INTERCHROMOSOMAL EFFECTS AND THE RELATION BETWEEN CROSSING-OVER AND NONDISJUNCTION1
PAUL ROBERTS
Department of Zmlogy, The University of Chicago, Chicago, Illinois2
Received July 5 , 1952
HE ability of the same chromosomal rearrangements to modify both crossing- and segregation of nonhomologous chromosomes in the female of Drosophila melanogaster permits a study of the relation of these two aspects of chromosome behavior under controlled conditions. A brief consideration of previ- ous investigations of these “interchromosomal effects” follows.
STURTEVANT (1919) first observed that a heterozygous inversion on the third chromosome increased crossing-over on chromosome 2. Since then, many investi- gators have shown that heterozygous inversions in any pair of major chromo- somes increase crossing-over in the other pairs. (A review of earlier work on this phenomenon has been published by SCHULTZ and REDFIELD 1951.) The cause of this effect remains unknown.
STURTEVANT’S report (MORGAN and STURTEVANT 194%) that disjunction of
x
chromosomes is also subject to modification by autosomal structural heterozy- gosity, an interchromosomal effect on segregation, introduced a paradox. His observation that heterozygous autosomal inversions increase the rate of primary nondisjunction of X chromosomes if they are also heterozygaus for an inversion seemed to conflict with the findings of BRIDGES (1916) and STURTEVANT and
BEADLE
(1936) that nondisjunctional X chromosomes are derived almost ex- clusively from no-exchange tetrads. Since heterozygous autosomal inversions in- crease crossing-over between X chromosomes, the proportion of no-exchange tetrads and therefore the amount of nondisjunction of the X chromosomes should be decreased.An attempt to resolve this contradiction was made by COOPER, ZIMMERING and
KRIVSHENKO
( 1955) who proposed that interchromosomal effects on segregation were caused by nonhomologous pairing between structurally heterozygous auto- somes and X chromosomes. Dissociation of nonhomologous partners would lead to not only X chromosome “nondisjunction” but also to autosomal “nondisjunc- tion” resulting in dominant lethals. This, they state, would produce selective elimination of low rank tetrads giving an apparent increase in multiple ex- changes, an “interchromosomal effect” on crossing-over.1From a dissertation submitted to the University of Chicago in partial fulfillment of the requirements for the degree of Doctor of Philosophy. This investigation was supported by United States Public Health Service Grant RG-7428 and a Traineeship under Public Health Service Training Grant 2G-150 from the National Institutes of Health.
2 Present address: Biology Division, Oak Ridge National Laboratory, Oak Ridge, Tennessee.
1692 P A U L ROBERTS
It has not been possible to verify the occurrence of nonhomologous pairing by cytological study of Drosophila oocytes. However, considerable genetic evidence for nonhomologods pairing comes from data showing highly nonrandom segre- gations of nonhomologous chromosomes. I n this type of experiment, pairing between nonhomologues may be induced in high frequencies by the presence in the genome of two or more nonhomologous chromosomes in which homologous pairing has been disturbed by a structural rearranganent or by addition of an extra element. Thus preferential Y-4 segregation was demonstrated by GRELL
(1957, 1959b) i n females carrying a Y and a heterozygous 3 - 4 translocation, and preferential X-2 segregation was shown by FORBES (1960) in females heterozygous for inversions on the X and second chromosome plus a heterozy- gous 2-3 translocation.
Although COOPER, ZIMMERING and KRIVSHENKO’S hypothesis that the in- creased nondisjunction observed under conditions of structural heterozygosity results from nonhomologous pairing has considerable genetic evidence to support it, their explanation of the increase in crossing-over that accompanies structural heterozygosity is in doubt. REDFIELD (1957) found that egg mortality (a direct consequence of the postulated elimination of no-exchange tetrads) in females carrying the heterozygous Curly and Payne inversions is too low to account for observed recombination increase between the X’s, and she has pointed out that the changed proportions of higher rank strands could not be accounted for by low rank elimination. Since COOPER et al. have only claimed that their hypothesis would partially account for an interchromosomal effect on crossing-over, it would be desirable to know when, and how much elimination of noncrossover tetrads contributes to changes in crossing-over under conditions of structural heterozygosity.
I n the experiments reported here, simultaneous measurements of X chromo- some crossing-over and nondisjunction were made using females carrying various combinations of heterozygous inversions. Evidence will be presented leading to a n interpretation of interchromosomal effects consistent with the recent hy- pothesis of GRELL (1962) that meiosis in Drosophila females is separable into two sequential phases: one concerned with crossing-over, the other with disjunc- tion. The results suggest that the primary interchromosomal effect of heterozy- gous inversions is to produce a real increase in crossing-over at a time when pairing is only between homologous chromosomes. This may be followed by nonhomologous pairing of noncrossover chromosomes. The extent to which non- homologous pairing occurs subsequent to exchange depends, roughly, on the proportion of no-exchange tetrads present.
MATERIALS A N D METHODS
I n the first series of crosses using different X-inversion heterozygotes, X
chromosome crossing-over and nondisjunction were studied in the presence of
one or two autosomal inversions, Zns(2L
+
2R)Cy, Cy, and Zns(3L+
3 R )CROSSING-OVER AND NONDISJUNCTION 1693 Curly (wings) while the third chromosome inversion carries the identifying dominant Deformed (eyes). To prevent possible loss of inversions through crossing-over, the X (except for ClB) and the two autosomal inversions were introduced into males which were then crossed to females carrying a series of X chromosome markers. Among the progeny of this cross were sisters carrying the appropriate inversion combinations which were then crossed to Bar (eye) males. This provided environmental uniformity during development and some control of genetic background among crosses to be compared involving the same X inver- sion. However, it should be noted that genetic backgrounds of flies arising from crosses of the different inversion stocks are different to an unknown degree; for this reason sister segregants from the original cross were used as controls.
