Extrachromosomal Elements Cause
a Reduced Division Potential in
nibl
Strains
of
Saccharomyces cerevisiae
Rosemary Sweeney and Virginia A. Zakian
Fred Hutchinson Cancer Research Center, 1124 Columbia Street, Seattle, Washington 98104 Manuscript received July 22, 1988
Accepted for publication April 21, 1989
ABSTRACT
The nibl allele of yeast confers a sensitivity to an endogenous plasmid, 2r DNA, in that nibl strains bearing 2r DNA (cir+) exhibit a reduction in division potential. In the present study, the reduction in division potential characteristic of nibl cir+ strains is shown to be dependent on the simultaneous presence of both the A and the D open reading frames of 2r DNA as well as on the presence of an unidentified extrachromosomal element other than 2 p DNA. Furthermore, in nibl strains, an uncharacterized extrachromosomal element can cause a less severe reduction of division potential in the absence of intact 2~ DNA. Thus, the nibl allele may confer a generalized sensitivity to extrachro- mosomal elements.
V
IRTUALLY all strains of Saccharomyces cerevisiae harbor 50-1 00 copies of an endogenous plasmid of 6318 base pairs called 2p DNA. In most cells, replication of 2~ DNA occurs exactly once during the S phase of the cell cycle (ZAKIAN, BREWER and FANG- MAN 1979) and requires the same gene products that control chromosomal DNA replication (LIVINGSTON AND KUPFER 1977; PETES and WILLIAMSON 1975). T h e 2p plasmid has been completely sequenced (HART- LEY and DONELSON 1980). A marked feature of its sequence is the presence of two 599 base pair inverted repeats between which recombination takes place, giving rise to two forms of the plasmid (BEGGS 1978). There are also five and one-half directly repeated copies of a 62-63 base pair sequence. This region (STB or REP3) is necessary in cis for the accurate partitioning of the plasmid during mitosis (KIKUCHITON and BROACH 1985). T h e plasmid contains an ARS (autonomously replicating sequence; BROACH and HICKS 1980) that functions as an origin of DNA replication (NEWLON et al. 1981 ; BREWER and FANG- MAN 1987; HUBERMAN et al. 1987). There are five open reading frames (ORFs), one of which, E, is small and has no known function. T h e A ORF encodes the FLP protein, which is required to catalyze recombi- nation between the inverted repeats (BROACH and HICKS 1980) and to facilitate amplification of 2p DNA copy number in situations where copy number is low
(VOLKERT and BROACH 1986; REYNOLDS, MURRAY and SZOSTAK 1987). T h e D ORF has been shown to positively regulate the amount of FLP gene product (MURRAY et al. 1987). T h e B and C ORFs (REP1 and
1983;JAYARAM, LI and BROACH 1983;JAYARAM, SUT-
The publication costs of this article were partly defrayed by the payment
of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. $1734 solely to indicate this fact.
Genetics 122: 749-757 (August, 1989)
REPB) are required for stable plasmid maintenance
and accurate partitioning during mitosis (JAYARAM,
LI and BROACH 1983; KIKUCHI 1983). They also play a role in negatively regulating the amount of the FLP gene product (MURRAY et al. 1987).
In most strains, 2 p DNA confers no phenotype: cells that have 2 p DNA (cir+ cells) and cells that lack 2p DNA (cir’ cells) are indistinguishable in terms of gross morphology and growth rates (FUTCHER and COX
1983). However, a chromosomal mutation called nibl confers sensitivity to the presence of 2 p DNA (HOLM
1982a,b). A nib1 cir+ strain produces irregularly shaped colonies with ragged edges (nibbled colony morphology). In contrast, nibl cir’ cells produce smooth, round, wild-type colonies (see Figure 1). T h e nib1 cir+ strains also have a subpopulation (>lo%) of large, abnormal-looking cells that fail to divide and that have an elevated copy number of 2 p DNA (Figure
750 R. Sweeney and
karl-1 S. cerevisiae strains, cytoductants expressing the S. carlsbergensis nuclear genotype and receiving 2p DNA displayed nibbled colony morphology. T h e allele conferring this sensitivity to 2p DNA, called nihl, was shown to segregate as a single recessive nuclear gene that mapped to chromosome XVZ, adja- cent to RAD1 (HOLM 1982a).
Since the copy number of 2p DNA is abnormally high in nibl cir+ strains and the presence of 2p DNA is essential for the expression of the Nib- phenotype, it was hypothesized that in wild-type cells the NIB1 gene product interacts with 2p DNA and in so doing limits replication of the plasmid to a single round in each cell cycle. In the absence of the NIB1 gene product, the amount of 2p DNA increases, and this increase was hypothesized to disrupt chromosomal replication or segregation (HOLM 1982b).
In the present work, we take a closer look at the interaction of nibl and 2p DNA. The data indicate that the recessive nibl mutation and two extra-chro- mosomal elements, 2p DNA and at least one unde- fined element, are necessary for maximal expression of the Nib- phenotype. Moreover, an undefined ex- trachromosomal element can cause a less severe Nib- phenotvpe in nibl strains in the absence of 2p DNA. In addition, show that maximal expression of the Nib- phenotype is dependent on either the presence
V. A. Zakian
of intact 2p DNA or on the simultaneous presence of the A and D ORFs.
MATERIALS A N D METHODS
The yeast strains used in this study are listed in Table 1.
