Copyright 0 1993 by the Genetics Society of America
Complex Function and Expression
of
Delta
During Drosophila Oogenesis
Laurel
B.
Bender, Pamela
J.
Kooh' and Marc A. T. Muskavitch'
Program in Genetics, Cell and Developmental Biology, Department of Biology, Indiana University, Bloomington, Indiana 47405
Manuscript received August 19, 1992
Accepted for publication December 24, 1992
ABSTRACT
Delta ( D l ) encodes a cell surface protein that mediates cell-cell interactions central to the specification of a variety of cell fates during embryonic and postembryonic development of Drosophila melanogaster.
We find that the Delta protein is expressed intermittently in follicle cells and in germ-line cells during stages 1-10 of oogenesis. Furthermore, Delta expression during oogenesis can be correlated with a number of morphogenetic defects associated with sterility observed in Dl mutant females, including failure of stalk formation within the germarium and subsequent fusion of egg chambers, necrosis in gerrn-line cells, and multiphasic embryonic arrest of fertilized eggs. We have also identified a Dl
mutation that leads to context-dependent defects in Dl function during oogenesis. Direct comparison of Delta protein expression with that of the Notch protein in the ovary reveals substantial, but incomplete, coincidence of expression patterns in space and time. We discuss possible roles for the Delta protein in cell-cell interactions required for cell fate specification processes during oogenesis in light of available developmental and histochemical data.
C
ELLS adopt specific fates as a consequence of specific intercellular communication events dur- ing the development of complex multicellular orga- nisms. T h e Delta ( D l ) gene plays a central role in many diverse cell specification events that depend on inter- cellular interactions throughout Drosophila morpho- genesis. T h e first evident zygotic requirement for Dlfunction is during embryonic neurogenesis, when it is essential for the proper partitioning of cells within the neurogenic regions of the embryo between neuronal and epidermal lineages (LEHMANN et al. 1983). Dl has been proposed to act in this process, in concert with the five other zygotic neurogenic genes, by participat- ing in an intercellular communication mechanism re- quired to preclude adoption of the neuronal fate by ectodermal cells within embryonic neurogenic regions (LEHMANN et al. 1983; ARTAVANIS-TSAKONAS and SIMPSON 199 1 ; CAMPOS-ORTEGA and JAN 1991). DL also has been shown to play a role in embryonic mesodermal development (CORBIN et al. 199 1).
In recent years, many roles for Dl during postem- bryonic development have been recognized. Dl me- diates cell fate decisions central to the formation of adult structures such as the sensory bristles, wings, legs and compound eyes during larval and pupal de-
velopment (DIETRICH and CAMPOS-ORTEGA 1984;
HEITZLER and SIMPSON 1991 ; T. R. PARODY and M. A. T. MUSKAVITCH, submitted; PARKS and MUSKAV- ITCH 1993). In addition, studies with temperature- sensitive DL mutants reveal many discrete phenocriti-
Center for Pediatric Research. Indiana University School ' of Medicine, Indi- Current address: Howard Hughes Medical Institute, Herman B. Wells
anagolis, Indiana 46202.
To whom correspondence should be addressed.
Genetics 133: 967-978 (April, 1993)
cal periods for Dl lethal and visible phenotypes during larval and pupal stages (T. R. PARODY and M. A. T. MUSKAVITCH, submitted). Furthermore, the complex and intriguing spatial distributions of Dl mRNA (Vds- SIN, VIELMETTER and CAMPOS-ORTEGA 1987; KOP-
CZYNSKI and MUSKAVITCH 1989; HAENLIN, KRAMAT-
SCHEK and CAMPOS-ORTEGA 1990) and protein (KOOH, FEHON and MUSKAVITCH 1993) throughout embryogenesis and postembryonic development sug- gest that DL functions in intercellular interactions in a wide variety of tissues during many stages of devel- opment.
Successful completion of Drosophila oogenesis also depends on a variety of intercellular interactions (SCHUPBACH 1987; MANSEAU and SCHUPBACH 1989; SPRADLING 1991), and previous studies by RUOHOLA et al. ( 1 99 1) have implied that DL function is required during multiple stages of this complex process. In this study, we investigate Dl function throughout Dro- sophila oogenesis by analyzing the relationship be- tween Delta protein expression patterns in the wild- type ovary and phenotypes observed in sterile Dl
mutant females and their progeny. We find that com- plex Dl expression during oogenesis is correlated with several specific defects we observe within developing
Dl mutant ovaries and with embryonic arrest of the progeny of DL mutant females.
DL encodes a cell surface transmembrane protein (VASSIN, VIELMETTER and CAMPOS-ORTEGA 1987; KOPCZYNSKI et al. 1988) that has been shown to inter- act with the protein encoded by at least one other neurogenic gene, N o t c h [ N (WHARTON et al. 1985;
968 L. B. Bender, P. J. Kooh and M. A. T. Muskavitch
1990). Postembryonic temperature-sensitive periods for
Dl
function overlap those for N in a number of tissues (SHELLENBARGER and MOHLER 1978; T. R.PARODY and M. A. T. MUSKAVITCH, submitted), and mutations in
Dl
or N produce similar phenotypes in adult ovaries (RUOHOLA et al. 1991; X u et al. 1992;this work). Extensive genetic studies suggest that di- rect interactions between the Delta and Notch pro- teins occur in vivo (VASSIN, VIELMETTER and CAMPOS- ORTEGA 1985; DE LA CONCHA et al. 1988; ALTON et al. 1989; SHEPARD, BROVERMAN and MUSKAVITCH
1989; BRAND and CAMPOS-ORTEGA 1990; X u et al.
1990). We have therefore compared the accumulation of Delta and Notch proteins during oogenesis and find that their highly regulated spatial and temporal expression patterns are quite similar, but do not co- incide directly during a number of stages of oogenesis. These data suggest that dynamic expression of Delta and Notch is related to a number of intercellular interactions central to the specification of cell fates during oogenesis.
MATERIALS AND METHODS
Drosophila strains: Drosophila cultures were maintained at 25" (unless otherwise stated) on standard cornmeal-mo- lasses-yeast-agar medium seeded with live baker's yeast. The
Dlw allele was first described by Xu et al. (1 990) and discovered in our laboratory to be a heat-sensitive, homo- zygous viable mutation (T. R. PARODY and M. A. T. Mus- KAVITCH, submitted). All other Dl alleles used in this work were maintained in balanced stocks in heterozygous combi- nation with TM6C, c u Sb e Tb ca [TM6C (CRAYMER 1984)l. Other chromosomes used include ru h th st cu sr Dl 9p39 e5
ca, ru h th st cu sr DlsF1o2 e' ca, st Dl6B37 e , ru h th st cu sr
(ALTON et al. 1988); DlBE23, ss DlBxe e4, ss DlBx9 e4 (ALTON et al. 1989); Dl" (JURGENS et al. 1984); and ss Dlcs (C. LEE- SHIN, M. LEVITON and J. W. POSAKONY, unpublished data). The w;T4b24c;DlRF stock contains two copies of the T4b24c
second chromosome, which carries two copies of a P element transposon that includes the entire Dl transcription unit as well as 7 kb of 5'-flanking sequences and 2 kb of 3'-flanking sequences (KOPCZYNSKI 1991). Homozygosity for the
T4b24c chromosome completely rescues several hypo- morphic Dl mutant phenotypes and partially rescues Dl null mutant phenotypes (KOPCZYNSKI 199 1). The wild-type strain used in this work was Oregon-R, hereafter referred to as wild type. Throughout the remainder of this paper, recessive markers have not been included with Dl allele designations.