Therefore, each experiment testing a different X inversion consisted of four crosses which were run simultaneously. Such an experiment will be designated by a letter of the alphabet with subscripts for the purpose of identification in Tables 2-4 and for subsequent reference. Thus, for example:
A x s ~ 7 = I n ( 1 ) ~ ~ 7 / ~ ~ e c c v c t u g f ~ P X B 8 8
Ax, = In(l)sc7/sc ec cu ct U g f; Cy/+
o
0 x B 8 8Ax, = In(l)sc7/sc ec cu ct U g f ; Payne, Dfd/+ ? 0 x B 8 8
Axcp = Zn(l)sc7/sc ec cu ct U g f; Cy/+; Payne, Dfd/+ 0 0 x B 8 8
Three different series of X chrommosome markers were used: (1) sc ec cu ct U g
f with inversions sc7, B y 2 , AB, and AM; (2) y2 ec cu U f car with inversions sc8,
s e 6 O - I 4 , C1B and Ins (1)dl-49, B y l ; (3) ec cu U
f
car with inversions y4 and dl-49,y f.". These markers have the following loci on the standard map: y2, yellow2, 0.0; sc, scute, O.O+; ec, echinus, 5.5; C U , crossveinless, 13.7; ct, cut, 20.0; U , ver-
milion, 33.0; g, garnet, 44.4; f, forked, 56.7; B, Ear, 57.0; car, carnation, 62.5
(BRIDGES and BREHME 1944.). Table 1 lists the different X inversions tested, giving the cytological and, where known, the genetic positions of the left and right breaks (BRIDGES and BREHME 1944).
A second series of crosses (Table 5 ) was made utilizing a n autosomal translo- cation that affects the exchange and nondisjunction of X chromosomes heterozy- gous for an inversion. Two X chromosome inversions which gave high rates of
nondisjunction and yet permitted considerable crossing-over outside the inver- sion were used: I n ( 1 )AM permitted measurement of crossing-over distally and I n ( 1 ) dl-4.9 near the centromere. Each of these inversions was tested in females carrying the translocation T( 2;3) X a ( X a = Xasta, scalloped wings) in addition to the Curly inversion. Sisters of the above females which lacked either the Curly inversion or the translocation were run as controls to measure crossing-over when nonhomologous pairing is reduced or absent. Thus the crosses involving AM were:
MACXa = In (1
1
AM/sc ec cu ct U g f ; Cy/T(2;3) X a O 9 X' B 8 8MAXa = I n ( l)AM/scec cu ct u gf; +/T(2;3)Xa ? 0 x B 8 8
MAC =In(l)AM/sceccLrctvgf;Cy/+O OX B 8 8
1694 PAUL ROBERTS
TABLE 1
X chromosome inversions tested
~ ~
~ ~~~~~~
Salivary gland data Genetic data Inversion and markers Left break Right break Left break Right break
sc7
C1, y v B
dl-49, y f6"
AM
AB
1A lBlLLlB3.4 l'B2
1 B4
4A5
4D7-4E1
8C16-8D1
about 9F
16Ai .%l 6A5.6
16A1.2-16A5.6 4D7-4Ei
17 11D3-11D8 20BC
5D5
17A6
1 1F2-11 F4
16E2-16E3
13F1
20E 11F2-1 IF4 2OA3
+
left of y near fu
not determined between QC between bb
and sc and sp-a between sc between cv
and sui- and ct between sc between sy
and bi and f u
between rb between fw and cu and g
near lz between B and B x near U between g
and sd
not determined as above not determined
A standard mating procedure was followed. Fifteen to 20 virgin females not older than 24 hours were mated per half-pint bottle for six days and then dis- carded. Counts were made until the 19th day of culture. Laboratory temperature was maintained at 24" 2 1°C.
RESULTS
The data showing the effect of autosomal inversions on crossing-over and non- disjunction of X chromosomes heterozygous for various inversions are presented in Tables 2, 3 and 4. Before discussing the results it is necessary to clarify the parameters used to measure nondisjunction in the various crosses. The frequency of primary nondisjunction (percent XX eggs) was calculated by doubling the number of exceptional female progeny and dividing this number by the number of regular female progeny plus twice the number of exceptional females. This is to correct for the half of the XX eggs which are effectively lethal when fertilized
by X sperm, and to correct for the fact that exceptional males (percent nullo X
eggsJ2) may arise from processes other than nondisjunction (see below), With most crosses (subscript XP) involving Payne, Dfd heterozygotes, the number of exceptional female progeny was multiplied by four to correct for failure to recognize one half of the exceptional females. Regular females which
are B / + ; Payne, Dfd/+ are not distinguishable from exceptional females which are
+/+;
Payne, Dfd/+. This type of correction was not necessary for those crosses in which the exceptional non-B Dfd females were homozygous U and thusCROSSING-OVER A N D N O N D I S J U N C T I O N 1695
TABLE 2
A simultaneous measurement of X chromosonw crossing over and nondisjunction
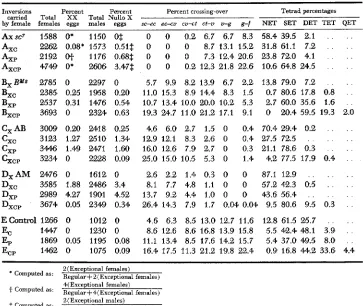
Inversions Percent Percent Percent crossing-over Tetrad percentages by carried female females eggs Total XX males Total Nullo eggs s c e c ec-cu cu+t ct-u u-g g-f NET SET DET TET QET
X
Axsc7 1588 O* 1150 O$ 0 0 0.2 6.7 6.7 8.3 58.4 39.5 2.1 . . . .
A,, 2262 0.08, 1573 0.51$ 0 0 0 8.7 13.1 15,.2 31.8 61.1 7.2 . . . .
Axp 2192 OJ- 1176 0.68$ 0 0 0 7.3 12.4 20.6 23.8 72.0 4.1 . . . .
Axcp 4749 Of 2606 3.47$ 0 0 0.2 12.3 21.8 22.6 10.6 64.8 24.5 . . . .
B , B M a 2785 0 2297 0 5.7 9.9 8.2 13.9 6.7 2.2 13.8 79.0 7.2 . . . .
BXc 2385. 0.25 1958 0.20 11.0 15.3 8.9 14.4 8.3 1.5 0.7 80.6 17.8 0.8 . .
B ,
, 2537 0.31 1476 0.54 10.7 13.4 10.0 20.0 10.2 5.3 2.7 60.0 35.6 1.6 . .