Media are described in ZAKIAN and SCOTT (1982). 34Ncir' was made by B. VELT by transforming 3482-16-1 with
pJDB219, a 2p based plasmid containing a defective allele of LEU2 that can be used to cure a strain of 2p D N A (DOBSON, FUTCHER and COX 198O), and screening for cir" isolates. 34ncir" and 34ncir'a are the products of seven sequential backcrosses of nib1 progeny to 34Ncir". The n i b l allele is originally from the cir" strain 5Dcyc-cyto#l. In constructing 34ncir" and 34ncir"a the presence of nibl was established by determining the phenotypes of diploids ob- tained by crossing segregants to nibl cir+ tester strains. Plasmids used in this study are shown in Figure 2. YCpL2 was constructed by K. RUNGE by inserting the 2.2-kb SalI- Xhol fragment bearing the yeast LEU2 gene (ANDREADIS et al. 1982) and the 2.7-kb Hind111 fragment bearing ARSl and CEN4 from YCpl9 (STINCHCOMR, MANN and DAVIS
1982) into pUCl9 (YANISCH-PERRON, VIEIRA and MESSING 1985). 82-6B-LEU2 was constructed by B. VEIT by inserting the SalI-XhoI fragment bearing LEU2 into the Sal1 site of
the pMB9 vector portion of plasmid 82-6B that contains one and one-half repeats of 2p DNA (HARTLEY and Donel- son 1980). Yeast strains bearing plasmids are denoted as follows: strain name/plasmid name. The presence of the nibl allele was assayed in haploid t i l + or cir" strains by crossing to a nib1 cir+ haploid tester strain (either 2101-3-
3 or 2103-6-2) that had nibbled colony morphology. Col- ony morphology of t h e resulting diploid was determined by streaking for single colonies and scoring multiple colonies for morphology after growth for 1 week on Ymin plates at
30".
Quick preparation of yeast D N A for determining the presence or absence of plasmids was performed essentially by the methods of A. LORINCZ as described in FOCUS (6(4), 1984). Cells from a colony (about 3 m m in diameter) or from an equivalent number of cells from a streak were transfered into 200 pI of 100 mM NaCl, 10 mM Tris-HCI (pH K O ) , 1 mM EDTA, 0.1 % SDS. Acid-washed glass beads were added to just below the meniscus. T h e tubes were vortexed for 3 min prior to extraction with an equal volume of pheno1:chloroform (1 : 1). The aqueous phase was brought to a volume of 4 0 0 pl with 10 mM Tris-HCI (pH 7.5), 1 mM EDTA and precipitated by adding 40 pl of 5 M NH4Ac and
1 ml of ethanol. Total yeast D N A used for transformation was prepared using the method of DAVIS et al. ( 1 980) or the alkaline lysis method of FILETICI, JUNAKOVIC and BALLARIO
( 1 985). Yeast colony hybridizations were performed using the method of HINNEN, HICKS and FINK (1 978). SOUTHERN
( 1 975) blots and hybridizations (WAHL, STERN and STARK 1979) were performed essentially as described.
Transformations were performed either by the sphero- plast method (BEGGS 1978) or by the lithium acetate method
(ITO et al. 1983). T h e 2 p plasmid was introduced in either
of two ways. In the first method, total yeast D N A from a cir+ strain that also contained a centromere plasmid or 2p
D N A purified from a two-dimensional agarose gel (PLUTA
Yeast Extraachromosomal Elements
TABLE 1
Strains used in this study
Strain Genotype Source
3482-16-1 dlATa met2 his?-I leu2-?,1 12 frpl-289 ura?-52 [cir+] 1.. HARTWELL
34Ncir" .\IATa me12 his3-1 leu2-3,I 12 frpl-289 ura3-52 [cir"] B. VElT
2513-1 1 - 1 ,WATa or07 fhr4 rad1 His- Lys- Fcy' L. HARTWELL
2 129-6-4 MATa lysl nibl [cir+] C. HOLM
212 MATa karl-1 adel frpl-289 ura3-52 Ieu2-3,112 [&+I R. SCLAFANI
213 .\.IATa karl-1 leu2-3, I12 ura3-52 his7 lei++] R. SCLAFANI
34nrir" a nibl strain that is largely congenic to 34Nrir" This study
34nrir"a ;I MATa strain that is largely congenic to 34ncir" This study
4027-1-2 MATa ade2 ura? leu2 his? met2 Cyh' [cir"] 1.. HARTWELL
.'l)cyc-cyto # I * .LfATa nibl cyh2 [cir"] D . LIVINGSTON
2101-3-3* ,\.IATa adel nib1 [rir+] C. HOLM
2 103-6-2* ,VfATa adel nibl Cvh' [rir+] C. HOLM
54nrir" ,LId4Ta leu2 ade2 nibl Cyh' [cir"] This study
34Nrir" a iVIRI strain that is largely congenic to 54nrir" This study
Stl.;tins nlarked with an asterisk are S. carlsbergensis. T h e strain 54ncir" is the result of a cross between the S. carlshergensis strain 5l)cyc-
c y t o * I and the S. rermisiae strain 4027-1-2. T h e remaining strains are S. cermisiae.
detected by its selectable marker, and 2 p D N A was detected by colony hybridization. T h e presence of plasmids was ver- ified by Southern hybridization.
T h e percentage of abnormal cells was determined by growing a log phase culture in YEPD, concentrating the cells by centrifugation, sonicating and counting at least 200 cells under the phase contrast microscope, scoring them as either normal or abnormal. Cells scored as normal were small and ellipsoid-shaped (Figure 1, panel C ) , and cells scored as abnormal were large, often with an irregular or
elongated shape (Figure 1, panel B ) . Cells were also plated on selective (if applicable) and nonselective plates to deter- mine viability and the percentage of cells containing plas- mid.
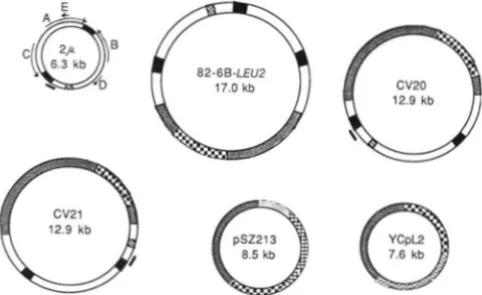
FIGCRE 2.- Pl;1smid D N A s . Plasmids ;we drawn proportional to s i x . C\'20 and C\'2 1 were provided by J. BROACH (BROACH and
HICKS 1980; BROACH et al. 1982). pS7.213 was provided by J. SZOSTAK (SZOSTAK and BLACKBURN 1982). Symbols are: 0 . 2 ~ D N A ;
..
inverted repeats of 2 p D S A ; R, the REP? region of 2 p D N A ;-, origin of replication of 2 p D N A ; +, ORFs of 2 p D N A ;
w.