Viability, egglay and hatch rate determination: Viability:
Duplicate crosses were performed in which three males of the genotype DlX/TM6C and three virgin females of the genotype Dly/TM6C were brooded daily in vials for six days at the stated temperature (Table 1). The female parent was
Dlw/TM6C in crosses generating heterozygotes that carry one DlRF allele. The female parent was D16B37/TM6C in crosses generating heterozygotes that carry one Dl6B37 allele and one allele other than Dlm. At least 180 progeny were scored as pupae and as adults to assess viability.
Egglay and hatch rate determinations: For the temperature- shift experiments, 20 virgin females of each desired geno- type, which had been raised at 18 O , were collected and held in yeasted vials for 1-5 days at 18'. Fewer females (numbers ~ 1 9 K 2 3 e I ca (LEHMANN et al. 1983); DfT3R)DlM2, ss DlBE3' e4 ro
are given parenthetically after eacipenotype) were assayed for the enot es DfT3R)DlM2/D1 (lo), DIBX9/DlRF (3),
D1'F1oz/Dl
5
(1 6), yp DBE"/DIRF (1 2) and D19p39/DlRF (1 9). Fe- males were then transferred with 20 wild-type males to an egglay chamber containing a Karo syrup-agar egglay plate spread with a thin coat of baker's yeast paste. Flies were preincubated at 22" for 2-3 days to allow the females to approach a steady-state egglay rate. Flies were then brooded on yeasted egglay plates for 24-hr periods for 3 days at 22"and then for 9 days at 29". The eggs on the plates were counted at the end of each 24-hr period, and up to 100 eggs from each plate were picked onto a grid on a fresh agar plate containing a dab of yeast paste at the center. These "hatch plates" were reincubated at 22" and rescored for hatching within 24-48 hr. At the end of each brood, the number of adult females that had died during the brood was noted. In all calculations of eggs laid/female/24 hr, dead females were assumed to have died at the beginning of the brood. For some genotypes [DfT3R)DlM2/DlRF, DlRF, DIRF/DIRF, D16B37/Dlcs and D1+/DlRF], egglaylhatch rate experiments were repeated on another occasion and gave results that were qualitatively very similar to those from the initial trial (data not shown). In these cases, data from one representative experiment is given in Table 1. For some genotypes (DBEZ3/DIRF, DIRF/DIRF, D16B37/D19K23, D16B37/Dlcs
and D1+/DIR3, control experiments were performed in which the egglaylhatch rate determinations were carried out for 8 days at 22". The data obtained (not shown) confirmed that, in the absence of a temperature shift, no significant decrease in egglay or hatch rate occurred from the beginning to the end of this time for these females. For the temperature-pulse experiment (Figure 2), 60 DIRF/DIRF
virgin female siblings raised at 18" were collected and conditioned with wild-type males at 22" as described for the temperature-shift experiments (above). Three egglay cham- bers, each containing 20 males and 20 females, were brooded for 24-hr periods as described above. After a 1- day brood at 2 2 " , the egglay chambers were pulsed at 29" for 0, 24 or 48 hr, and then returned to 22". Egglay and hatch rates were determined as described above.
Examination of Hoechst-stained ovaries: Flies were raised at 18", and adults were collected at 18" from 0-3 days post-eclosion. Male and female flies were conditioned together in bottles of medium coated with yeast paste. Flies were transferred to a fresh yeasted bottle daily, for four days, at 22". For temperature-shift experiments, flies were subsequently incubated at 29" for different amounts of time in yeasted bottles that were changed daily. Ovaries were then dissected in phosphate-buffered saline (PBS; 150 mM
NaCI, 14 mM Na2HP04, 6 mM NaH1P04, pH 7.0) and washed three times on ice in ice-cold PBS. During the last wash, the tubes containing the ovaries were shaken briskly to dissociate the ovarioles from each other. The tissue was then fixed in 4% (w/v) paraformaldehyde in PBS for 20 min at ambient temperature and washed three times in PBS at ambient temperature. Ovaries were then stained in 1 pg/ml Hoechst dye (H33258 or H33342, Sigma Chemical Com- pany, St. Louis, MO) in PBS for 1 min. Stained tissue was washed twice in PBS and mounted either in 50% glycerol
+
100 mM Tris-HC1, pH 8.0, or in gehtol (HEIMER and TAYLOR 1974). Stained ovaries were illuminated by epiflu- orescence using the U V channel of either a Zeiss Universal or a Zeiss Axioskop compound microscope (Carl Zeiss, Inc.) and photographed using a MC-63 or MC-100 35-mm cam- era (Carl Zeiss, Inc.) and Kodak Tri-X Pan 400 black and white film.Delta Function in Oogenesis 969
TABLE 1
Viability, fertility and visible defects associated with selected Dl genotypes
Viability" Egglay rateb Percent hatched'
18" 29"
Genotype Pupal Adult Pupal Adult 22" 29" 22" 29" Wings' ~ e g s d
Wild type ND ND ND ND 42 53 88 77 0 0
Dl+/Dl" 99 94 109 124 54 58 95 90 0 0
D16B'7/Dl" 92 102 0 0 12 2 69 0 (22) ND ND
Dx3R)DlM2/Dlwf 76 55 0 0 0.9 0.2 0 (7) 0 ( 1 ) 12 10
D1CS/D16B'7g 91 63 12 0 17 12 35 64 13 13
w:T4b24c:DlW ND ND ND ND 2 1 1 48 (33) 42 ND ND
D P I D P ' ~
105 95 0 0 18 4 81 5 (65) 4 0.3D I ~ ~ ~ I D ~ ~ ~ ~
101 98 76 96 0.4 2 0 (8) 6 (31) 4 0Values given as fraction of expected viability. For all inter se crosses between flies heterozygous for the various Dl alleles or a wild-type third chromosome, and the TM6C balancer chromosome, a Tb+/Tb- pupal ratio of 0.33 was assumed to indicate 100% pupal viability for the heterozygous non-balancer class; a Sb+/Sb- adult ratio of 0.33 was assumed to indicate 100% adult viability for the heterozygous non-balancer class. ND, not determined.
Egglay rates given as average number of eggs laid/female/day. The egglay rate at 22' is derived from counts made on the third day of incubation at 22". The egglay rate at 29" is derived from counts made on the third day after the shift to 29".
'
Hatch rates were scored for the same broods of eggs that were scored for egglay rates (footnote 6 ) . Up to 100 eggs were scored for each brood. When fewer eggs were scored, the actual number is indicated in parentheses.Wing and leg defects were scored according to a system described in MATERIALS AND METHODS. Wing scores range from 0 (no defects) to 17, and leg scores range from 0 (no defects) to 18. Average values are given for 10 adults of each genotype.
include DlB ''/Dl , Dl*'/Dl", and DIBEz'/Dl".
Other enot es that exhibit conditional, heat-sensitive decreases in viability and egglay and hatch rates similar to those of Dl"/DIRF
fDJ3R)DlMZ deletes the Dl gene (ALTON et al. 1988). Other enot es that exhibit nonconditional decreases in viabilit and e la and hatch rates similar to those of DJ3R)DlMZ/D1" include D 1 9 p 3 9 / D F , Dlg9/Dl" and DlsF'02/D1". Egglay rates for DlBx9/DlRYand D$F'o$D1"
were determined on the second day after the shift.
8
%
(Figure 3), as does D1cs/D16B'7, include D19K23/Dl , Dlcs/Dl", and D19K2'/D16B37.
Other genotypes that exhibit moderate eg la and hatch rates at
BY
22" or 29", and a low but significant level of egg chamber fusionabove for Hoechst-stained ovaries; wild-type flies were raised and conditioned at 25". Ovaries were dissected in PBS on ice, teased apart to separate ovarioles, and fixed as above, then washed three times in ice-cold PBS. Antibody staining was performed as described elsewhere for other Drosophila tissues (KOOH, FEHON and MUSKAVITCH 1993). The mouse anti-Delta polyclonal antiserum M5 stains wild- type Drosophila embryos, but does not stain embryos homo- zygous for DJ3R)DlM , whlch ehmlnates the Dl gene. The M5 antiserum also stains Delta-expressing S2 cells, but does not stain Notch-expressing S2 cells or untransfected S2 cells (KOOH, FEHON and MUSKAVITCH 1993). The rat anti-Notch polyclonal antiserum R135 (a gift of R. G. FEHON and S.
