University of Pennsylvania
ScholarlyCommons
Publicly Accessible Penn Dissertations
1-1-2015
Micrornas and the Sex Specific Development of the
Neonatal Brain: A Point of Vulnerability to the
Programming Effects of Prenatal Stress
Christopher Morgan
University of Pennsylvania, [email protected]
Follow this and additional works at:
http://repository.upenn.edu/edissertations
Part of the
Developmental Biology Commons
This paper is posted at ScholarlyCommons.http://repository.upenn.edu/edissertations/1098
For more information, please [email protected].
Recommended Citation
Morgan, Christopher, "Micrornas and the Sex Specific Development of the Neonatal Brain: A Point of Vulnerability to the Programming Effects of Prenatal Stress" (2015).Publicly Accessible Penn Dissertations. 1098.
Micrornas and the Sex Specific Development of the Neonatal Brain: A
Point of Vulnerability to the Programming Effects of Prenatal Stress
Abstract
Epidemiological studies have linked prenatal stress to increases in the incidence of neurodevelopmental disorders, including schizophrenia and autism spectrum disorders, associations that are often sex-dependent. In addition, biological sex is a strong predictor of many aspects of these disorders, including incidence, presentation, and therapeutic outcomes. While many factors contribute to these effects, sex-specific responses to fetal antecedents during key developmental windows are likely involved. The male brain is organized in a sex specific manner by a surge of testes-synthesized testosterone during the perinatal period. In appropriate cell populations this testosterone is converted to estrogen by a neuronal-specific aromatase where it acts through estrogen receptors to masculinize the neural substrate. While the primary effector, estrogen, is shared, the cellular processes responsible for this divergent development vary widely across brain regions. miRNAs, with their ability to regulate the expression of hundreds of genes, may be an exciting and novel regulatory mechanism poised to translate this estrogen signal into brain region-specific responses. The work in this dissertation identifies sexual differentiation of the brain as a point of sex-specific vulnerability to the multigenerational programming effects of early prenatal stress. Paternal (F1) prenatal stress exposure attenuates the perinatal testosterone surge, leading to dysmasculinized physiology, including increased stress sensitivity, in second-generation (F2) male offspring. Further, we reveal a novel role for the miRNA
environment in programming the neurodevelopmental effects of paternal stress exposure and, more generally, in organizing the sexually dimorphic brain. Finally, we empirically map miRNA recognition elements across the transcriptome of the neonatal hypothalamus by Argonaute HITS-CLIP, and identify a network of genes targeted by organizational estrogen with functional relevance to sexual differentiation of the brain. Together these findings point to a developmental window of susceptibility during which the programming effects of early prenatal stress exposure may manifest. As such, identifying sex-specific developmental processes affected during this window, such as the dynamic changes in the miRNA environment we have highlighted, may lead to novel therapeutic targets or biomarkers predictive for neurodevelopmental disorders.
Degree Type
Dissertation
Degree Name
Doctor of Philosophy (PhD)
Graduate Group
Pharmacology
First Advisor
Tracy L. Bale
Keywords
Developmental Programming, Sex Differences
Subject Categories
Developmental Biology
i
MICRORNAS AND THE SEX SPECIFIC DEVELOPMENT OF THE NEONATAL BRAIN:
A POINT OF VULNERABILITY TO THE PROGRAMMING EFFECTS OF PRENATAL
STRESS
Christopher P. Morgan
A DISSERTATION
in
Pharmacology
Presented to the Faculties of the University of Pennsylvania
in
Partial Fulfillment of the Requirements for the
Degree of Doctor of Philosophy
2015
Supervisor of Dissertation Graduate Group Chairperson
_____________________ _____________________
Tracy L. Bale, Ph.D. Julie A. Blendy, Ph.D.