.
pBR322 ( o r pUCI9) D N A ; Z, ARSI CEN4; ARSI; 0, and HIS3.contained 2 p DNA. These were then cured of the centro- mere plasmid (or CV21) by growth in non-selective medium. In the second method, yeast cells were transformed with the plasmid 82-GB-LEU2. After selection on leucine omission plates, transformants were transferred to nonselective plates. FLP-mediated recombination between the two copies of the inverted repeats adjacent to vector sequences gives rise to a small plasmid containing LEU2 but no ARS (which is quickly lost) and intact 2~ D N A (which is retained).
In some experiments, plasmids were introduced into the cycloheximide- resistant strains 54nci~') or 54Ncir" via crosses w i t h the karl strain 212 (containing 2 p D N A and a
test plasmid). Cytoductants were recovered in the following way. Four or five hours after initiating mating on a YEPD plate made with Noble agar (Difco), dumbbell-shaped zy- gotes were separated from the mating mixture by microma- nipulation and allowed to grow into colonies on YEPD plates. These colonies were then streaked onto YEPD plates containing cycloheximide or to leucine omission plates con- taining cycloheximide. Colonies were picked and tested for nuclear markers to verify that they were cvtoductants bear- ing the 54ncir" (or 54Ncir") nucleus. T h e test plasmid was
RESULTS
Requirement of elements other than 2c( DNA for the expression of the Nib- phenotype in a nib2 strain: Previous work (HOLM 1982a,b) could be inter- preted to mean that 2p DNA is necessary and suffi- cient to produce the Nib- phenotype in nib2 strains of yeast. As a first step in determining the portion of 2p DNA that is required for nibbled colony morphol- ogy in nib2 strains, congenic cir" S. cerevisiae strains, differing only at the NIB2 locus, were constructed (34Ncir" and 34ncir" see MATERIALS AND METHODS).
Contrary to expectation, 34ncir" did not display nib- bled colony morphology after introduction of 2p
DNA by transformation. In addition, cultures of some of these transformants were examined microscopically and found to lack the abnormal-looking cells charac- teristic of nib1 cir+ strains (Figure 1 ; HOLM 1982b). T h a t this strain had an authentic nib2 allele was con- firmed by demonstrating that nib1 was linked to RAD2
(data not shown). This result could be explained if the
2 p DNA received via transformation were defective and incapable of causing the Nib- phenotype. Alter- natively, expression of the Nib- phenotype might require elements other than nib2 and 2p DNA.
752 R. Sweeney and V. A. Zakian
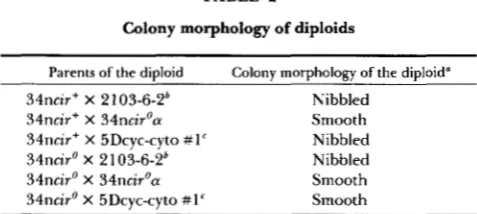
TABLE 2
Colony morphology of diploids
Parents of the diploid Colony morphology of the diploid"
34ncir+ X 2 103-6-2' Nibbled
34ncir+ X 34ncir'a Smooth
34ncir+ X 5Dcyc-cyto #1' Nibbled
34ncir' X 2 103-6-2' Nibbled
34ncir' X 34ncir'a Smooth
34ncir' X 5Dcyc-cyto # l C Smooth
Colony morphology was scored on Ymin or Yc plates incubated
2103-6-2 is a nib1 cir+ S. carlbergensis strain (see Table 1).
one day at 30" followed by 1 week at room temperature.
' 5Dcyc-cyto # 1 is a nibl cir' S. carlbergensis strain (see Table 1).
and a transformant of 34ncir' containing
211
DNA were crossed to several test strains, and the colony morphology of the diploids was observed (Table2).
T h e test strains included two S. carlsbergensis strains,2
103-6-2 (a nibl cir+ strain with nibbled colony mor- phology) and 5Dcyc-cyto #1 (a nibl cir' strain with smooth colony morphology), and one S. cerevisiae strain, 34ncir'a (a nibl cir' strain congenic to 34ncir0, differing only at the mating type locus). These data reconfirm that 2p DNA is necessary for the expression of the Nib- phenotype and demonstrate that the 2p DNA received by 34ncir' via transformation was com- petent to cause nibbled colony morphology in S. curls- bergensis/34n diploid strains (Table 2). Therefore, some other element provided by the S. carlsbergensis genome must be necessary to produce nibbled colony morphology. That this additional element(s) can be transferred to and maintained in S. cerevisiae is shown by the fact that HOLM (1982a) was able to construct nibl cir+ S. cerevisiae strains that displayed nibbled colony morphology.The additional genetic element(s) does not behave as a chromosomal gene(s): T o determine the nature of the additional element(s) necessary for the expres- sion of the Nib- phenotype, its meiotic segregation was monitored. A S. cerevisiae nibl cir+ strain (2129- 6-4; HOLM 1982a) that exhibits nibbled colony mor- phology was crossed to 34ncir0, which lacks the addi- tional element(s). The 2p DNA present in 2 129-6-4 should segregate to all four spores (LIVINGSTON
1976), an expectation confirmed in this cross by Southern hybridizations on DNA from progeny from individual spores (data not shown). All spores should also carry the nib1 allele, an expectation confirmed by crosses to nibl tester strains as described in MATERIALS AND METHODS. If the additional element necessary for the expression of the Nib- phenotype is a single, dominant chromosomal locus, it is expected to segre- gate 2:2 in meiosis, producing tetrads consisting of two spores with nibbled and two spores with smooth colony morphology. Likewise, if 34ncir' bears a reces- sive chromosomal element that suppresses the Nib- phenotype or if 34nci:.' is a chromosome VZZ disome (shown previously to :uppress the Nib- phenotype;
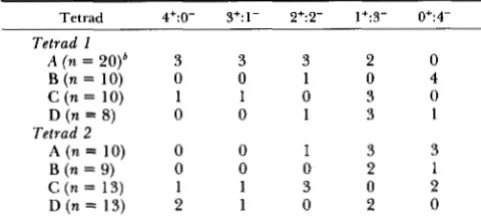
TABLE 3
Segregation of additional element required for nibbled colony morphology
No. viable spores per tetrad
Segregation of colony morphology"
4 ( n ~ 2 3 ) ~ 4+:0-
""-
3+:1- 2+:2- 1+:3- 0+:4-0 0 6 1 1 6
3 (n = 9) 3+:0- 2+:1- 1+:2- 0+:3-
""
0 1 2 6
2 (n = 22) 2+:0- 1+:1- 0+:2-
1 (n = 4) 1 + 1-
0 (n = 3)
" -
2 4 16
-
2 - 2
Colony morphology of each spore from a cross between 2 129- 6-4 and 34ncir' was examined on YEPD plates incubated for one week at 30". Although individual colonies could be scored as nibbled (-) or smooth (+), the extent of nibbling varied among the different spore clones.