ARTAVANIS-TSAKONAS, Yale University, New Haven, Con- necticut) stains wild-type embryos in the patterns expected for Notch, and stains Notch-expressing S2 cells, but does not stain untransfected S2 cells (KWH, FEHON and MUSKAV-
ITCH 1993). Confocal microscopic images were collected using a Biorad MRCGOO system and MRC-850 laser attached to a Nikon compound microscope (wild-type data) or using a Biorad MRC6OO system and MRC-860 laser attached to a Zeiss Axiovert compound microscope (DIRF/DIRF data). Re- finement and processing of computer-generated images were performed as described elsewhere (KOOH, FEHON and MUSKAVITCH 1993).
Scoring of phenotypic defects in wings and legs: Each fly was given a score based on specific features of the right wing and the right set of legs. Points were assigned for specific defects commonly seen for Dl mutants, such that a score of zero indicated wild-type wings or legs, and a non- zero score indicated affected appendages.
Wing scores: one point was assigned for each specific defect, including each "delta," or widening, at the intersec- tion of wing veins 2-5 with the outer margin (4 points maximum); thickening of either crossvein (2 points maxi-
mum); wing veins 2-5 that contained any thickened portion
(4 points maximum); veins 2-5 for which thickening ex- tended over more than half the length of the vein (4 points maximum); wing margin notching; wing blade blistering; and ectopic wing vein material not connected to a vein. The theoretical maximum wing defect score was 17.
Leg scores: one point for any of the following structures that contained a defect, including thickening, shortening, and twisting: coxa, femur and tibia. For tarsal segments 1 through 5, defects were scored as the absence of those structures. The theoretical maximum leg defect score was
24, but scores never exceeded 18 because tarsal segments 1 and 5 were always present. For wing and leg scores pre- sented in Table 1, the average value for ten representative flies (five males and five females) is given.
RESULTS
970 L. B. Bender, P. J. Kooh and M. A. T. Muskavitch
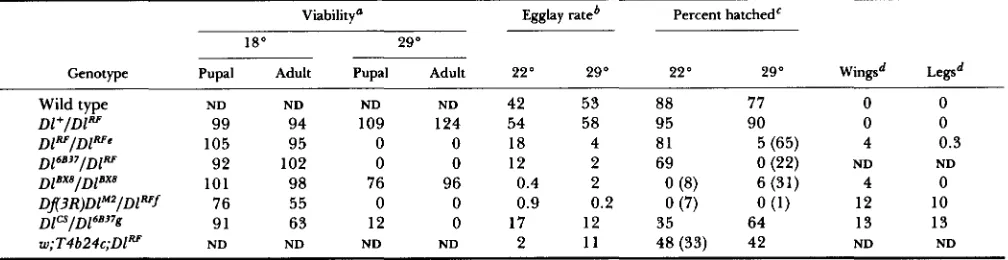
FIGURE 1 .-Delta protein expres- sion in the ovary. Ovaries from wild- type and Dl mutant adult females stained using anti-Delta antibodies, and photographed using confocal ep- ifluorescence microscopy. Anterior is to the left. (A) Germarium. Regions 1 , 2 and 3 (the stage 1 egg chamber) are labeled and separated by arrows. Arrowheads indicate vesicular fea- tures. (B and C) Young egg cham- bers. Stages are indicated by num- bers. Arrowhead in B indicates vesic- ular features. Arrow in C indicates a ring canal. (D) Detail of a stage 8 egg chamber. Arrowhead indicates junc- tion between the oocyte and a nurse cell where both cells abut the follicle cell layer. (E) Stage 9 egg chamber. Arrowhead points to a ring canal. (F) Posterior end of a stage 10A egg chamber. Arrows indicate cyto- plasmic vesicular features within fol- licle cells. (G) Anterior portion of a late stage 1 OB egg chamber. Arrows indicate vesicular features present within the stream of nurse cell con- tents entering the oocyte. (H) Ante- rior end of an ovariole from a DlRf/
DlRf homozygous female incubated at 29" for 72 hr before dissection. Large vesicular bodies (arrows) are visible in the cytoplasm of some of the nurse cells. Scale bar (panel B) 6
pm in panel A, 9 pm in panel D, 19 pm in panel E and 22 pm in all other panels.
germarium than in region 1. Delta accumulation pat- terns observed in these regions include diffuse cyto- plasmic staining and small, brightly stained vesicular features. Cells on the surface of the germarium in which Delta is expressed (Figure 1A) apparently cor- respond to profollicular mesodermal cells. Staining we detect within the germarium (KING 1970) implies that Delta is also expressed in germ-line derivatives within this portion of the ovariole. Delta protein lo- calization in the germarium is consistent with the demonstration by RUOHOLA et al. (1 99 1) that
DZ
tran- scripts accumulate within the germarium.Subsequent to the passage of a stage 1 egg chamber into the vitellarium, follicle cells divide during stages
1-6 of oogenesis and nurse cell and follicle cell nuclei undergo polyploidization. Within the vitellarium, Delta exhibits diffuse cytoplasmic accumulation in the cells within egg chambers during stages 1-3 (Figure 1, B and C). Delta protein appears to accumulate in vesicular features associated with the membranes of
nurse cells and oocytes beginning during stages 4-5
(Figure
1B).
T h e junctions between the follicle cells and the nurse cells and oocyte stain intensely during stages 5-6, while the membranes juxtaposed between adjacent follicle cells stain less intensely and the basal(i.e., external to the egg chamber) membranes of the follicle cells exhibit background levels of staining (Fig- ure 1, B and C). I t is not possible to determine the relative contributions of different cell types at the junctions between follicle cells and nurse cells, or
between follicle cells and the oocyte, to Delta mem- brane accumulation in these junctional regions with the resolution afforded by confocal microscopy. Cy-
toplasmic Delta accumulation in germ line-derived cells and follicle cells remains diffuse and uniform during these stages. Delta is also apparent in the membranes surrounding the ring canals that intercon- nect the nurse cells and oocyte (Figure 1, C and E).
Delta Function in Oogenesis 97 1
form a columnar monolayer over the oocyte. T h e egg chamber elongates during stages 7-9 of oogenesis, and the oocyte increases in volume relative to the nurse cell chamber. Yolk accumulates in the oocyte beginning during stage 8. Delta expression falls to background levels in the membranes at the junctions between the oocyte and follicle cells by stages 7-8,
while Delta levels remain unchanged in the mem- branes between nurse cells and follicle cells (data not shown, Figure 1 D). This expression pattern is consist- ent with in situ hybridization data indicating that Dl transcripts accumulate in nurse cells and in follicle cells that surround the egg chamber during this stage, except for those follicle cells at the posterior pole (RUOHOLA et al. 199 1). An abrupt demarcation at the nurse cell-oocyte boundary is visible in the stage 8 egg chamber between anteriorward follicle cells with high levels of membrane-associated Delta in lateral and apical membranes, and posteriorward follicle cells with little or no membrane-associated Delta (Figure 1D). Delta expression appears to decrease during stages 9 and 10A in lateral follicle cells that continue to migrate posteriorward to encompass the oocyte (Figure 1, E and F). Delta accumulation in follicle cell, nurse cell, and oocyte membranes within the follicle is distinctly reduced during stages 9 and 1 OA (Figure 1, E and F). Vesicular cytoplasmic Delta staining, which becomes more pronounced in all cell types during these stages, apparently corresponds to clear- ance of Delta from cell membranes via endocytosis.