Bxcp 3693 0 2324 0.63 19.3 24.7 11.0 21.2 17.1 9.1 0 20.4 59.5 19.3 2.0
C,AB 3009 0.24) 2418 0.25 4.6 6.0 2.7 1.5 0 0.4 70.4 29.4 0.2 . . . .
C,, 3123 1.27 2510 1.34 12.9 12.1 8.3 2.6 0 0.4 27.5 72.5 . . . . . .
cxp
344.6 1.49 2471 1.60 16.0 12.6 7.9 2.7 0 0.3 21.1 78.6 0.3 . . . .&,,
3234 0 2228 0.09 25.0 15.0 10.5 5.3 0 1.4 4.2 77.5 17.9 0.4 . .D,AM 2476 0 1612 0 2.6 2.2 1.4 0.3 0 0 87.1 12.9 . . . . . . Dxc 3585 1.88 2486 3.4 8.1 7.7 4.8 1.1 0 0 57.2 4.2.3 0.5 . . . .
Dxp 2989 4.27 1901 4.52 13.7 9.2 4.4 1.0 0 01 43.6 56.4 . . . . . .
D,,, 3674 0.05 2349 0.34 26.4 14.3 7.9 1.7 0.04 0.04 9.5 80.6 9.5 0.3 . .
EControl 1266 0 1012 0 4.6 6.3 8.5 13.0 12.7 11.6 12.8 61.5 25.7 . . . .
EP 1869 0.05 11% 0.08 11.1 13.4 8.5 17.6 14.2 15.7 5.4 37.0 49.5 8.0 . .
EC 1447 0 1230 0 8.6 12.6, 8.6 16.8 13.9 15.8 5.5 42.4 48.1 3.9' . .
EOP 1462 0 1075 0.09 16.4 17.5 11.3 21.2 19.8 22.4 0.9 16.8 44.2 33.6, 4.4
Z(Exceptiona1 females)
4(Exceptional females)
* Computed as: Regular+2(Exceptional females) Computed as: ReaularC4iExce~tional females)
$ Computed as:
~~
Z(Exceptiona1 males) Regular+2(Exceptional males)
that the above assumption was justified. This fact and the equality of numbers of C y and non-Cy exceptional females indicate that there is no pronounced tendency of an X to preferentially associate with and disjoin from the inverted autosome rather than its normal homologue.
1696 PAUL R O B E R T S
TABLE 3
A simultaneous memurement of X chromosome crossing-ouer and nondisjunction
Inversions carried by female
Percent Percent Total XX Total Nullo X females eggs males eggs
Fx sc8
Fxc F X P F X C P
Gx Y4 GXC GXP GXCP
Hx CIB Hxc HXP HXCP
3497 0.06 3792 0.26 3905 0.41 2738 0.0
3138 0.13 2950 0.2 3047 0.4 2056 0.09
3223 0.18 2835 3.0 3013 1.62 2726 0.14
204.9 28 75 2918 2149 265 7 2678 2526 1933 934 700 460 420 7.1 15.2 21.8 25.2 6.0 17.4 27.1 34.8 0.5+ 6.1 11.6 37.6 Percent crossing-over
yz-c ec-cu cu-v u-f f - c a r
0.3 1.0 3.6 3.3 1.4 1.0 4.0 9.0 8.9 4.4 1.9 5.1 15.2 15.8 6.1 5.2 10.4 34.3 34.6 16.1
0.8* 1.4 2.3 3.1 0.6 3.2 4.9 6.4 9.9 0.9 4.7 10.0 12.0 17.4 1.6 11.9 17.4 29.4 37.2 3.2
. . O 0 0 0.3 . . 0 0 0 0.7
. . o
0 0 1.0. . 3.0 3.0 0 2.7
Crossover types
NCO SCO DCO TCO
2486 2438 1953 775 2468 21 13 1641 767 926 652 403 247 137 43 7 608 966
107 310 1 486 3 752 5
3 5 4 6 9
TA,BLE 4
A simultaneous measurement of X chromosome crossing-over and nondisjunction
Inversions Percent Percent Percent crossing-over Tetrad percentages
by female females eggs males eggs +ec ec-cu cu-U u-f f-car NET SET DET T E T QET
carried Total XX Total Nullo X -
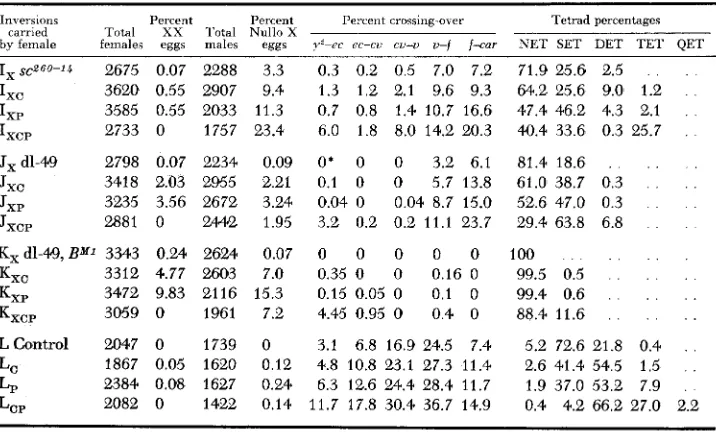
I X S C ~ G O - ~ ~ 2675
Ixc 3620 I X P 3585 I X C P 2733
Jxdl-49 2798 Jxc 3418 JXP 3235
JXCP 288 1
K, dl-49, B” 3343 Kxc 3312 KXP 3472 KXCP 3059
0.07 0.55 0.55 0 0.07 2.03 3.56 0 0.24 4.77 9.83 0 2288 2907 2033 1757 2234 2955 2672 244.2 26% 2603 21 16 1961
3.3 0.3 0.2 0.5 7.0 7.2 9.4 1.3 1.2 2.1 9.6 9.3 11.3 0.7 0.8 1.4 10.7 16.6 23.4 6.0 1.8 8.0 14.2 20.3
0.09 O * 0 0 3.2 6.1 2.21 0.1 0 0 5.7 13.8 3.24 0.04. 0 0.04 8.7 15.0 1.95 3.2 0.2 0.2 11.1 23.7
0.07 0 0 0 0 0 7.0 0.35 0 0 0.16 0 15.3 0.15 0.05 0 0.1 0 7.2 4.45 0.95 0 0.4 0
71.9 25.6 2.5 . . .