n is the number of tetrads having said number of viable spores.
HOLM 1982a), nibbled colony morphology should seg- regate 2:2. If the additional element is one (or more) extrachromosomal element(s), one expects that all (or most) spores will exhibit nibbled colony morphology (since the diploid was homozygous for nibl ). Most known extrachromosomal elements like 2p DNA, double-stranded RNAs, and mitochondria segregate 4:O in meiosis (LIVINGSTON 1976; SOMERS AND BEVAN 1969; GILLHAM 1974). However, URE3 (an extra- chromosomal element that enables a ura2 mutant strain to grow in the presence of ureidosuccinic acid plus glutamic acid) is retained in only a subset of the meiotic progeny in a URE3 X wild-type cross (33- 87%; LACROUTE 1971; AIGLE AND LACROUTE 1975).
Yeast Extraachromosomal Elements 753
TABLE 4
Colony morphology of spores from backcrosses of Nib- spores"
Tetrad 4+:0- 3+:1- 2+:2- 1+:3- 0+:4- Tetrad 1
A (n = 20)6 3 3 3 2 0
B ( n = 10) 0 0 1 0 4
D (n = 8) 0 0 1 3 1
A (n = 10) 0 0 1 3 3
B (n = 9) 0 0 0 2 1
C (n = 13) 1 1 3 0 2
D (n = 13) 2 1 0 2 0
c
(n = 10) 1 1 0 3 0Tetrad 2
a Each of 4 spores from two tetrads derived from the cross of
2 129-6-4 X 34ncir' (Table 3) in which each spore produced a Nib- colony was crossed to 34ncir'. Colony morphology of each of the four members of tetrads containing four variable spores was ex- amined on YEPD piates incubated for 1 week at 30".
'
n is the number of tetrads dissected.fewer than four viable spores. For tetrads with three viable spores, some of the anticipated values (three 0 Nib+:3 Nib- tetrads; four 1 Nib+:2 Nib- tetrads) are different from the actual values (six 0 Nib+:3 Nib- tetrads; two 1 Nib+:2 Nib- tetrads), and, therefore, spore death is probably not random. If we assume that Nib- spores are more likely to die than Nib+ spores (a reasonable hypothesis given their reduced division potential), the data suggest that the 0 Nib+:4 Nib- class is underrepresented in tetrads with four viable spores. Even if we assume that all spore inviability is due to death of Nib+ spores and all tetrads with fewer than four viable spores are taken into consideration, the 2+:2- class of tetrads would equal only about 40% of the total. Therefore, the additional elements(s) required to express nibbled colony morphology does not segregate 2:2 in the majority of tetrads and is not a single chromosomal locus.
T h e data presented in Table 3 can be explained if the additional element is an extrachromosomal ele- ment that segregates to most, but not all, spores in meiosis. Alternatively, the additional element could be either two, unlinked, dominant chromosomal ele- ments, either of which can cause the Nib- phenotype, or two, unlinked, recessive suppressors, both of which are needed to suppress the Nib- phenotype. T o ad- dress these latter possibilities, each spore from two tetrads in which all four spores had nibbled colony morphology was recrossed to 34ncir' (Table 4). If the extra elements are two chromosomal elements as pos- tulated above, then nibbled colony morphology should segregate 2:2 in each cross. However, in three of the eight crosses there were no tetrads in which nibbled colony morphology segregated 2:2 (in the rest of the crosses, nibbled colony morphology segregated 2:2 in 14-43% of the tetrads; Table 4). These results eliminate the possibility that the additional elements are either two, unlinked, dominant chromosomal ele- ments, either of which can cause the Nib- phenotype, or two, unlinked, recessive suppressors, both of which
are needed to suppress the Nib- phenotype. There- fore, the fact that a majority
(78%)
of the spores from all tetrads in the cross 21 29-6-4 X 34ncir' had nib- bled colony morphology (Table 3) is best explained by the hypothesis that the additional factor is an extrachromosomal element that, like URE3, segre- gates to most, but not all, spores in meiosis.At least two of the 2 p ORFs are required for the
Nib- phenotype: T h e strain 54ncir' was the result of a single cross between the S. carlsbergensis nibl cir' strain 5Dcyc-cyto #1 and the S. cerevisiae strain 4027-
1-2. When 2 p DNA is introduced into 54ncir' by transformation, 80% of the transformants display nib- bled colony morphology. Therefore, 54ncir' must carry all element(s) other than 2p DNA that are nec- essary for expression of the Nib- phenotype.
