P-element con-
structs can dramatically diminish the frequency of trans-
formation and transposition, one might expect a
priori
that the stability of MlwB was most likely a result of the
large size of the exogenous DNA insertion in the vector.
Contrary to this expectation, the M789 [mini-white]
transposon was also stable in the germline, even though
this construct is less than half the size of MlwB. We
have not tested the possibility that mariner mobility is inhibited by any inserted DNA fragment, no matter how small, but this seems unlikely in view of the results with Minos, which is also a member of the Tcl-mariner super- family and in which transposons with an insertion of exogenous DNA are efficiently mobilized in both the
soma and the germline (LOUKERIS et al. 1995). (The
insertion site of exogenous DNA in Minos is within an intron, but there are no introns in mariner.)
A second hypothesis for the germline stability of
the MlwB transposon is that the reduced frequency of mobilization results from DNA sequences neighboring the insertion. In this case, the transposon should re- gain its mobility after transposition to a new chromo-
somal location. However, the transposed MlwB ele-
ment M159J remained stable in the soma and germline in the presence of h26-Mos2, suggesting that suppres- sion of transposition is associated with the MlwB transposon itself rather than with the genomic site. The result also demonstrates that mariner transposase can catalyze transposition by recognition of transpo- son ends that are at least 13 kb apart. Taken together, the results suggest that full mobility of mariner con-
structs requires continuity of a region near the SacI
site, which is disrupted in the insertions. It seems rea-
sonable to relate the reduced mobilization of SacI in-
sertions to the inefficiency of germline transforma-
tion, which is only 0.5% with these constructs
(LIDHOLM et al. 1993; LOHE et al. 1995b), -20-fold
lower than the transformation efficiency of Mosl itself
(LOHE et al. 1995b). How it is that MLEs in the human
genome have generated a large number of copies con- sisting of little more than the inverted repeats remains
to be determined (MORGAN 1995).
By means of
fly
stocks that are homozygous for boththe MlwB target transposon and the h26-MosI transpo- sase source, it was possible to screen for rare events of MlwB germline transposition or excision over many
generations. Both transposition and excision of MlwB
were detected by this method, and transposition was a genuine mariner-mediated event by molecular criteria. Germline excision of the M789 [mini-white] transpo- sons was also detected by this method; it takes place at an estimated rate of 0.04% per chromosome per generation.
Excision of marinerresults in a characteristic footprint
(BRYAN et al. 1990), but excision of the MlwB element
resulted in different outcomes in the two independent transformant lines M108 and M159. Given its member-
ship in the same transposon superfamily as Tc3, it seems
likely that mariner transposes in a cut-and-paste mecha-
nism analogous to that of Tc3, in which 3' overhangs
created by the initial cleavage form a heteroduplex re- paired in one way or the other by host mismatch repair
(VAN LUENEN et al. 1994). In the case of mariner, the resulting footprint consists of three base pairs from ei-
ther end of mariner plus the TA duplication (BRYAN
et al. 1990). In M108 excision, however, the gap was
apparently repaired using, as a template, a distinct se-
quence differing at six nucleotide positions out of 117
from the sequence originally present at the genomic
site before the MlwB insertion. Template-dependent gap repair of this type has been reported previously for
the P element (GLOOR et al. 1991; NASSIF et al. 1994).
The template-dependent gap repair with mariner would
not have been observed without the fortuitous insertion
suitable template sufficiently homologous to the flank-
ing genomic DNA.
Relative to the big picture, transposon mobility has been used to advantage in the study of gene function in D. melunogaster, by insertional mutagenesis or moni-
toring expression of an enhancer trap at different sites
in the genome. Germline stability of mariner transpo-
sons that harbor exogenous DNA would preclude their
use in similar experiments. However, in the context
of genome modification, transposon stability may be
preferred because of the possibility of uncontrolled mo-
bilization of a transposon that has been genetically engi- neered. Further studies on factors causing the stability of mariner transposons that carry exogenous DNA are necessary to increase our knowledge of the molecular
mechanisms of transposition of mariner as well as to
enhance its potential utility in molecular genetics and molecular entomology.
This work was supported by grant GM-33741 from the National Institutes of Health.
LITERATURE CITED
BIACKMAN, R. IC, M. M. D. KOEHLER, R. GRIMAILA and W. M. GELBART, 1989 Identification of a fully-functional hobo transposable ele- ment and i t s use for germline transformation of Drosophila.
BRYAN, G., D. G mand D. HARTL, 1990 Insertion and excision of the transposable element manner in Drosophila. Genetics 125
103-114.
FRANK, L. H., H.-K CHEUNC and R. S. COHEN, 1992 Identification and characterization of Drosophila female germ line transcrip tional control elements. Development 114: 481-491.
GARCIA-FERN~DEZ, J., J. R. BAYASCASRAM~REZ, G. MARFANY, A. M. MUNOZ-MAIZMOL, A. CASALI et al., 1995 High copy number of highly similar mariner-like transposons in planarian (Platyhelmin- the): evidence for a trans-phyla horizontal transfer. Mol. Biol. Evol. 1 2 421-431.