64.2 25.6 9.0 1.2 . .
47.4 46.2 4.3 2.1 . .
4Q.4 33.6 0.3 25.7 .
81.4 18.6 . . . ,
61.0 38.7 0.3 . . .
52.6 47.0 0.3 . .
29.4 63.8 6.8 . .
100 . . . . .
99.5 0.5 . . . . .
99.4 0.6 . . . . . .
88.4 11.6 . . . . . .
LControl 2047 0 1739 0 3.1 6.8 16.9 24.5 7.4 5.2 72.6 21.8 0.4 . .
LC 1867 0.05 1620 0.12 4.8 10.8 23.1 27.3 11.4 2.6 41.4 54.5 1.5 . .
LP 2384 0.08 1627 0.24 6.3 12.6 24.4 28.4 11.7 1.9 37.0 53.2 7.9 LCP 2082 0 1422 0.14 11.7 17.8 30.4 36.7 14.9 0.4 4.2 66.2 27.0 2.2
CROSSING-OVER A N D NONDISJUNCTION 1697 exceptional males was tested in order to detect and eliminate any Y-carrying females among those being studied.
From Tables 2-4, it is apparent that there are significant amounts of primary nondisjunction only in crosses (subscripts XC and XP) in which there is one autosomal inversion in the genome in addition to the X inversion. The controls and females carrying both Curly and Payne have rates which are not significantly different. The X inversions which, when present with either Curly or Payne, give statistically significant increases in percentage of primary nondisjunction when 95 percent confidence limits are compared with 95 percent confidence limits of their controls are: AB, AM, CIB, dl-49 and Ins( 1)dl-49, B M 1 . Zn(l)
sceso+4 has a significant but slight increase over the control value for both Ixc
and Ixp. Zn(l)sc8 shows a statistically significant increase in cross F,, but not in
Fx,. The experimental results of cross F must be considered as not significantly different from controls. Cross F,, is not as sensitive an indicator of nondisjunction as Fxc because the method of determining percentage of nondisjunction in the presence of the Dfd marker involves multiplication of the number of exceptional females by four with a resulting spread in confidence limits. Inversions
p,
sc7and BM2 give no significant increase over their controls.
The experiments being reported here were designed to measure not only the disjunctional properties of the X chromosomes but their exchange properties as well. The parameter most difficult to estimate is the frequency of crossing-over within the
X
inversions. Two measures of this are given in Table 3: recovered double crossovers and exceptional males. With the long sc8 and inversions (Fand G) there is a high proportion of exceptional males (as compared with ex- ceptional females). This is to be expected with long inversions where con- siderable double crossing-over is possible. Since double crossovers within a para- centric inversion are the only crossovers recovered and since there is a high proportion of these, the longest inversions (sc8 and y'+) do not appear to interfere with crossing-over to any marked degree, as STURTEVANT and BEADLE (1936) also concluded.
It is more difficult to determine whether crossing-over is reduced within shorter inversions since the frequency of double crossovers would be expected to be low. STURTEVANT and BEADLE (1936), using attached-X's with inversions, found that crossing-over is reduced in heterozygotes for sc7 and dl-49 and state that it is probably reduced in C1B. NOVITSKI and BRAVER (1954) have concluded that ex- change within a heterozygous dl-49 inversion is reduced to about one fourth of that expected of the same region without an inversion.
1698 P A U L ROBERTS
sc260-14 which is manifested by recovered doubles within the inversion and by the excess of exceptional males as compared with exceptional females.
The recovery of appreciable numbers of double crossovers within inversions and a pronounced excess of exceptional males as compared with exceptional females gives a rough estimate of the amount of undetectable crossing-over within a n inversion. Using these criteria, crossing-over appears to be near normal i n sc8 and y4, and there appears to be considerably more crossing-over within the relatively long GIB and sc260-J4 inversions than within the shorter inversions so that one cannot compare the total exchange in short with the total exchange in the long inversions. For this reason, inversions sc8, y4, CIB, and S C ~will be ~ ~ - ~ ~ excluded from the following comparison. A comparison of crossing-over among the remainder of the inversions must be only a rough approximation since the inversions differ in size, but crossing-over within the inversions must be of the same order of magnitude and low compared with crossing-over outside the inversions.
Tetrad analysis of the crossover data is given in Tables 2 and
4.
Table 3 lists only crossover types for sc8, y" and C1B for reasons stated above. The analysis was made using the formula derived by CHARLES from the generalized formula ofWEINSTEIN ( STEINBERG 1936), which assumes no chromatid interference. QET = 16 QCO
TET = 8(TCO - 4QCO) DET
SET
NET = NCO - SCO
+
DCO - TCO -I- QCO 4(DCO ~ 3TC0+
6QCO)2 (SCO - 2DC0
+
3TC0 - 4QCO)where NET = frequency of no-exchange tetrads, SET = frequency of single ex- change tetrads, etc. NCO = frequency of noncrossovers, SCO = frequency of single crossovers, etc.
This analysis provides the basis of a n interesting comparison between the exchange and disjunctional properties of X inversion heterozygotes. I n general, those inversion heterozygotes which give the greatest percentages of nondis- junction in crosses with subscripts XC and XP, are those which have undergone the least crossing-over and consequently have the highest percentage of no exchange tetrads. The linear regression of percentage of nondisjunction on percentage of no-exchange tetrads for crosses involving XC and XP is plotted in Figure 1. The percentage of crossing-over from f to car was not measured in the
sc7 heterozygotes thus a correction was necessary in order to compare this inver- sion with the others. Exchange in this region was estimated to be at least equal to the same region in the sc260-14 heterozygote (approximately ten percent). This brought the percentage of no exchange tetrads in crosses Axo and AXP from 31.8 percent and 23.8 percent to 12 percent and four percent, respectively. The regression of nondisjunction on percent of no exchange tetrads for crosses involv- ing either Curly or Payne is highly significant (P
<
.01).