In order to determine the portions of
2p
DNA that are required for production of the Nib- phenotype, recombinant DNA plasmids containing 2p DNA were introduced by transformation into 54ncir'. Two plas- mids with single interruptions in otherwise full length2p
DNA were used: CV20, which is interrupted inFLP or the A ORF, and CV21, which is interrupted
in the D ORF
UAYARAM,
LI and BROACH 1983). Strains produced by transformation of 54ncir' by either CV20 or CV2 1 had smooth colony morphology (data not shown). This result means either that both the A and D ORFs are necessary for the expression of the Nib- phenotype or that some property dis- played only by intact 2p DNA is required for the expression of the Nib- phenotype. Alternatively, if only the subset of cells carrying the plasmid have a reduced division potential, the percentage of such cells might not be sufficient to ensure that the colony itself displays a nibbled morphology. Because CV20 and CV21 are lost at rates of about 1 O-' per cell per generation UAYARAM, LI and BROACH 1983), about 30% of the cells in a selectively grown colony do not contain plasmid DNA.754 R. Sweeney and V. A. Zakian
TABLE 5
Percentage of nibl transformants having a high percentage of abnormal cells
Percentage of isolates Average percentage having > l o %
Strain" abnormal cells'
of plasmid- bearing cells'
54ncir0/2p 87 (21) NA
54nciro/CV20 13 ( 8 ) 69
54nciro/CV21 0 (7) 71
54nciro/YCpL2 13 ( 8 ) 69
54ncir0 0 ( 8 ) NA
a 54ncir' was transformed with either CV20, CV2 1 , YCpL2 or
2 p DNA.
* This percentage is not corrected for the percentage of plasmid- bearing cells in the culture. The number in parentheses indicates the number of isolates examined.
' T h e percentage of plasmid-bearing cells was determined by simultaneous plating on selective (Yc-leucine) and nonselective (YEPD) plates. N A , not applicable.
versely with viability (data not shown). T h e percent- age of abnormal cells therefore constitutes an alter- native way to observe the reduced division potential characteristic of cells that display a nibbled colony morphology. We will hereafter refer to strains that have greater than or equal to 10% abnormal cells as having a high percentage of abnormal cells and to strains having fewer than 10% abnormal cells as hav- ing a low percentage of abnormal cells. Using this criterion, only 2p DNA, and not CV20 or CV21, caused a high percentage of abnormal cells in the majority of the isolates, when introduced by transfor- mation into the strain 54ncir' (Table 5). Again, these results support the earlier conclusion that the Nib- phenotype requires, at the least, the A and the D ORFs or possibly all of 2p DNA. Since the A and D ORFs are both required for high copy number of 2p DNA (VOLKERT and BROACH 1986; REYNOLDS, MUR-
R A Y and SZOSTAK 1987; MURRAY et al. 1987), these results may simply reflect the dependence of the Nib- phenotype on high copy number of 2p DNA.
Abnormal cell morphology and reduced division potential but not nibbled colony morphology can be conferred on nib2 cells by elements other than 2 p DNA: Because nibbled colony morphology had ini- tially been observed by introducing 2p DNA into a nib1 cirO strain via a cross with a hurl cir+ strain (HOLM
1982a), various plasmids were transferred into 54ncir0, or a congenic NIB1 strain, 54Ncir' via crosses with a hurl strain (212) that contained 2p DNA plus a test plasmid. Cytoductants containing 2~ DNA, the test plasmid or no plasmid were isolated. Most (95%) nibl cytoductants that received 2p DNA had a high percentage of abnormal cells (Table 6). T h e majority of these cytoductants (66%) also exhibited nibbled colony morphology (Table 6). Most nibl cytoductants that received one of the test plasmids, YCpL2 (a circular centromere plasmid), pSZ2 13 (an acentric circular plasmid), CV20 or CV2 1, also had high per- centages of abnormal cells (Table 6). Surprisingly,
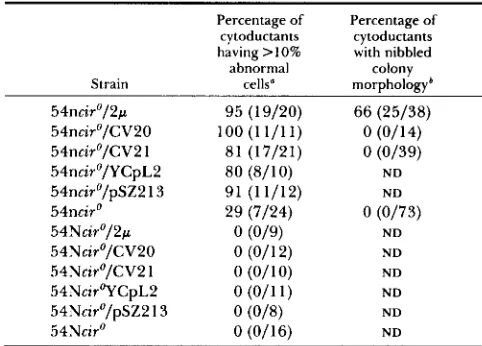
TABLE 6
Percentage of cytoductants having a high percentage of abnormal cells and nibbled colony morphology
Percentage of Percentage of cytoductants cytoductants having 110% with nibbled colony morphology'
54ncir0/2p 95 (1 9/20) 66 (25/38)
54nciro/CV20 l O O ( l l / l l ) 0 (0/14)
54nciro/CV21 81 (17121) 0 (0/39)
54nciro/YCpL2 80 (8/10) ND
54nciro/pSZ2 13 91 (1 1/12) ND
54ncir0 29 (7/24) 0 W73)
54Nciro/2p 0 (019) ND
54Nciro/CV20 0 (011 2) N D
54Nciro/CV2 1 0 (01 10) N D
54NcirOYCpL2 0 (0/11) N D
54Nciro/pSZ21 3 0 ( o m ND
54Ncir0 0 (0/16) ND
abnormal Strain cells"
Percentage of cytoductants examined in each class that had greater than or equal to 10% abnormal cells. The value in paren- theses indicates the number of cytoductants examined having greater than 10% abnormal cells over the total number of cytoduc- tants examined. The high percentage of abnormal cells does not require the presence of the recombinant DNA plasmid (see RE- SULTS).