Concomitant with the centripetal migration of a subset of follicle cells as they invaginate between the oocyte and the nurse cell chamber, the contents of the nurse cells stream into the oocyte during late stage 1 OB and stage 1 1. During the latter part of stage 10 (stage 10B, Figure lG), Delta becomes more highly concentrated at the boundary between the nurse cell chamber and the oocyte, and Delta protein appears in the oocyte cytoplasm. We infer that these patterns reflect the transfer of nurse cell contents into the oocyte. Delta is also expressed in a subset of the centripetally migrating follicle cells that come to re- side between the oocyte and the nurse cell chamber during stage 11 (Figure lG). The nurse cells degen- erate during stages 12-14, while the follicle cells complete synthesis of the chorion that encases the oocyte. From stage 11 through the completion of oogenesis, Delta accumulation is diffuse and near background levels throughout the follicle except at the nurse cell-oocyte border (see Figure 4G, below).
Multiple developmental defects in
DZ
mutant ova- ries are correlated with Delta expression patterns: RUOHOLA et al. (1 99 1) previously reported that female sterility observed for heat-sensitive D16B37/Dl" mu- tant females shifted to restrictive temperature was correlated with a number of defects in oogenesis. We have extended the investigation of Dl requirements100
80
5
c 605 40 U
a,
r m L
20
o ! . , . , . , . , . , . , .
0 A 2 v v 4 6 day8 10 12 .
FIGURE Z.-Fertility of DlRF females pulsed at restrictive temper- ature. (A) Egglay rates. The horizontal axis corresponds to the day on which eggs were counted. Adults were brooded 24 hr at 22" (day I), then shifted to 29" (large arrowhead) for 0, 24 or 48 hr. The 24-hr pulsed females were shifted to 22" after day 2 (leftward small arrowhead), and the 48 hr-pulsed females were shifted to 22" after day 3 (rightward small arrowhead). The relative depression in egglay rate observed on day 1 for all sets of females reflects the brief conditioning of the females on yeast paste before the experi- ment began. (B) Hatch rates. Broods of eggs collected during acquisition of data for panel A were scored for hatching. The horizontal axis in panel B corresponds to that in panel A.
during oogenesis through the use of temperature pulse protocols (Figure 2) and the analysis of a variety of Dl mutant genotypes (Table 1). We initially found that females homozygous for the heat-sensitive DlRF
mutation (T. R. PARODY and M. A. T . MUSKAVITCH, submitted), when mated to wild-type males, are fertile when raised to eclosion and maintained at 18 O
.
How- ever, these females become sterile when shifted to 29" following eclosion (Table 1 , data not shown). This sterility has two causes: reduction in egglaying, with kinetics similar to those reported by RUOHOLA et al. (1991), and reduction in the hatch rate of eggs that are laid, previously undescribed. These data imply that maternal Dl function is necessary for oogenesis and embryogenesis.972 L. B. Bender, P. J. Kooh and M . A. T. Muskavitch of decrease and recovery of egglay rates following a
24 hr pulse, monitored over short time intervals (6- 12 hr) after the pulse (data not shown), imply that reduction in Dl function prior to stages 6-8 of ooge- nesis is detrimental to egglaying. In contrast to egg- laying, initial pulse experiments with DIRF/DIRF fe- males revealed that hatch rates returned to control levels with recovery times dependent on pulse dura- tion following 29" pulses of 24 or 48 hr (Figure 2B). Decrease and recovery of hatch rates following a 24- hr pulse, monitored over short time intervals (data not shown), imply that Dl function is required between the 16-cell cyst stage in regions 2 and 3 of the ger- marium and stages 6-8 of oogenesis for successful completion of embryogenesis. For oogenesis and em- bryogenesis, stages during which Dl function is re- quired correspond to intervals during oogenesis within which we observe Delta protein expression.
Examination of ovarian morphology in a variety of
Dl mutants (see Table 1) revealed a number of distinct defects during oogenesis (Figure 3) that can be di- rectly related to wild-type Delta expression patterns. T h e predominant defect in ovaries from DIRF/DIRF
females shifted to 29" for at least 24 hr is the failure of stalk formation between germaria and stage 1 egg chambers (Figure 3C), a Dl mutant phenotype previ- ously noted by RUOHOLA et al. (1991). T h e stalk formation defect in DIRF/DIRF females can be rescued by homozygosity for a second chromosome that car- ries two copies of a P element transposon including the intact Dl transcription unit [T4b24c (KOPCZYNSKI
199 1 ; data not shown)]. We note further that in the place of stalks, follicle cell epithelia one or two cells in thickness form between 16-cell cysts that enter the vitellarium during heat treatment of DIRF/DIRF fe- males. These epithelia appear to degenerate when the 29" incubation period is extended to 72 hr, resulting in the formation of tubular follicle cell sheaths sur- rounding multiple oocytes and nurse cells containing necrotic nuclei (Figure 3D). In fact, few egg chambers appear to develop past stage 9 in DIRF/DIRF females incubated at 29" for 72 hr. It is likely that blockage of the ovarioles by degenerating multiple egg cham- bers is the cause of the irreversible egglaying defect described above for heat-pulsed DIRF/DIRF females (Figure 2). Similar defects, with varying degrees of severity, are also observed for a variety of Dl mutants (Figure 3, see legend).
Analysis of the D1+/DlRF embryos laid by DIRF/DIRF
females, following mating to wild-type males, when the hatch rate is reduced to its lowest point revealed that development in these embryos arrests multiphas- ically. Arrest phenotypes include failure of fertiliza- tion, distorted body axes, and neural hypertrophy; many arrested embryos exhibit no obvious mutant phenotype (data not shown). We also find that reduc- tions in hatch rates are rescued by the T4b24c trans-
poson (Table l), as described above for ovarian de- fects.
We assessed Delta expression in ovarioles from
DIRF/DIRF females incubated at 29" for 72 hr. In a typical mutant ovariole (Figure lH), Delta protein appears to be expressed at levels comparable to those in wild type, but exhibits a subcellular distribution pattern that differs from that seen in wild type in several respects. Staining appears more intense within some regions of the cytoplasm and less intense within the membranes of nurse cells. Follicle cell membranes show varying intensities of staining, and apical concen- tration of Delta in follicle cells is not observed in some regions within the ovariole. In addition, large vesicu- lar bodies that contain Delta, in contrast to the smaller vesicles seen in wild-type ovaries, appear in nurse cell cytoplasm. We speculate that the altered Delta distri- bution pattern in heat-treated DlRF homozygotes may result from trafficking defects associated with the DlRF
mutant protein at restrictive temperature, and that the large vesicles contain arrested trafficking inter- mediates.
The DZBx8 allele exhibits context-dependent de- fects in ovarian function: Previous studies have re- vealed that Dl function exhibits context dependence,
i e . , the relative severity of Dl mutant phenotypes correlated with a given mutant genotype can vary for different imaginal tissues in a single organism (C. L. REGAN, L. A. SILVER and M. A. T. MUSKAVITCH, in preparation). T o determine whether this cellular "context dependence" of Dl function extends to the ovaries as well, we scored pupal and adult viability and wing and leg defects for Dl genotypes for which we had examined female fertility and ovarian mor- phology (Table 1, Figure 3). Most of the Dl mutant genotypes we examined (e.g., DERF/DJ3R)DlM2) yield comparably severe effects for all phenotypes scored. We infer that these genotypes result in a general reduction in Dl function in all tissues.