CROSSING-OVER A N D N O N D I S J U N C T I O N
z 0
c
U
z
3
7
-
E
n
z 0
2 I-
z
Y
a
a w
4
/
1699
10
PERCENT NO-EXCHANGE TETRADS
FIGURE 1 .-Regression of percent nondisjunction on percent no-exchange tetrads for six X inversions.
that large numbers of exceptional progeny are obtained in crosses M A C X a and N d C X a and that they are all, or nearly all, Curly females and Xasta males. It is evident that the highly nonrandom assortment of the Curly inversion and the Xasta translocation among the exceptional progeny in M A C X a and N d c X a is the
1700 PAUL ROBERTS TABLE 5
A simultaneous measurement of X chromosome crossing-ouer and nondisjunction in females bearing an autosomal inuersion-translocation complex
Number of females Number of males
Exceptional Exceptional Percent Percent crossing-over
Constitution exceptional
of female Regular C y X a
+
Regular Cy X a+
progeny sc-ec ec-cu cu-t ct-u Total MAcx,AM 2770 301) 0 0 1926 0 44.0 0 13.6 10.3 9.5 4.4 0.8 25.0 MAXa 2125 . 0 2 1632 . . 0 0 0.05 8.1 7.8 4.2 0.5 20.6MAC 1984 11 . . 10 1419 9 . . 13 1.26 7.2 6.9 3.0 0.4 17.5
M* 1622 . . . . 3 1239 . . . . 1 0.1 1.8 1.5 0.6 0.2 4.1
Percent crossinr-over
u-f f-car Total
NdCXa dl-49 2234 203 6 0 1463 2 300 0 11.0 6.3 16.3 22.6
Ndxa 2298 . . 5 6 2017 . 4 8 0.5 6.7 11.3 18.0
N d C 1754 11 . 9 1363 13 . 14 1.48 4.6 11.0 15.6
Nd 1059 . . . . 1 850 . . . . 1 0.1 4.1 6.2 10.3
Since previous tests (BRIDGES 1916; STURTEVANT and BEADLE 1936) of excep- tional females to determine whether they carry crossover or noncrossover X
chromosomes have been made with secondary exceptions, and since it is known that not only is there partial homology between the X and the Y but that the two
X’s and the Y can form trivalents (COOPER 1948), it seemed desirable to test X
chromosomes that have nondisjoined in a system in which there is genetic evi- dence that an X has paired with a nonhomologous chromosome. Consequently the first 119 exceptional Curly females from cross MAO and the first 114 ex- ceptional Curly females from cross Ndcxa (Table 5 ) were progeny tested. All of the 233 exceptional females carried noncrossover X chromosomes. Further con- sideration of Table 5 will be found in the Discussion section of this paper.
DISCUSSION
A striking feature of the crosses in Tables
2-4
is that statistically significant numbers of exceptional female progeny resulting from nonhomologous pairing are recovered only when one, not two, autosomal inversions are present with theCROSSING-OVER A N D N O N D I S J U N C T I O N 1701
A
possible explanation that must be considered is whether exceptional progeny are always eliminated as dominant lethals when there are three inversions present. This is not likely from the following considerations. In a female with an X inversion and one autosomal inversion, if one X paired with the Curly inver- sion and the other X with the normal second chromosome, one would expect that eggs receiving two X chromosomes would be nullo-2 whereas nullo X eggs would be diplo-2. Fertilization would result in autosomal aneuploids that are lethals. It is apparent from the data that pairing is not exclusively of this type since exceptional progeny are recovered, often with considerable frequency. It is doubt- ful whether this type of pairing occurs with any appreciable frequency sinceCOOPER et al. (1955) have shown that egg mortality is usually not more than twice the percentage of XX plus nullo X eggs. Only if recovered exceptionals were a small fraction of the dominant lethals would the above type of pairing have to be considered seriously. The point is that according to the above scheme the opportunity for the recovery of exceptional females should actually be in- creased with two autosomal inversions present in addition to the
X
inversion since the X’s could pair with different autosomes thereby not producing auto- somal aneuploidy in all XX eggs. The failure to recover exceptional females when two autosomal inversions are present must then be the result of a reduction of X-autosomal pairing to the level of the controls.There are at least two possible explanations for the virtual absence of X- autosomal pairing in the presence of two autosomal inversions. The first is competition for associations among nonhomologues when there are more than two such elements present. GRELL and GRELL (1960) have shown that when there are present three elements which can pair nonhomologously, removal of one element may lead to increased association between the other two. They have also shown that different elements show different degrees of attraction for a non- homologue. It can be seen fmm Tables 2-4 that X-2 or X-3 pairing occurs with roughly the same facility with the various
X
inversions with either Curly or Payne although X-3 pairing usually exceeds X-2 pairing. In order to account, solely by competition, for absence of X-autosomal pairing when two autosomal inversions are present, one would have to postulate virtually 100 percent 2-3pairing with exclusion of the X. The inversions Curly and Payne are not unique in this respect since only .01 percent exceptional females are obtained with inversions dl-49, Curly and Dichaete present in the genome
(PTASHNE
1960). One would not expect two different combinations of autosomal inversions to show equal facility for pairing with competitive exclusion of the X. Therefore, it is un- likely that competitive pairing alone could account for absence of X-autosomal pairing. A second possible explanation is that the increased crossing-over observed when two autosomal inversions are present plays a role. This possibility will be discussed below.I t will be recalled that COOPER et at. (1955) postulated a mechanism by which
no-exchange X chromosome tetrads might be eliminated, resulting in an apparent increase in X chromosome crossing-over. I t involved X-autosomal pairing with
1702 PAUL ROBERTS
As can be seen by examination of Tables 2 4 , the greatest increase in crossing- over on the X is found when both Curly and Payne inversions are present in the genome. Clearly, the increase in crossing-over under these circumstances cannot be explained by selective elimination of noncrossover X tetrads whose elements were involved in X-autosomal pairing since there is no X-autosomal pairing. Also, in Table 4, cross K, elimination of noncrossover X tetrads could not account for the absence of crossing-over in K, with the appearance of crossover classes only in females heterozygous for at least one autosomal inversion. COOPER et al.