Professor of Neuroscience Professor of Pharmacology
Dissertation Committee
Teresa M. Reyes, Ph.D., Research Assistant Professor of Pharmacology
Zhaolan Zhou, Ph.D., Assistant Professor of Genetics
Olivier Berton, Ph.D., Assistant Professor of Neuroscience in Psychiatry
ii
MICRORNAS AND THE SEX SPECIFIC DEVELOPMENT OF THE NEONATAL BRAIN: A POINT OF VULNERABILITY TO THE PROGRAMMING EFFECTS OF PRENATAL STRESS
COPYRIGHT 2015
iii
ACKNOWLEDGMENT
I had no expectations of staying in graduate school this long, but I am glad that I did. It’s
been a privilege to work with and around such a motivated, intelligent, and creative
group of people over the last 7 years. It is only through their help, encouragement, and
camaraderie that I can leave today with a PhD attached to my name.
Above all, my mentor, Tracy Bale, deserves many thanks for her commitment to my
scientific training. She continued be my advocate despite all the evidence I gave her
against me—at least 534 unanswered emails, countless meetings in her office about
paper drafts I hadn’t yet started, last-minute morning-of thesis committee preparations,
and a 42”x56” poster in full black and white at SfN 2012. She also financed this very
expensive adventure with no less than four concurrent NIH grants. No one can sell a
scientific story with visual statistics like she can, and I’ve learned greatly from her
example. (Really Tracy, no one could have been a more supportive mentor…Thank you)
My fellow graduate students:
- Greg Dunn, whose unimaginable success in neuroscience art is at once inspiring
and completely paralyzing, has been my best friend inside and outside the lab. Our
shenanigans defined my graduate career, and I can only hope to follow his example
into a land far, far away from academic science.
- Alexis Gerber Howerton decorated my bench with lights every year at Christmastime,
a symbol of the spirit and general good cheer she added to my life. Her exploits in
iv
- Ali Buch Rodgers’s edits were essential to all of my abstracts, and her puppy
dog-like loyalty and admiration were rivaled only by Maggie May, and equally
undeserved.
The lab postdocs:
- Dr. Chris paved the way for the secret mid-day gym break that has saved my sanity.
- Katie Morrison’s skeptical face in lab meetings kept me honest and humbled.
- Stefanie Bronson alone treated my sharp embryo-only surgical tools with respect.
The new lab members—Jen, Eldin, and Bridget Nugent—accepted my oddball schedule
at face value when they joined and haven’t asked too many probing questions. All of the
techs—Cindy, Chris Howard, Jess, that guy who went to Harvard Med School whose
name I can’t remember, and also Mufei—were extra hands when I needed them, knew
how to rush order reagents when I really needed them, and spent their valuable time
cleaning up the mess I made with the SRY mouse colony.
Thanks are also due to Dolores, who left me sleeping undisturbed in the conference
room far more times than I’d like to admit, to all the former lab members, notably Bridget
Mueller, who found just enough significance stars in the EPS model on the Barnes Maze
to doom me to nearly a decade of frustratingly subtle and disappearing phenotypes, and
to all of the ULAR technicians who could not be trusted with changing cages and in
doing so commandeered innumerable hours of my life.
And finally, to all the musicians who contributed to the soundtrack of The Bale Lab, and
to the C57/Bl6:129 hybrid mouse strain with its imperfect balance of stress reactivity and
v
ABSTRACT
MICRORNAS AND THE SEX SPECIFIC DEVELOPMENT OF THE NEONATAL BRAIN:
A POINT OF VULNERABILITY TO THE PROGRAMMING EFFECTS OF PRENATAL
STRESS
Christopher P. Morgan
Tracy L. Bale
Epidemiological studies have linked prenatal stress to increases in the incidence of
neurodevelopmental disorders, including schizophrenia and autism spectrum disorders,
associations that are often sex-dependent. In addition, biological sex is a strong
predictor of many aspects of these disorders, including incidence, presentation, and
therapeutic outcomes. While many factors contribute to these effects, sex-specific
responses to fetal antecedents during key developmental windows are likely involved.