In contrast, other Dl mutant genotypes affect only subsets of imaginal tissues. For instance, DlBX8/DlBx8
females lay few eggs (Table 1) and their ovaries display fused egg chambers and necrosis (Figure 3, G and H), while these flies exhibit wild-type viability and leg development, and only slight defects in wing devel- opment. None of these phenotypes show significant variation within a range of temperatures from 18 " to 29" (Table 1, data not shown). Within ovaries from
DlBX8/DlBx8 females, levels and spatial distributions of Delta protein expression are similar to those of wild type (data not shown), implying that the apparent attenuation of Dl function in ovaries of this genotype is not due to tissue-specific reduction in Dl expression. Another set of Dl mutant genotypes ( e . g . , D1cs/D16B37)
Delta Function in Oogenesis 973
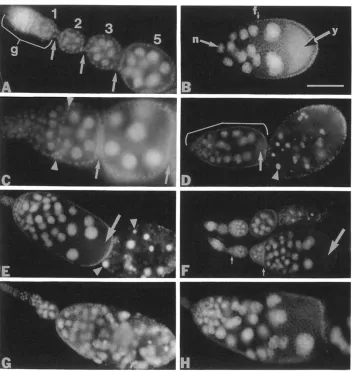
FIGURE 3.-Ovaries from Dl mutant and wild-type flies. Ovaries from adult females were stained with the DNA-intercalating dye Hoechst, and photographed using epifluorescence microscopy. Anterior is to the left. (A) Anterior portion of an ovariole from a Dl+/DIRF female conditioned at 22" before dissection. This tissue is morphologically indistinguishable from ovaries of wild-type females incubated at 22" or
29" (data not shown); g, germarium. Developing egg chambers are numbered according to stage of oogenesis. Stalks connecting egg chambers are indicated by arrows. (B) Stage 9 egg chamber from a Dl+/DIR' female conditioned at 22". This tissue is morphologically indistinguishable from ovaries of wild-type females incubated at 22" or 29" (data not shown). Diffuse autofluorescence in the posterior portion of the egg chamber indicates the presence of yolk (y); n, nurse cell nucleus; f, follicle cell nucleus. ( C ) Anterior tip of an ovariole from a DIRF/DIR'
female conditioned at 22", then incubated at 29" for 24 hr before dissection. Arrowheads indicate a layer of follicle cells that separates successive cysts. Older egg chambers are separated by stalks (locations indicated by arrows). (D) Ovariole from a DIRF/DIR' female conditioned at 22", then incubated at 29" for 72 hr before dissection. The anterior end of this ovariole (bracket) is comprised of several egg chambers fused with the germarium. Arrow indicates yolk at the posterior end of this structure. Arrowhead indicates a pycnotic nurse cell nucleus within necrotic stage 8 or 9 egg chamber@) at the posterior end of this ovariole. (E) Ovariole from a D~3R)DlM2/DlRF female conditioned at 22". Arrowheads indicate pycnotic nurse cell nuclei. Arrow indicates yolk. (F) Two ovarioles from a Dlcs/D16B'7 female conditioned at 22". then incubated for 72 hr at 29" before dissection. The anterior end of the upper ovariole has wild-type morphology. The lower ovariole exhibits a partially expressive mutant fused egg chamber phenotype. Small arrows point to positions where stalks have failed to form between egg chambers. Large arrow indicates yolk. The posterior-most egg chamber in this ovariole has twice the normal number of nurse cell nuclei.
(G and H) Ovarioles from DlBx8/DlBx8 females conditioned at 22". ( G ) Dl mutant stalk formation defect shows partial expressivity: the anterior end of the ovariole contains egg chambers with wild-type morphology, while the posterior end is made up of fused egg chambers. (H) Greater expressivity of the stalk formation defect, with the absence of necrosis. Scale bar (panel B) 20 pm in panels A and C; 80 pm in panel G and
50 pm in all other panels.
Egglay rates and hatch rates for females of these the ovary in a context-dependent fashion.
genotypes are relatively high at 18" and 29" (Table Ovarian expression of Delta and Notch are simi- l), and their ovaries are only mildly affected by 29" lar, but not identical: Temperature-sensitive N mu-
incubation (e.g., Figure 3F). tants incubated at restrictive temperatures (RUOHOLA
These data reveal that DlBx8 is a distinctly ovary- et al. 1991;
Xu
et al. 1992) exhibit ovarian defects biased Dl allele, and that other Dl mutant genotypes very similar to those observed for DZ mutants(Ruo-
974 L. B. Bender, P. J. Kooh and M. A. T. Muskavitch
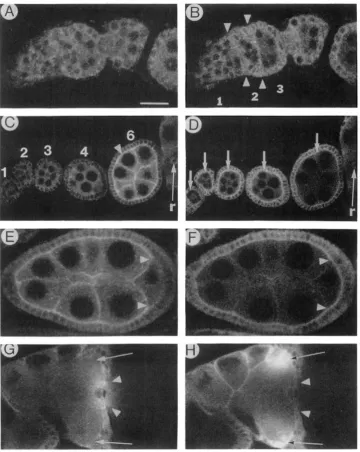
FIGURE 4.-Comparison of Delta and Notch protein expression in the ovary. Ovaries from wild-type adult females were stained with a mixture of anti-Delta and anti-Notch antibodies, and photographed using dual-channel confocal epifluorescence microscopy. These images are less well resolved than those in FlGURE 1 because a larger pinhole setting is required for dual channel imaging. Anterior is to the left.
(A, C , E and G) Tissues observed in the fluorescein (Delta-specific) channel. (B, D. F and H) Same tissue samples as in A, C. E and G, respectively, observed in the Texas red (Notch-specific) channel. (A and B) Germarium. Areas between opposing pairs of arrowheads in B contain invaginating mesodermal cells, with membranes that contain Notch. Numbers between and flanking arrowheads indicate regions of the germarium. (C and D) Young egg chambers. Stages are indicated in C. Arrowhead in panel C indicates Delta in membranes at the nurse cell-follicle cell junction of a stage 6 egg chamber. Arrows in panel D indicate Notch in follicle cell membranes of stages 1-6 egg chambers. Notch may also be present in nurse cell membranes. The ring canal (r) stains with Delta and Notch antibodies. (E and F) Stage 8 egg chamber. Notch is visible in the juxtaposed oocyte and follicle cell membranes, where Delta is barely detectable (note areas between arrowheads). (G and H) Anterior end (nurse cell cap) of a stage 1 1 egg chamber. As in Figure lG, Delta protein is visible as it streams into the oocyte, but Notch protein is not visible in this stream (note areas between arrowheads). Delta is prominent at the oocyte-nurse cell boundary. Notch appears to be associated with the specialized follicle cells that give rise to the chorionic appendages of the eggshell: Delta is much less prominent in these regions (compare arrows). Scale bar (panel A) 8 pm in panels A and B, 24 pm in panels E and F and 14 pm in all other panels.
bryonic development (T. R. PARODY and
M.
A. T. Delta and Notch exhibit diffuse cytoplasmic accu-Delta Function in Oogenesis 975
In contrast, membrane-localized Delta is not as clearly evident in these cells (Figure 4A). Within the vitellar- ium, Delta is absent from the membranes of stalk cells throughout oogenesis (e.g., Figure 4A), while Notch accumulates in these membranes (Xu et al. 1992).
Delta and Notch exhibit relatively constant levels of diffuse cytoplasmic expression in stage 1-6 egg chambers, but their membrane accumulation patterns differ dramatically during these stages. Notch is pres- ent in the follicle cell membranes of stage 1-6 egg chambers (Figure 4D), while Delta is not apparent in these membranes until stage 4-5 (Figure 4C). Delta and Notch display apically polarized accumulation patterns within the membranes of all follicle cells during these stages. However, little if any Delta ac- cumulates in the basal follicle cell membranes, where Notch is present. Delta and Notch proteins also ac- cumulate in nurse cell membranes and cytoplasm. However, nurse cell membranes stain very intensely for Delta in comparison to Delta staining within the entire cyst (Figure 4C), while these membranes stain relatively weakly for Notch in comparison to overall Notch staining (Figure 4D). Furthermore, levels of Delta protein in the nurse cell membranes increase rapidly during stages 4-6, while levels of Notch pro- tein in these membranes remain fairly constant.