'
Colony morphology was examined after one week of growth on YEPD plates at 30". The percentage indicates the percentage of the cytoductants examined that had nibbled colony morphology. The value in parentheses indicates the number of cytoductants having nibbled colony morphology over the total number exam- ined.some of the plasmidless nibl cytoductants (29%) had high percentages of abnormal cells, although they did not have nibbled colony morphology (Table 6). This result contrasts with the parental nib1 cir'" strain: out of eight cultures grown from independent colonies, none had high percentages of abnormal cells (Table 5). As expected, all NIB1 cytoductants from the con- genic strain, whether or not they received plasmid DNA, had low percentages of abnormal cells (Table
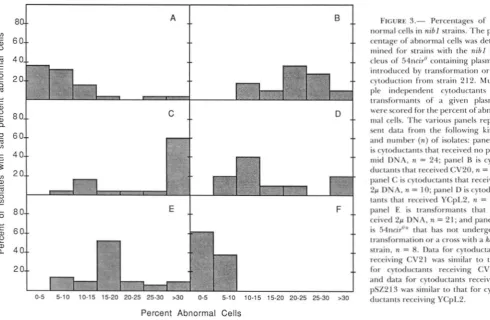
Although most nib1 cytoductants that received either 2p DNA or one of the test plasmids had high percentages of abnormal cells, only cytoductants con- taining 2p DNA had nibbled colony morphology (Table 6). There was, in addition, a quantitative dif- ference in the percentage of abnormal cells: most cytoductants containing 2p DNA had higher percent- ages of abnormal cells than did cytoductants contain- ing either CV20, CV21, pSZ213 or YCpL2 or than cells carrying 2p DNA that had been introduced by transformation (Figure 3). These data indicate that transformants or, especially, cytoductants that have received 2p DNA have a more severe phenotype, including both high percentages of abnormal cells and nibbled colony morphology, than do cytoductants that have received CV20, CV2 1, pSZ213 or YCpL2, which exhibit high percentages of abnormal cells but have smooth colony morphology. T h e pSZ213 plasmid is
Yeast Extraachromosomal Elements
Percent Abnormal Cells
containing pSZ2 13 that had high percentages (>25%) of abnormal cells had low percentages ( 5 2 0 % ) of plasmid-bearing cells. Moreover, when cytoductants containing CV20 and CV21 are cured of these plas- mids by growth in non-selective medium, six out of six isolates had virtually the same percentage of ab- normal cells after curing. These data indicate that the presence of plasmid D N A is not necessary to cause a high percentage of abnormal cells. This result suggests that another extrachromosomal factor that is readily transferred to the recipient cell in a cross with a Karl
strain may be responsible for the high percentage of abnormal cells. We hypothesize that this other ele- ment is transferred at relatively high frequency when another plasmid is transferred but can also be trans- ferred alone (thereby accounting for the high per- centage of abnormal cells in some of the cytoductants that received n o plasmid). In addition, the higher
percentages of abnormal cells seen in nib1 cytoduc- tants that contain 2p D N A as compared to transform- ants that contain 2p D N A may be accounted for by the presence of this other element in cytoductants but not in transformants.
DISCUSSION
O u r results (Table 2) demonstrate that at least o n e dominant genetic element in addition to 2p D N A is required to cause nibbled colony morphology in a
nibl strain. A variety of extrachromosomal genetic
FIGURE 3.- Percentages o f ab- normal cells in nibl strains. T h e per- centage of abnormal cells was deter- mined for strains with the nib1 nu- cleus o f 54ncir'' containing plasmids introduced by transformation or by cytotluction from strain 212. Multi- ple independent cytociuctants or transformants o f ;I given plasmid
were scored for the percent o f abnor- mal cells. T h e various panels repre- sent data from the following kinds and number (n) o f isolates: panel A
is cytoductants that received n o plas- mid DNA, n = 24: panel R is cyto- ductants that received CV20. n = 1 I ; panel C is cytoductants that received 2p D N A . n = 10: panel D is cytoduc- tants that received YCpI.2, n = 1 0 ;
panel I.: is transformants that re- ceived 2p D N A , n = 21; and panel F
is ~411Ci~"" that has not undergone transformation or ;I cross with a k a r f
strain. n = 8. Data for cytoductants receiving CV21 w a s similar to that for cytoductants receiving CV20.
and data for cytoductants receiving pSZ213 was similar to that for cyto- ductants receiving YCpL2.
elements has been observed in yeast. Some of these have been characterized biochemically, for example, 2p D N A (HARTLEY and DONELSON 1980), various forms of double- stranded RNAs involved in killer function (WICKNER 1986), and mitochondrial D N A (DUJON 198 1). Other extrachromosomal elements like the t,b factor (TUITE et al. 1982) and [URES] (LAC- ROUTE 197 1 ; AICLE and LACROUTE 1975) have been characterized genetically, but their biochemical basis is unknown. Although the meiotic segregation (Table 3) of the additional element needed to manifest nib- bled colony morphology is most easily explained by its being an extrachromosomal element that, like [URES], shows irregular meiotic segregation (LAC- ROUTE 197 1; AICLE and LACROUTE 1975), its exact identity was not determined.
T h e r e is a likely explanation for why the require- ment for the additional extrachromosomal element was not noted in the original work with this system (HOLM 1982a,b). T h e nibl allele was crossed into a
cir+ S. cerevisiae strain. Only those segregants display- ing nibbled colony morphology were chosen for fur- ther analysis; i.e., there was selection for segregants that maintain the additional element. Consistent with this interpretation, in early crosses the Nib- pheno- type did not segregate 2:2 in these cir+ strains (HOLM
1982a).
756 R. S w e e n e y and V. A. Zakian
phology, we have constructed a nibl strain (54ncir')
that has smooth colony morphology and a low per- centage of abnormal cells when it contains no plasmid DNA and nibbled colony morphology and a high percentage of abnormal cells when it receives 2p DNA via transformation. Therefore, this strain must con- tain all elements other than 2p DNA necessary to express the Nib- phenotype. Two chimeric plasmids (CV20 and CV21) containing the full length of 2p DNA, interrupted within either the A or the D ORF, do not induce nibbled colony morphology or a high percentage of abnormal cells when they are intro- duced into 54ncir' by transformation. Therefore, the A and the D ORFs are simultaneously required for a reduction in division potential. Since the gene product of the D ORF is a positive regulator of transcription from the A ORF (MURRAY et al. 1987), and the gene product of the A ORF is necessary for the amplifica- tion of 2p DNA copy number (REYNOLDS, MURRAY and SZOSTAK 1987; VOLKERT and BROACH 1986), this result is consistent with the idea that high copy num- ber of 2p DNA is necessary to cause the Nib- pheno- type. Alternatively, some property unique to intact 2p DNA, such as the ability to attach to a specific site within the nucleus, is required.