actually referred to the eliminated tetrads as “low rank tetrads” rather than no- exchange tetrads. The precise meaning of this phrase is not clear, but it might mean single exchange as well as no-exchange tetrads. However, in the present sample, it has been shown by progeny testing that 100 percent of the exceptional females carrying chromosomes that have been paired nonhomologously carry noncrossover X chromosomes. Since crossover chromosomes do not pair non- homologously, there is no known mechanism by which elimination of tetrads of higher rank than no exchange could occur, and REDFIELD (1957) has already shown that elimination of all noncrossovers would not account for the magnitude of recombination increase found under conditions of structural heterozygosity. It appears, then, that one interchromsomal effect is a real increase in crossing- over brought about by the presence of rearranged chromosomes, most typically inversions, in the genome. Since the elimination of noncrossover tetrads does not appear to be an important mechanism for increasing crossover frequencies
(except under special circumstances which will be discussed below), the seeming paradox of a single heterozygous autosomal inversion increasing both X chromo- some crossing-over and nondisjunction requires another explanation. If one considers the older theory of meiosis in which chromosomes pair once and for all, exchange in any homologous region and then disjoin, it is difficult to imagine a mechanism by which a heterozygous autosomal inversion could increase in- timacy of pairing of X chromosomes (as seems to be implied by increased crossing-over), yet increase the frequency of nonhomologous X-autosomal pair- ing if these events are as closely related in time and in configuration as is implied by the older theory of meiosis. A consideration of the data in Table 5 suggests a two phase model in which crossing-over and interchromosomal effects on crossing- over occur earlier in meiosis than the time of pairing for chromosome disjunction. Figure 2 illustrates the possible types of X-autosomal pairing with 2L-3R, the chromosome of T(2,3)Xa which is at a pairing disadvantage due to inversion heterozygosity in both arms (FORBES 1960). The figure must be considered purely schematic and based on inference from the genetic behavior of the chromo- somes. Figure 2A illustrates the possible situation in which one X is paired with 2L-3R while the other X is unpaired. This configuration might lead to frequent loss of the unpaired X, but the following considerations will show that this does not occur.
CROSSING-OVER A N D N O N D I S J U N C T I O N 1703
ln(2L)Cy
2RJlr
In (2 R) Cy 3R X I " \ , 2 L In(3R)P , 3R3 L A
2L
3 R
3 L
2 L
\LX2
E
FIGURE 2.-Diagrams of, (A), bivalent-univalent and, (B), trivalent formation in the In(l)/X; Cy/T(2;3)Xa genotype.
equal ratios of the following types of eggs (considering only alternate disjunction from the translocation) should be formed:
1 XX C y 1 nullo-X X a 1
x
Xa:lx
cyThe first two types of egg will result, upon fertilization, in equal numbers of exceptional Curly females and exceptional Xasta males and the second two egg
types, equal numbers of Curly and Xasta regular progeny.
If the unpaired X fails to be included with either nucleus the following types of egg will be produced:
1 nullo-X Xa:l X Cy.
1704 PAUL ROBERTS
1.4: 1 for both sexes. The reason for h i s is not known, but since females heterozy- gous for Curly and Xasta with normal X chromosomes also produced more Xasta than Curly progeny, it may be due to the greater relative viability of Xasta rather than a phenomenon related to nonhomologous pairing. A consideration of the exceptional males and females in Table 5, crosses M A C X a and N d C X a reveals
that the ratio of exceptional X a males to C y females is approximately 1.5: 1.
This is not sufficiently different from the ratio of X a to C y regular progeny so that one can say that loss of an unpaired X is frequent.
In the other possibility illustrated in Figure 2B both X’s pair with 2L-3R forming a trivalent. If there is random disjunctiomn from a trivalent, three types of egg should be formed with equal frequency:
1
x,
Xa:lx,
c y1
x,
Xa:lx,
c y1 X,X2 Cy:l nullo-X X a .
Fertilization should result in equal numbers of Curly exceptional females and Xasta exceptional males, and equal numbers of Curly and Xasta regular progeny. The data do not permit a choice between trivalent formation and the bivalent- univalent of Figure 2A, but if there is bivalent-univalent formation, loss of the unpaired X is infrequent.
The limits of the percentage of nonhomologous pairing can be estimated from the percentage of exceptional progeny. With inversion AM (cross MAC,,) the percentage of exceptional progeny is 740/5436 or 13.6 percent, Because excep- tional males were all Xasta and exceptional females all Curly, they can be lumped. It is reasonably certain that with this small inversion, exceptional males arose from nonhomologous pairing rather than any of the other mechanisms previously mentioned which would have resulted in equal numbers of Curly and Xasta exceptional males. The percentage of XX plus nullo X eggs is then 1480/ 61 76 or 23.96. Since random disjunction from a trivalent would lead to twice as many regular as exceptional progeny, the maximum amount of nonhomologous pairing that could be taking place is approximately 72 percent. In the unlikely event that disjunction of the X’s is always from a trivalent with both X’s going to one pole and the 2G3R element of the Xasta translocation going to the opposite pole, the minimum amount of nonhomologous pairing would then be 24 percent for MACXa. Similar calculations place the amount of nonhomologous pairing for N d C X a between 20 and 60 percent if there is trivalent formation. If, on the other hand, there is bivalent-univalent X-autosomal pairing, half the progeny are regular and half exceptionals as previously mentioned in the discussion of Figure 2A. Therefore, the actual hequency of nonhomologous pairing is ap- proximately 48 percent for MACXa and 40 percent for NdCXa. Since we do not know which of the two configurations occurs with the greater frequency the minimum amount of nonhomologous pairing assuming random disjunction from either is 48 percent for MACXa and 40 percent for NdCXa.