By stages 7-8, Delta is no longer present at the junctions between posterior follicle cells and the oo-
cyte (data not shown, Figure 4E), where Notch protein is still apparent (Figure 4F). In fact, Notch protein
continues to be expressed in the membranes of all of the follicle cells that surround the egg chamber during stages 7 through 9. During stage 9 and continuing through the completion of oogenesis, Notch accumu- lation in follicle cell membranes gradually decreases and is present only at low levels in late stage follicles
(XU et al. 1992), as we observe for Delta (Figure lG; data not shown). Notch persists in nurse cell mem- branes during stages 11-12. In addition, Notch ac- cumulates during stage 1 1 in two specialized groups
of follicle cells situated dorsolaterally at the nurse cell chamber-oocyte junction (Figure 4H). No correspond- ing Delta protein accumulation is seen in these cells (Figure 4G). These cells eventually form the chorionic
appendages (KING 1970; PARKS and SPRADLING
1987), which often develop abnormally in the ovaries of
N‘”’
females (XU et al. 1992). T h e stream of nurse cell contents that continues to enter the oocyte during stage 11, which apparently contains Delta protein (Figure 4G), does not contain detectable Notch pro- tein (Figure 4H).DISCUSSION
Complex Delta expression correlates with pleio- tropic Dl function during oogenesis: Analysis of the developmental correlates of female sterility observed in Dl mutants reveals multiple defects that imply mul- tiple requirements for Dl function during oogenesis
(RUOHOLA et al. 199 1 ; this work). Patterns and dynam- ics of Delta expression also are correlated with pleio- tropic Dl function within the ovariole. Delta expres- sion in follicle cells may reflect multiphasic require- ments for Dl function in subsets of follicle cells during oogenesis, while the significance of Delta expression in the nurse cells is less clear. T h e multiphasic Delta expression we observe is also consistent with previous data regarding multiphasic accumulation of Dl tran- scripts during oogenesis (RUOHOLA et al. 199 1).
Mutant analysis and protein localization identify the germarium as a structure within which Dl function is required. Dl function is required for the formation of stalks between egg chambers (RUOHOLA et al. 1991; this work), but does not appear to be required for the migration of profollicle cells between 16-cell germ line-derived clusters. Even though follicle cell septa do form between successive egg chambers in Dl mu- tants, they apparently break down later in oogenesis resulting in the “fusion” of multiple egg chambers within ovarioles. Stalks may therefore serve a vital function in maintaining egg chamber integrity. Fur- thermore, maintenance of individual egg chamber integrity may be critical to continued follicle growth and development because we observe necrosis in fused multiple egg chambers. Delta expression in regions 2 and 3 of the germarium appears to be correlated with Dl function required for partitioning of a subset of follicle cells of the egg chamber into stalk cell and polar cell precursor populations (RUOHOLA et al.
Dl function is also required for specification of the correct number of posterior polar cells within devel- oping follicle cell populations preceding formation of the vitelline membrane and chorion (PARKS and SPRA-
DLING 1987; FASANO and KERRIDGE 1988; GROSSNIK-
LAUS et al. 1989). RUOHOLA et al. (1991) ,have dem- onstrated that egg chambers from Dl mutant females contain excess posterior polar cells. This apparently results from errors in cell fate specification, within a subset of posterior follicle cells, that become evident during stages 6-8 of oogenesis. We find that Delta is expressed in all follicle cells during stages 5-6 and that expression ceases in posterior follicle cells by stages 7-8 and in lateral follicle cells during stages 9 and 10. We infer that expression of Delta in posterior follicle cells preceding stage 8 is required for proper partitioning of posterior follicle cells into polar cell and flanking cell populations (RUOHOLA et al. 1991).
We find that Delta is expressed throughout most, if not all, of the somatic and germ-line cells within the germarium, but that expression ceases in stalk cells following their specification. Similarly, Delta is ex- pressed in all follicle cells during stages 5 and 6, and this expression ceases in posterior follicle cells follow- ing the appearance of polar cell and flanking cell populations (RUOHOLA et al. 1991). These patterns are analogous to temporal expression patterns we have
976 L. B. Bender, P. J. Kooh and M . A. T. Muskavitch observed for Delta during specification of neuroblasts
and bristle precursor cells during embryonic and post- embryonic development, respectively (KOOH, FEHON and MUSKAVITCH 1993). In these cases, Delta is ini- tially expressed throughout groups of cells within which particular fates are being specified and ceases in subsets of cells that stably assume alternative fates within such groups. In these latter instances, Dl func- tion is required to prevent the stable adoption of neuroblast or bristle precursor cell fates within subsets of ectodermal cells within embryonic neurogenic re- gions and imaginal proneural groups (LEHMANN et al. 1983; SKEATH and CARROLL 1991, 1992; PARKS and MUSKAVITCH 1993). If this qualitative aspect of Dl function is conserved in all tissues, then Dl function is required to inhibit adoption of the polar cell precursor fate in follicle cells that stably adopt the stalk cell fate early in oogenesis and to inhibit the adoption of the polar cell fate in posterior follicle cells that stably adopt flanking cell fates during mid-oogenesis, as ini- tially proposed by RUOHOLA et al. (1 99 1).
In each of these instances, we propose that Delta is initially expressed by all cells within populations that share a given developmental potential and that Delta expression ceases in cells that stably adopt alternative fates. These findings may be more easily reconciled with the premise that Delta functions by a mutual inhibition mechanism, as proposed by GORIELY et al. (1991), rather than by a lateral inhibition mechanism [see ARTAVANIS-TSAKONAS and SIMPSON (1 99 l)]. In the former case, Delta would function as a component of an intercellular signaling mechanism that precludes stable adoption of an alternative, secondary fate by all members within an equivalence group. Cells that with- draw from the inhibitory network formed by cells within the equivalence group would then be able to stably adopt the secondary fate. Cessation of Delta expression would correlate with withdrawal from a network of interacting cells and could be a necessary prerequisite for the stable adoption of a given second- ary fate.
T h e aberrant localization of bicoid mRNAs in Dl mutant egg chambers indicates an additional require- ment for Dl function during oogenesis and implies that wild-type Dl function is essential for establishment of the anterior-posterior (A-P) axis within the oocyte (RUOHOLA et al. 1991). Establishment of this axis is reflected by the polarized distributions of bicoid and
oskar transcripts that become apparent during stages 6-8 of oogenesis (RUOHOLA et al. 199 1). We find that Delta is expressed in somatic and germ line-derived components of the follicle preceding and during this developmental interval. T h e existence of a germ line- dependent system for patterning of the oocyte and follicle cells that requires the function of cappuccino, spire, fs(1)KlO and other genes (MANSEAU and SCHUP-
BACH 1989) raises the possibility that Delta expression
in germ-line derivatives could be essential for this process. However, RUOHOLA et al. (1991) inferred that Dl function is not required in the germ line for oogenesis based on the absence of visible morpholog- ical defects in ovarioles that contain Dl mutant germ- line cells and wild-type somatic cells. Given these data, it appears that Delta expression in follicle cells is essential for establishment of the A-P oocyte axis, as suggested by RUOHOLA et al. (1 991), and that Delta protein expression in germ line-derived cells may re- flect a redundant or cryptic role for Delta in the germ line.