When 2p DNA is introduced into 54ncir' via a cross with a Karl strain, its presence is correlated with a high percentage of abnormal cells. Other plasmids, such as CV20, CV2 1, YCpL2 or pSZ2 13, can also be transferred into 54ncir' via a cross with a Karl strain. Cytoductants often show a high percentage of abnor- mal cells that correlates with a reduced division poten- tial. This high percentage of abnormal cells is not dependent on the presence of plasmid, since isolates that have been cured of plasmid still have a high percentage of abnormal cells. Therefore, it seems likely that an extrachromosomal element other than the selected plasmid is transferred during the cross with the Karl strain and that this element can itself cause a high percentage of abnormal cells in nibl
strains. T h e fact that a significant percentage (29%) of cytoductants that received no plasmid DNA dis- played a high percentage of abnormal cells (Table 6) suggests that this element is also transferred in the absence of plasmid transfer.
T h e severity of the Nib- phenotype varies both from strain to strain and among different colony isolates from the same strain. For example, different colonies from the same clone differ in the severity of the phenotype (Figure 1, panel A; HOLM 1982b) as do different transformants or cytoductants produced in the same experiment. Also, 2p transformants have lower percentages of abnormal cells than do strains that receive 2p DNA by cytoduction (Figure 3). These differences may be due to the different amounts of the unidentified extrachromosomal element(s) in in- dividual cells or in transformants vs. cytoductants. It
is tempting to speculate that this element(s) may be the same as the extrachromosomal element required for nibbled colony morphology that we have charac- terized by tetrad analysis.
In spite of the fact that cytoductants that have received CV20, CV21, pSZ213 or YCpL2 have high percentages of abnormal cells, only cytoductants that have received 2p DNA display nibbled colony mor- phology. This observation indicates that nibbled col- ony morphology and a high percentage of abnormal cells are not identical phenotypes, although both phe- notypes correlate with a reduced division potential. As demonstrated by macromanipulation (HOLM 1982b), some cells that appear abnormal are capable of dividing, and these might give rise to a smooth rather than a nibbled colony morphology.
In summary, in order to display nibbled colony morphology, nibl strains must contain both 2p DNA and at least one, as yet unidentified, extrachromo- somal element. Expression of the Nib- phenotype requires the presence of at least the A and D ORFs of 2p DNA, both of which are involved in the control of
2 p DNA copy number. A less severe reduction in division potential, reflected in a high percentage of abnormal cells, can be detected in nibl cir' strains after the introduction of extrachromosomal elements (other than 2p DNA) by crosses with hurl strains. These results might indicate that cells carrying the
nibl allele have some generalized cellular defect that makes them sensitive to the presence of a number of different extrachromosomal elements, of which 2p DNA is only one. Since the reduced division potential displayed by nib1 strains is not specific for 2p DNA, this system is unlikely to yield information important for an understanding of 2p copy number control.
This work was supported by National Institute of Health Train- ing Grant AGO0057 awarded to the Pathology Department of the University o f Washington which supported R.S. as a postdoctoral trainee and by grants to V.A.Z. from the National Institutes of Health and the American Cancer Society. We thank A. TRAYLOR for technical assistance, B. BREWER, M . CONRAD and C. HOLM for their critical reading of the manuscript, L. HARTWELL for helpful discussion of the data, and J. Bryant for preparation o f the manu- script.
L I T E R A T U R E C I T E D
AIGLE, M . , and F. LACROUTE, 1975 Genetical aspects of [ U R E 3 ] ,
a nonmitochondrial, cytoplasmically inherited mutation in yeast. Mol. Gen. Genet. 1 3 6 327-335.
ANDREADIS, A., Y.-P. Hsu, G. B. KOHLHAW and P. SCHIMMEL, 1982 Nucleotide sequence o f yeast LEU2 shows 5"noncoding
region has sequences cognate to leucine. Cell 31: 319-325. BEGGS, J. D., 1978 Transformation of yeast by a replicating hybrid
plasmid. Nature 275: 104-108.
BREWER, B., and W . L. FANGMAN, 1987 The localization of rep- lication origins on ARS plasmids in S. cerevisiae. Cell 51: 463- 471.
757
BROACH, J. R., and J. B. HICKS, 1980 Replication and recombi- nation functions associated with the yeast plasmid, 2p circle. Cell 21: 501-508.
CLARK-WALKER, G. D., and G. L. G. MIKLOS, 1974 Localization and quantification of circular DNA in yeast. Eur. J. Biochem.
CONDE, J., and G. R. FINK, 1976 A mutant of Saccharomyces cerevisiae defective for nuclear fusion. Proc. Natl. Acad. Sci.
DAVIS, R. W., M. THOMAS, J. CAMERON, T. P. ST. JOHN, S. SCHERER and R. A.PADGETT, 1980 Rapid DNA isolations for enzymatic and hybridization analysis. Methods Enzymol. 65: 404-4 1 1. DOBSON, M. J., FUTCHER, A. B. and B. S. Cox, 1980 Loss of 2pm
DNA from Saccharomyces cerevisiae transformed with the chi- maeric plasmid pJDB219. Curr. Genet. 2: 201-205.
DUJON, B., 1981 Mitochondrial genetics and functions, pp. 505- 635 in The Molecular Biology of the Yeast Saccharomyces, edited by J. N. STRATHERN, E. W. JONES and J. R. BROACH. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. DUTCHER, S. K., 1981 Internuclear transfer of genetic informa-
tion in k a r l - l / K A R I heterokaryons in Saccharomyces cereuisiae. Mol. Cell. Biol. 1: 245-253.
FILETICI, P., N. JUNAKOVIC and P. BALLARIO, 1985 Rapid alkaline preparation for yeast circular covalently closed DNA mole- cules. Curr. Genet. 9 123-1 26.
FUTCHER, A. B., and B. S. Cox, 1983 Maintenance of 2-pm circle plasmid in populations of Saccharomyces cereuisiae. J. Bacteriol.
FUTCHER, H. B., and B. S. COX, 1984 Copy number and the stability of 2-pm circle-based artificial plasmids of Saccharomyces cerevisiae. J. Bacteriol. 157: 283-290.