GARZA, D., M. MEDHORA, A. KOCA and D. L. HARTL, 1991 Introduc- tion of the transposable element manner into the germline of Drosophila mlanogaster. Genetics 1 2 8 303-310.
GLOOR, G. B., N. A. NASSIF, D. M. JOHNSON-SCHLITZ, C. R. PRESTON and W. R. ENGEIS, 1991 Targeted gene replacement in Drosoph- ila via P-element-induced gap repair. Science 253: 11 10- 11 17. JACOBSON, J. W., M. M. MEDHORA and D. L. HARTL, 1986 Molecular
structure of a somatically unstable transposable element in Dro sophila. Proc. Natl. Acad. Sci. USA 8 3 8684-8688.
LIDHOLM, D.-A,, G. H. GUDMUNDSSON and H. G. B o w , 1991 A highly repetitive, mariner-like element in the genome of Hyalo
phma cecropia. J. Biol. Chem. 266: 11518-11521.
LIDHOLM, D.-A,, A. R. LOHE and D. L. HARTI., 1993 The transposable element manner mediates germline transformation in Drosophila melanogaster. Genetics 134: 859-868.
LINDSLEY, D. L., and G. G. ZIMM, 1992 The Genome of Drosophila melanogaster. Academic Press, New York.
EMBO J. 8: 211-217.
LOHE, A. R., and D. L. HARTL, 1996 Cermline transformation of Drosophila aidis with the transposable element mariner. Genetics
LOHE, A. R., E. N. MORIYAMA, D.A. LIDHOLM and D. L. HARTL, 1995a Horizontal transmission, vertical degeneration, and stochastic loss of mariner-like transposable elements. Mol. Biol. Evol. 12:
LOHE, A. R., D.-A. LIDHOI.M and D. L. HARTI., 1995b Genotypic effects, maternal effects and grand-maternal effects of immobi- lized derivatives of the transposable element manner. Genetics
140: 183-192.
LOUKERIS, T. G., B. ARC& I. LIVADARIS, G. DIALEKTAKI and C. SAVAKIS, 1995 Introduction of the transposable element Minos into the germ line of Drosophila mlanogaster. Proc. Natl. Acad. Sci. USA
MARUYAMA, K., and D. L. HARTI., 1991 Evidence for interspecific transfer of the transposable element mariner between Drosophila and Zap-rionus. J. Mol. Evol. 3 3 514-524.
MEDHORA, M., K. MARUYAMA and D. L. HARTI., 1991 Molecular and functional analysis of the mariner mutator element Mosl in Dro
sophila. Genetics 128: 311-318.
MORGAN, G . T., 1995 Identification in the human genome of mobile elements spread by DNA-mediated transposition. J. Mol. Biol.
NASSIF, N., and W. ENGELS, 1993 DNA homology requirements for mitotic gap repair in Drosophila. Proc. Natl. Acad. Sci. USA 9 0
NASSIF, N. A,, J. PENNEY, S. PAL, W. R. ENGELS and G. B. GLOOR, 1994 Efficient copying of nonhomologous sequences from ectopic sites via P-element-induced gap repair. Mol. Cell. Biol. 1 4 1613-
1625.
O'BROCHTA, D. A,, W. D. WARREN, K J. SAVILLE and P. W. ATKINSON, 1996 Hermes, a functional non-drosophilid insect gene vector from Musca domestica. Genetics 142: 907-914.
OCHMAN, H., F. J. AYALA and D. L. HARTI., 1993 Use of the polymer- ase chain reaction to amplify segments outside the boundaries of known sequences. Methods Enzymol. 2 1 8 309-321. ROBERTSON, H. M., 1993 The manner transposable element is wide-
spread in insects. Nature 3 6 2 241-245.
ROBERTSON, H. M., 1995 The Tcl-marinersuperfamily of transposons in animals. J. Insect Physiol. 41: 99-105.
ROBERTSON, H. M., and D. J. LAMPE, 1995 Recent horizontal transfer of a manner transposable element among and between Diptera and Neuroptera. Mol. Biol. Evol. 12: 850-862.
ROBERTSON, H. M., and E. G. MACLEOD, 1993 Five major subfamilies of manner transposable elements in insects, including the Medi- terranean fruit fly, and related arthropods. Insect Mol. Biol. 2:
ROBERTSON, H. M., C. R. PRESTON, R. W. PHILLIS, D. JOHNSON- SCHLITZ, W. K. BENZ et al., 1988 A stable genomic source of P-element transposase in Drosophila melanogaster. Genetics 119:
75-83.
RUBIN, G. M., and A. C. SPRADLING, 1982 Genetic transformation of Drosophila with transposable element vectors. Science 218:
RUBIN, G. M., and A. C. SPRADLING, 1983 Vectors for Pelement- mediated gene transfer in Drosophila. Nucleic Acids Res. 11:
VAN LUENEN, H. G. A. M., S. D. COLLOMS and R. H. A. PIASTERK, 1994 The mechanism of transposition of Tc3 in C. elegans. Cell
79: 293-301.
143: 365-374.
62-72.
92: 9485-9489.
254 1-5.
1262-1266.
125-139.
348-353.
6341-6351.