CROSSIIVG-OVER A N D N O N D I S J U N C T I O N 1705
Then, on the older theory of meiosis, no interchromosomal effect on crossing- over in regular progeny could result from nonhomologous pairing since all non- homologously paired X chromosomes go to exceptional off spring. Any increase in crossing-over in MACXa or NdCXa over their respective controls would have to be due to the Curly inversion, but the addition of the Curly inversion to Xasta should have little effect on X chromosome exchange because the number of heterozy- gous inversions present is not increased over the control. Since Xasta carries Zn(2R)Cy and Zn(3R)P, the addition of ins(2L
+
2R)Cy makes Zn(2R)Cy homozygous and 2L heterozygous for Zn(2L)Cy. In order to determine the effect of Curly in the presence of Xasta uncomplicated by a high frequency of non- homologous pairing, crossing-over on the X was measured in C y / X a and X a females without X inversions. Since heterozygous inversions modify exchange on the X, a strict comparison with MACXa and N d c X a is not possible. Crossing-over at the tip of the X was slightly greater in C y / X a females than in X a females, but there was no proximal difference. If crossing-over is computed by including the exceptional and eliminated progeny in the noncrossover class (see below), then crossing-over should be reduced by approximately two times the observed exceptions or 24 percent in the case of MAcxa and 20 percent in the case of N d C X a . The value of 19.1 obtained for MACXa is not significantly different from the con- trol value of 20.6, and a similar calculation for N d C X a results in a figure of 18.1 which is the same as its control (18.0). The absence of the expected crossover reduction is inconsistent with the older theory of meiosis even if 100 percent directed segregation from a trivalent occurs.If there is bivalent-univalent formation or random disjunction from a trivalent, even more pronounced crossover reductions are anticipated according to the older theory of meiosis. If chromosomes pair once and for all, crossover in homol- ogous regions and then disjoin, crossing-over should be reduced by 48 percent of the control value in the case of
MACXa
and by 40 percent of the control value in the case ofNdCXa
when exceptional progeny are included in the computation. Again, the experimental and control values mentioned in the preceding para- graph are not significantly different.The most reasonable explanation for the failure of high frequencies of non- homologous pairing to decrease crossing-over has recently been advanced by
GRELL (1962). She made simultaneous measurements of Y-2 pairing and cross- ing-over on chromosome 2 in XXY females heterozygous for an autosomal in- version and a translocation. GRELL has concluded that nonhomologous associa- tions do not produce no-exchange tetrads but that these associations take place only between elements of two no-exchange tetrads, i.e., no-exchange tetrads are not a consequence but a precondition of nonhomologous pairing. GRELL postulates that meiosis is divisible into the sequential events of “exchange pairing” between homologous loci which may be followed by exchange, and “distributive pairing” at which time noncrossover elements may pair nonhomologously.
1706 P A U L ROBERTS
without corresponding decreases in crossing-over are readily explainable. It is possible to interpret the data of Table 5 according to the two-phase theory of meiosis, for it is apparent from crosses MACXa and Nncxa that high frequencies of nonhomologous pairing do not interfere with crossing-over which is actually increased over the controls MAXa and NdXa. On the two-phase theory, since only the noncrossover chromosomes could pair nonhomologously, and thus be excluded from regular progeny, the crossover frequency in regular progeny would be expected to increase.
The primary action of inversion heterozygosity appears to be at the time of crossing-over when exchange between an inversion and its normal homologue is often reduced as discussed previously. It is at this time that the “interchromo- somal effect” on crossing-over in the other tetrads of the genome takes place, by a yet unknown mechanism. Inversion heterozygosity for two chromosomes is not sufficient for appreciable increases in nonhomologous pairing unless both inver- sions appreciably reduce crossing-over with their respective homologues (Tables
2 , 3 , sc8,
p,
BMa, and sc7 [corrected] ).
Nonhomologous pairing at the time of pair- ing for disjunction, “distributive pairing,” is only between noncrossover chromo- somes as shown here for the X and as shown for 2 by GRELL. It follows that the amount of nonhomologous pairing should be proportional to the percentages of no-exchange tetrads for each of the participating chromosomes.The linearity of the relationship between no-exchange X tetrads and amount of nonhomologous pairing (Tables
2 4
and Figure 1) implies that the percentage of autosomal no-exchange tetrads is fairly constant from experiment to experi- ment. The interchromosomal effects of the different X inversions on autosomal exchange ( STEINBERG and FRASER 1944) were apparently not diverse enough to disturb the linearity of the regression.CROSSING-OVER A N D N O N D I S J U N C T I O N 1707 If we reconsider the data of Table 5 from this new perspective, it is worthwhile computing what the crossover values of
MACXa
and NdCXa might be if the XX plus nulloX
eggs are taken into the computation by assuming that these represent eliminated noncrossover chromosomes from no-exchange tetrads. If in M A C X a 24 percent of the eggs are XX plus nullo X (see above),
then the total male progeny including eliminated noncrossover chromosomes should be 1926/0.76 or 2534. Dividing the number of crossovers by this figure gives a value of 19.1 which is not significantly different from the control value of 20.6. Similar calculations forNdcxa result in a figure of 18.1 which is also the same as the control (18.0). Therefore, elimination of noncrossover chromosomes by X aneuploidy can produce considerable changes in crossover values when there is a great deal of nondisjunction. Tables 2-4 show that nondisjunction is usually much lower than this and often absent even when the X is heterozygous for an inversion, while exchange is usually increased by inversion heterozygosity relatively independent of the amount of nondisjunction. Therefore changes in crossover values due to elimination of noncrossover chromosomes from autosomal nondisjunction and X aneuploidy must usually be minor when they occur.
The problem of the absence of X-autosomal pairing in the presence of both the Curly and Payne inversions when such pairing occurs in the presence of one or the other also deserves reconsideration in the light of GRELL’s hypothesis. If
crossing-over is greatly increased with all major chromosomes heterozygous for inversions, then the probability of simultaneous occurrence of two no-exchange tetrads in the same genome may be virtually zero. Since single exchange within the inversions leads to recovery of noncrossovers, this is not necessarily reflected by tetrad analysis in Tables 2-4. If this is the correct explanation, there may be
no 2-3 pairing either. Egg mortality observed under these conditions would then have to be explained by the suggestion of REDFIELD (1957) that it may be largely due to increased crossing-over within the inversions rather than auto- somal nondisjunction.