Maternal Dl function is required for embryogen- esis: We have shown that reduction of Dl function in females also leads to reduced hatch rates of eggs laid following fertilization by wild-type fathers, implying that maternal Delta expression is required for the completion of embryogenesis. Arrested embryos ex- hibit an array of phenotypes, ranging from severe derangements of the embryo to apparently wild-type morphology. This variability presumably arises, in part, because DlRF encodes detectable levels of protein and yields only partial loss of Dl function in many tissues at restrictive temperatures (T. R. PARODY and M. A . T . MUSKAVITCH, submitted). T h e finding by RUOHOLA et al. (1 99 1) that some bicoid transcripts are incorrectly localized in Dl mutant ovarioles implies that maternal-effect lethality associated with reduced maternal Dl function may result, at least in part, from defects in establishment of the A-P axis within the developing oocyte.
We have also found that Delta protein appears to be transferred into the oocyte along with other nurse cell contents during stage 10 of oogenesis, and have previously shown that maternal Dl transcripts are loaded into the unfertilized egg (KOPCZYNSKI et al. 1988). However, our temperature pulse analysis of Dl mutants implies that Dl function may be dispensable after stage 8 of oogenesis. T h e functional roles of maternally loaded Dl transcripts and protein during embryogenesis therefore remain unclear.
A proposed structural basis for context-dependent
Delta Function in Oogenesis 977
the overall spatial and temporal patterns for Delta expression associated with this allele are comparable to those of wild-type Delta. We therefore favor the hypothesis that cell type-specific variation in the func- tional impact of DlBx8, and other tissue-biased Dl mu- tations ( C . L. RECAN, L. A. SILVER and M. A. T . MUSKAVITCH, in preparation), can result when such mutations affect different members of the epidermal growth factor (EGF)-like repeat array within the Delta extracellular domain (VASSIN, VIELMETTER and CAM- POS-ORTEGA 1987; KOPCZYNSKI et al. 1988). It is possible that interaction of Delta with particular pro- teins could depend on the integrity of particular EGF- like repeats within different cellular contexts. Analysis of a variety of mutations that differentially affect function of the Notch protein in different tissues provides ample precedents for this hypothesis: tissue- biased disruptions in N function are correlated with missense mutations in different subsets of EGF-like repeats within the Notch extracellular domain [see REBAY et al. (1 99 l)].
D o Delta and Notch function coordinately as sig- nal and receptor during oogenesis? Analysis of Dl and N functions in the establishment of cell fates within imaginal proneural groups has supported the hypothesis that Delta functions as a signal and Notch as a receptor necessary for proper adoption of epider- mal and neural fates during metamorphosis (HEIT- ZLER and SIMPSON 1991). T h e hypothesis that Delta
and Notch physically interact is further supported by the finding that Delta-expressing cells can aggregate with Notch-expressing cells in culture (FEHON et al. 1990). T h e temporal coordination of Delta and Notch expression in a variety of cell populations within which cell fates are being specified, and apparent coendo- cytosis of Delta and Notch during and after cell fate specification, further imply that Delta and Notch func- tion as a signal-receptor pair in UZUO during embryonic
and postembryonic development (KWH, FEHON and MUSKAVITCH 1993).
If Delta and Notch function as a signal-receptor pair during oogenesis, we would predict, based on precedents during embryonic and postembryonic de- velopment, that their expression should be temporally coordinate in cell populations in which their coordi- nate function is required for cell fate specification. In fact, we find that Delta and Notch expression do overlap spatially and temporally, for the most part, for stages during which either is expressed. T h e spa- tial and temporal dynamics of Delta and Notch expres- sion in follicle cells are consistent with involvement of Delta and Notch in signalling pathways central to the specification of distinct cell types within follicle cell populations (PARKS and SPRADLING 1987; FASANO and KERRIDGE 1988; GROSSNIKLAUS et al. 1989). Delta and Notch may also function coordinately in the oo- cyte reorganization that takes place between stages 7
and 9 of oogenesis, which results in the localization of certain messages to either the anterior or the posterior pole of the embryo (MANSEAU and SCHUPBACH 1989; RUOHOLA et al. 199 1 ; THEURKAUF et al. 1992). T h e distinct concentrations of Delta and Notch in the apical membranes of follicle cells and/or germ-line cells of stage 5-6 egg chambers could reflect roles for Delta and Notch in soma-germ line signaling necessary for this reorganization (MANSEAU and SCHUPBACH
1989; RUOHOLA et al. 199 1 ; THEURKAUF et al. 1992; CHEUNG, SERANO and COHEN 1992).
Delta is probably not involved in every process that requires Notch during oogenesis. For instance, Delta does not appear to accumulate at stage 11 in regions where dorsal appendages will develop during stages 12-1 4, within which we and others (Xu et al. 1992) have observed expression of Notch. Our examinations of DL mutant ovaries revealed very few instances of aberrant dorsal appendage formation, a phenotype described for N mutant ovaries (XU et al. 1992). It is entirely plausible that Delta will not be required for every process that requires Notch, particularly in light of the possibility that Notch may interact with a variety of ligands (REBAY et al. 1991). Genetic and biochem- ical approaches should allow us to identify proteins in addition to Notch that interact with Delta, within the ovary as well as in other tissues, during development in Drosophila.
We thank S. ARTAVANIS-TSAKONAS for suggestions and the use of laboratory facilities, R. FEHON and L. CARON for technical advice on antibody staining, and G. CRAMER for her deft technical assist- ance with dissections and viability scoring. Thanks to L. SILVER for alerting us to the fertility defects of DlBx8/DlBx8 females. We are grateful to T . KAUFMAN and the Howard Hughes Medical Institute for use of the Slide Maker, and to the Indiana Institute for Molec- ular and Cellular Biology for use of the confocal microscope. We also thank C. BERG, A. PARKS, T. S C H ~ ~ B A C H , and our reviewers for critical comments on the manuscript. L.B.B. was supported by a postdoctoral fellowship from the Indiana Business Modernization and Technology Center through the Indiana Institute for Molecular and Cellular Biology. This work was supported by grant GM33291
from the National Institutes of Health to M.A.T.M.
L I T E R A T U R E C I T E D
ALTON, A. K., K. FECHTEL, A. L. TERRY, S. B. MEIKLE and M. A. T. MUSKAVITCH, 1988 Cytogenetic definition and morpho- genetic analysis of Delta, a gene affecting neurogenesis in
Drosophila melanogaster. Genetics 118: 235-245.
ALTON, A. K., K. FECHTEL, C. C. KOPCZYNSKI, S. B. SHEPARD, P. J. KWH and M. A. T . MUSKAVITCH, 1989 Molecular genetics of Delta, a locus required for ectodermal differentiation in Drosophila. Dev. Genet. 10: 261-272.
ARTAVANIS-TSAKONAS, S., and P. SIMPSON, 199 1 Choosing a cell fate: a view from the Notch locus. Trends Genet. 7: 403-408.
BRAND, M . , and J. A. CAMPOS-ORTEGA, 1990 Second-site modi- fiers of the split mutation of Notch define genes involved in neurogenesis in Drosophila melanogaster. Roux's Arch. Dev. Biol. 198: 275-285.
CAMPOS-ORTEGA, J. A., and Y. N. JAN, 1991 Genetic and molec- ular bases of neurogenesis in Drosophila melanogaster. Annu. Rev. Neurosci. 1 4 399-420.
978 L. B. Bender, P. J. Kooh and M. A. T. Muskavitch for a highly selective RNA transport system and its role in
establishing the dorsoventral axis of the Drosophila egg. De- velopment 114: 653-66 l .
CORBIN, V., A. M. MICHELSON, S. M. ABMAYR, V. NEEL, E. ALCAMO, T. MANIATIS and M. W. YOUNG, 1991 A role for the Dro- sophila neurogenic genes in mesoderm differentiation. Cell 67:
31 1-323.
CRAYMER, L., 1984 Drosophila Inform. Serv. 6 0 234-236.