GILHAM, N. W., 1974 Genetic analysis of the chloroplast and mitochondrial genomes. Annu. Rev. Genet. 8: 347-391. HARTLEY, J. L., and J. E. DONELSON, 1980 Nucleotide sequence
of the yeast plasmid. Nature 2 8 6 860-864.
HINNEN, A,, J. B. HICKS and G. R.FINK, 1978 Transformation of yeast. Proc. Natl. Acad. Sci. USA 75: 1929-1933.
HOLM, C., 1982a Sensitivity to the yeast plasmid 2p DNA is conferred by the nuclear allele n i b l . Mol. Cell. Biol. 2: 985- 992.
HOLM,^., 1982b Clonal lethality caused by the yeast plasmid 2p DNA. Cell 2 9 585-594.
HUBERMAN, J. A., L. D. SPOTILA, K. A. NAWOTKA, S. M. EL- ASSOULI and L. R. DAVIS, 1987 The in vivo replication of the yeast 2pn1 plasmid. Cell 51: 473-481.
ITO, H., Y. FUKUD, K. MURATA and A. KIMURA, 1983 Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 153: 163-168.
JAYARAM, M., Y.-Y. LI and J. R. BROACH, 1983 The yeast plasmid
Zp circle encodes components required for its high copy prop- agation. Cell 34: 95-104.
JAYARAM, M., A. SUTTON and J. R. BROACH, 1985 Properties of REP3:a cis-acting locus required for stable propagation of the Saccharomyces cerevisiae plasmid 2 pm circle. Mol. Cell. Biol. 5:
KIKUCHI, Y., 1983 Yeast plasmid requires a cis-acting locus and two plasmid proteins for its stable maintenance. Cell 35: 487- 493.
LACROUTE, F., 1971 Non-Mendelian mutation allowing ureido-
LIVINGSTON, D. M., 1976 Inheritance of the 2pm DNA plasmid
41: 359-365.
USA 73: 3651-3655.
154: 612-622.
2466-2475.
succinic acid uptake in yeast. J. Bacteriol. 106: 519-522.
from Saccharomyces. Genetics 86: 73-84.
LIVINGSTON, D. M., and D. M. KUPFER, 1977 Control of Saccha- romyces cerevisiae 2pm DNA replication by cell division cycle genes that control nuclear DNA replication. J. Mol. Biol. 116:
MANIATIS, T., E. F. FRITSCH and J. SAMBROOK, 1982 Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
MORTIMER, R. K., and D. C. HAWTHORNE, 1966 Genetic mapping in Saccharomyces. Genetics 53: 165-173.
MURRAY, J. A., H. SCARPA, M. Ross1 and G. CFSARENI 1987 Antagonistic controls regulate copy number of the yeast 2p plasmid. EMBO J. 6 4205-4212.
NEWLON, C. S., R. J. DEVENISH, P. A. SUCI and C. J.ROFFIS, 1981 Replication origins used in vivo in yeast, pp. 501-516 in Structure and DNA Protein Interactions of Replication Origins, edited by D. S. RAY and C. F. FOX (ICN-UCLA Symposia on Molecular and Cellular Biology, Vol. XXI). Academic Press, New York.
PETFS, T . D., and D. H. WILLIAMSON, 1975 Replicating circular DNA molecules in yeast. Cell 4: 249-253.
PLUTA, A. F., G. M. DANI, B. B. SPEAR and V. A. ZAKIAN, 1984 Elaboration of telomeres in yeast: recognition and mod- ification of termini from macronuclear DNA. Proc. Natl. Acad. Sci. USA 81: 1475-1479.
REYNOLDS, A. E., A. W. MURRAY and J. W. SZOSTAK, 1987 Roles of the 2pm gene products in stable maintenace of the 2pm plasmid of Saccharomyces cerevisiae. Mol. Cell. Biol. 7: 3566- 3573.
SOMERS, J. M., and E. A. BEVAN, 1969 The inheritance of the killer character in yeast. Genet. Res. 13: 71-83.
SOUTHERN, E. M., 1975 Detection of specific sequence among DNA fragments separated by gel electrophoresis. J. Mol. Biol.
STINCHCOMB, D. T., C. MANN and R. W. DAVIS, 1982 Centro- meric DNA from Saccharomyces cereuisiae. J. Mol. Biol. 1 5 8
SZOSTAK, J. W., and E. H. BLACKBURN, 1982 Cloning yeast telo- meres on linear plasmid vectors. Cell 29: 245-255.
TUITE, M. F., P. M. LUND, A. B. FUTCHER, M. J. DOBSON, B. S.
Cox and C. S. MCLAUGHLIN, 1982 Relationship of the [ p s i ]
factor with other plasmids of Saccharomyces cerevisiae. Plasmid
8: 103-1 11.
VOLKERT, F. C., and J. R. BROACH, 1986 Site-specific recombi- nation promotes plasmid amplification in yeast. Cell 46: 541- 550.
WAHL, G. M., M. STERN and G. R. STARK, 1979 Efficient transfer of large DNA fragments from agarose gels to diazobenzyloxy- methyl-paper and rapid hybridization by using dextran sulfate. Proc. Natl. Acad. Sci. USA 7 6 3683-3687.
WICKNER, R. B., 1986 Double-stranded RNA replication in yeast: the killer system. Annu. Rev. Biochem. 55: 373-395.
YANISCH-PERRON, C., J. VIEIRA and J. MESSING, 1985 Improved phage cloning vectors and host strains: nucleotide sequences of M13mp18 and pUCl9 vectors. Gene 33: 103-1 19.
ZAKIAN, V. A., B. J. BREWER and W. L. FANGMAN, 1979 Repli- cation of each copy of the yeast 2 micron DNA plasmid occurs during the S phase. Cell 17: 923-934.
ZAKIAN, V. A., and J. F. Scorn, 1982 Construction, replication and chromatin structure of T R P l RI circle, a multiple-copy synthetic plasmid derived from Saccharomyces cereuisiae chro- mosomal DNA. Mol. Cell. Biol. 2: 221-232.
249-260.
98: 503-517.
157-179.