Finally, a comparison of crossing-over in controls E and L with the remainder of the crosses in Tables 2-4 permits some observations on the nature of the inter- chromosomal effect on crossing-over which are consistent with the proposal of
STEINBERG and FRASER (1944) that this effect is primarily physiological in nature. The crosses involving both autosomal inversions in addition to the X
inversions generally show proportionately greater increases in crossing-over in certain regions compared with the increases in E and L under the same condi- tions of autosomal inversion heterozygosity but lacking X inversions. (Compare
Ax and Axcp with E and ECP, regions u-g and g-f; C,CXcp with &Ecp, sc-ec;
Dx and DXCp with E and ECP, sc-ec;
I,
and,
,
,
I
with L andL,,,
f-car; andJxcp with L and Lcp, f-car.) In several of these crosses the absolute value for exchange in a given region, usually farthest from the breakpoints, is considerably increased over the same region in E,, or Lce. This is apparently true for crossing- over within inversions as well as without, as can be seen from the crosses involv- ing y 4 and SI?. The percent crossing-over for FXCP, cu-car and Gxcp, yz-f is com-
1708 PAUL ROBERTS
inversions do not interfere appreciably with crossing-over so it is possible to consult the tetrad analysis for
Lp
to estimate the percentage of unrecovered triples. This value of 27 percent suggests that there are enough unrecovered triple crossovers to again increase the percent crossing-over above the control. If this interchromosomal effect on crossing-over is not due to differential via- bility interactions between the inverted X’s and the various dominant and reces- sive markers used, it represents a synergistic effect of the X inversion acting with the autosomal inversions to produce crossover increases in certain regions of theX greater than the increases produced by the heterozygous autosomal inversions alone. This suggests that the “inter”-chromosomal effect may, in certain situa- tions, become an “intra”-chromosomal stimulator of crossing-over. Since there is a mechanical effect of a n inversion to decrease crossing-over with its normal homologue, it is doubtful whether there is a simultaneous stimulus to increase
crossing-over with its homologue which is also mechanical. It seems more likely that the interchromosomal effect on crossing-over is primarily physiological in nature.
SUMMARY
Simultaneous measurements of X chromosome crossing-over and nondisjunc- tion were made with combinations of ten different heterozygous X inversions and the Curly and Payne autosomal inversions. The percent of X nondisjunction with the different inversions showed a statistically significant linear regression on percent of noncrossover tetrads.
Increases in crossing-over were observed with either or both autosomal inver- sions present but X nondisjunction rarely occurred when both Curly and Payne were present. This is further evidence that the increase in crossing-over with heterozygous inversions in the genome is real and not merely a n artifact from elimination of noncrossovers involved in nonhomologous pairing.
High frequencies of nonhomologous pairing in inversion-translocation bearing females were not accompanied by a reduction in crossing-over. Progeny tests of exceptional females from such crosses showed that all carried noncrossover X
chromosomes. The results support the hypothesis of GRELL (1962) that meiosis in females of Drosophila melanogaster is separable into two phases, one concerned with crossing-over, the other with disjunction.
It is concluded that there are two interchromosomal effects, the primary one a physiological stimulus of crossing-over at the time of exchange when only homologues are paired. Subsequently, in the second effect, only elements of two no-exchange tetrads are sufficiently affected by less specific attractive forces that they pair nonhomologously. The extent to which nonhomologous pairing occurs subsequent to exchange depends, roughly, on the proportion of no-exchange tetrads present.
ACKNOWLEDGMENT
CROSSING-OVER A N D N O N D I S J U N C T I O N 1709
LITERATURE CITED
BRIDGES, C. B., 1916 Non-disjunction as proof of the chromosome theory of heredity. Genetics
BRIDGES, C. B., and K. S. BREHME, 1944 The Mutants of Drosophila melanogaster. Carnegie COOPER, K. W., 1948 A new theory of secondary non-disjunction in female Drosophila melano-
gaster. Froc. Natl. Acad. Sci., U S . 34: 179-187.
COOPER, K. W., S. ZIMMERING, and J. KRIVSHENKO, 1955 Interchromosomal effects and segre- gation. Prw. Natl. Acad. Sci., U.S. 4 1 : 91 1-914.
FORBES, C., 1960 Non-random assortment in primary non-disjunction in Drosophila melano- gaster. Proc. Natl. Acad. Sci., U.S. 4.6 : 22S225.
GRELL, R. F., 1957 Non random assortment of non-homologous chromosomes. (Abstr.) Genetics 42: 374.
Non random assortment of non-homologous chromosomes in Dromphila melanogaster. Genetics 44: 421-435.
A new hypothesis on the nature and sequence of meiotic events i n the female of Drosophila melanogaster. Proc. Natl. Acad. Sci., U.S. 48: 165-1 72.
GRELL, R. F., and E. H. GRELL, 1960 The behavior of non-homologous chromosomal elements involved in non-random assortment in Drosophila melanogaster. Proc. Natl. Acad. Sci., U.S. 46: 51-57.
Maintenance of a Drosophila stock center, in connection with investigations on the constitution of the germinal material in relation to heredity. Carnegie Inst. Wash. Yearbook 43: 164-165.
An analysis of crossing over within a heterozygous in- version in Drosophila melanogaster. Genetics 39 : 197-209.
The behavior of strong and weak centromeres at second anaphase of Drosophila melanogaster. Genetics 45 : 499-506.
Egg mortality and interchromosomal effects on recombination. Genetics 42: 712-728.
Interchromosomal effect on crossing over in Drosophila.
Cold Spring Harbor Symposia Quant. Biol. 16 : 175-197.
The effect of autosomal inversions on crossing over i n the X chromo- some of Drosophila melamgmter. Genetics 2 1 : 615-624.
Studies on the effect of X chromosome inversions on crossing over in the third chromosome of Drosophila melanogaster. Genetics 29: 83-103.
Contributions to the genetics of Drosophila melanogaster. 111. In- herited linkage variations in the second chromosome. Carnegie Inst. Wash. Publ. 278:
305-341.
The relations of inversions i n the X chromosome
to crossing over and disjunction. Genetics 21: 554-604. 1: 1-52, 107-163.
Inst. Wash. Publ. 552.
1959b
1962
MORGAN, T. H., and A. H. STIJRTEVANT, 1944
NOVITSKI, E., and G. BRAVER, 1954
PTASHNE, M., 1960
REDFIELD, H., 1957
SCHULTZ, J., and H. REDFIELD, 1951
STEINBERG, A. G., 1936
STEINBERG, A. G., and F. C. FRASEX, 1%
STURTEVANT, A. H., 1919