DAVIS, C. G . , 1990 The many faces of epidermal growth factor repeats. New Biol. 2: 410-419.
DE LA CONCHA, A., U . DIETRICH, D. WEIGEL and J. A. CAMPOS- ORTEGA, 1988 Functional interactions of neurogenic genes of Drosophila melanogaster. Genetics 118: 499-508.
DIETRICH, U., and J. A. Campos-Ortega, 1984 The expression of neurogenic loci in imaginal epidermal cells of Drosophila mela- nogaster. J. Neurogenet. 1: 315-332.
FASANO, L., and S. KERRIDGE, 1988 Monitoring positional infor- mation during oogenesis in adult Drosophila. Development
FEHON, R. G . , P. J. KWH, I. REBAY, C. L. REGAN, T . Xu, M. A. T . Muskavitch and S. ARTAVANIS-TSAKONAS, 1990 Molecular interactions between the protein products of the neurogenic loci Notch and Delta, two EGF-homologous genes in Drosophila. Cell 61: 523-534.
GORIELY, A., N. DUMONT, C. DAMBLY-CHAUDI~RE and A. GHYSEN,
199 1 The determination of sense organs in Drosophila: effect of neurogenic mutations in the embryd. Development 113:
1395-1404.
GROSSNIKLAUS, U., H. J. BELLEN, C. WILSON and W. J. GEHRING,
1989 P-element mediated enhancer detection applied to the study of oogenesis in Drosophila. Development 107: 189-200.
HAENLIN, M., B. KRAMATSCHEK and J. A. CAMPOS-ORTEGA,
1990 The pattern of transcription of the neurogenic gene
Delta of Drosophila melanogaster. Development 110: 905-9 14.
HEIMER, G. V., and C. E. D. TAYLOR, 1974 Improved mountant for immunofluorescence preparations. J. Clin. Pathol. 27: 254.
HEITZLER, P., and P. SIMPSON, 1991 The choice of cell fate in the epidermis of Drosophila. Cell 64: 1083-1092.
JURCENS, G., E. WIESCHAUS, C. NUSSLEIN-VOLHARD and H. KLUD-
INC, 1984 Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster. Roux's Arch. Dev. Biol. 193: 283- 295.
KIDD, S., M. R. KELLEY and M. W. YOUNG, 1986 Sequence of the
Notch locus of Drosophila melanogaster: relationship of the en- coded protein to mammalian clotting and growth factors. Mol. Cell. Biol. 6 3094-3108.
KING, R. C., 1970 Ovarian Development in Drosophila melanogaster.
Academic Press, New York.
KWH, P. J., R. G. FEHON and M. A. T . MUSKAVITCH,
1993 Implications of dynamic patterns of Delta and Notch expression for cellular interactions during Drosophila devel- opment. Development (in press).
KOPCZYNSKI, C. C., 1991 A molecular characterization of the Drosophila neurogenic gene Delta. Ph.D. Thesis, Indiana Uni- versity, Bloomington.
KOPCZYNSKI, C. C., and M. A. T. MUSKAVITCH, 1989 Complex spatio-temporal accumulation of alternative transcripts from the neurogenic gene Delta during Drosophila embryogenesis. Development 107: 623-636.
KOPCZYNSKI, C. C., A. K. ALTON, K. FECHTEL, P. J. KOOH and M. A. T . MUSKAVITCH, 1988 Delta, a Drosophila neurogenic gene, is transcriptionally complex and encodes a protein related to blood coagulation factors and epidermal growth factor of vertebrates. Genes Dev. 2: 1723-1735.
LEHMANN, R., F. J I M ~ N E Z , U. DIET RICH^^^ J. A. CAMPOS-ORTEGA, 104 245-253.
1983 On the phenotype and development of mutants of early neurogenesis in Drosophila melanogaster. Roux's Arch. Dev. Biol. 192: 62-74.
MANSEAU, L. J., and T . SCHUPBACH, 1989 cappuccino and spire:
two unique maternal-effect loci required for both the anterio- posterior and dorsoventral patterns of the Drosophila embryo. Genes Dev. 3: 1437-1452.
PARKS, A. L., and M. A. T . MUSKAVITCH, 1993 Delta function is required for bristle organ determination and morphogenesis in Drosophila. Dev. Biol. (in press).
PARKS, S., and A. SPRADLING, 1987 Spatially regulated expression of chorion genes during Drosophila oogenesis. Genes Dev. 1:
497-509.
REBAY, I., R. J. FLEMING, R. G. FEHON, L. CHERBAS, P. CHERBAS and S. ARTAVANIS-TSAKONAS, 1991 Specific EGF repeats of Notch mediate interactions with Delta and Serrate: implications for Notch as a multifunctional receptor. Cell 67: 687-699.
RUOHOLA, H., K. A. BREMER, D. BAKER, J. R. SWEDLOW, L. Y. J A N
and Y. N. JAN, 1991 Role of neurogenic genes in establish- ment of follicle cell fate and oocyte polarity during oogenesis in Drosophila. Cell 66: 433-449.
SCHUPBACH, T., 1987 Germ line and soma cooperate during oogenesis to establish the dorsoventral pattern of egg shell and embryo in Drosophila melanogaster. Cell 49: 699-707.
SHELLENBARGER, D. L., and J. D. MOHLER, 1978 Temperature- sensitive periods and autonomy of pleiotropic effects of l(l)NL"',
a conditional Notch lethal in Drosophila. Dev. Biol. 62: 432- 446.
SHEPARD, S. B., S. A. BROVERMAN and M. A. T . MUSKAVITCH,
1989 A tripartite interaction among alleles of Notch, Delta
and Enhancer ofsplit during imaginal development of Drosoph- ila melanogaster. Genetics 122: 429-438.
SKEATH, J. B., and S. B. CARROLL, 1991 Regulation of achaete- scute gene expression and sensory organ pattern formation in the Drosophila wing. Genes Dev. 5: 984-995.
SKEATH, J. B., and S. B. CARROLL, 1992 Regulation of proneural gene expression and cell fate during neuroblast segregation in the Drosophila embryo. Development 114: 939-946.
SPRADLING, A., 1991 Developmental genetics of oogenesis, in
Drosophila Development, edited by M. BATE and A. MARTINEZ- ARIAS. Cold Spring Harbor Laboratory, Cold Spring Harbor Press, N.Y.
THEURKAUF, W. E., S. SMILEY, M. L. WONG and B. M. ALBERTS,
1992 Reorganization of the cytoskeleton during Drosophila oogenesis: implications for axis specification and intercellular transport. Development 115: 923-936.
VASSIN, H., J. VIELMETTER and J. A. CAMPOS-ORTEGA,
1985 Genetic interactions in early neurogenesis of Drosophila melanogaster. J. Neurogenet. 2: 291-308.
V ~ S I N , H., J. VIELMETTER and J. A. CAMPOS-ORTEGA, 1987 The neurogenic gene Delta of Drosophila is expressed in neurogenic territories and encodes a putative transmembrane protein with EGF-like repeats. EMBO J. 6 3431-3440.
WHARTON, K . A,, K. M. JOHANSEN, T. XU and S. ARTAVANIS- TSAKONAS, 1985 Nucleotide sequence from the neurogenic locus Notch implies a gene product that shares homology with proteins containing EGF-like repeats. Cell 43: 567-58 1 .
Xu, T . , I. REBAY, R. J. FLEMING, T. N. SCOTTCALE and S. ARTA- VANIS-TSAKONAS, 1990 The Notch locus and the genetic cir- cuitry involved in early Drosophila neurogenesis. Genes Dev. 4: 464-475.
Xu, T., L. CARON, R. G. FEHON and S. ARTAVANIS-TSAKONAS,
1992 The involvement of the Notch locus in Drosophila oo-
genesis. Development 115: 91 3-922.