The male brain is organized in a sex specific manner by a surge of testes-synthesized
testosterone during the perinatal period. In appropriate cell populations this testosterone
is converted to estrogen by a neuronal-specific aromatase where it acts through
estrogen receptors to masculinize the neural substrate. While the primary effector,
estrogen, is shared, the cellular processes responsible for this divergent development
vary widely across brain regions. miRNAs, with their ability to regulate the expression of
hundreds of genes, may be an exciting and novel regulatory mechanism poised to
translate this estrogen signal into brain region-specific responses. The work in this
dissertation identifies sexual differentiation of the brain as a point of sex-specific
vi
Paternal (F1) prenatal stress exposure attenuates the perinatal testosterone surge,
leading to dysmasculinized physiology, including increased stress sensitivity, in
second-generation (F2) male offspring. Further, we reveal a novel role for the miRNA
environment in programming the neurodevelopmental effects of paternal stress
exposure and, more generally, in organizing the sexually dimorphic brain. Finally, we
empirically map miRNA recognition elements across the transcriptome of the neonatal
hypothalamus by Argonaute HITS-CLIP, and identify a network of genes targeted by
organizational estrogen with functional relevance to sexual differentiation of the brain.
Together these findings point to a developmental window of susceptibility during which
the programming effects of early prenatal stress exposure may manifest. As such,
identifying sex-specific developmental processes affected during this window, such as
the dynamic changes in the miRNA environment we have highlighted, may lead to novel
vii
TABLE OF CONTENTS
ACKNOWLEDGMENT-...-III!
ABSTRACT-...-V!
TABLE-OF-CONTENTS-...-VII!
LIST-OF-TABLES-...-X!
LIST-OF-ILLUSTRATIONS-...-XI!
CHAPTER-1:-GENERAL-INTRODUCTION-...-1!
What is a Neurodevelopmental Disorder? ... 1
!
The Multifactorial Etiology of Neurodevelopmental Disorders ... 2
!
Transgenerational Effects ... 4
!
Modeling the Neurodevelopmental Programming Effects of Prenatal Stress ... 5
!
Sexual Differentiation of the Male Brain ... 7
!
microRNA biogenesis and function ... 9
!
Overview of Dissertation ... 11
!
viii
Introduction ... 13
!
Materials and Methods ... 15
!
Results ... 23
!
Discussion ... 27
!
Figures and Legends ... 32
!
Tables ... 40
!
CHAPTER-3:-SEX-DIFFERENCES-IN-MICRORNA>MRNA-NETWORKS:-NOVEL-
EPIGENETIC-PROGRAMMING-MECHANISMS-IN-THE-SEXUALLY-DIMORPHIC-NEONATAL-HYPOTHALAMUS-...-42!
Introduction ... 42
!
Materials and Methods ... 44
!
Results ... 48
!
Discussion ... 53
!
Figures and Legends ... 65
!
Tables ... 75
!
CHAPTER-4:-GENERAL-DISCUSSION-...-78!
G x E x D x (S) ... 78
!
ix
Transgenerational Epigenetic Impact of Prenatal Stress ... 80
!
Importance of Modulators of Gene Expression in Sexual Differentiation. ... 83!
Estrogen Regulation of the Neonatal Brain miRNA ... 86!
Estrogen-responsive miRNAs Regulate Networks of Genes Functionally Relevant to Sexual Differentiation of the Brain ... 87!
Conclusion ... 89!
x
LIST OF TABLES
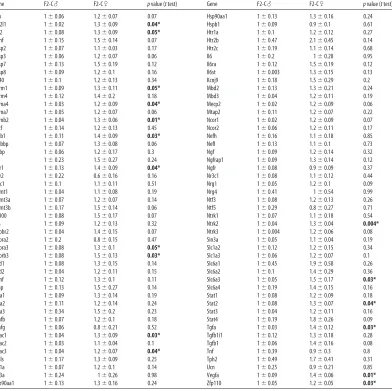
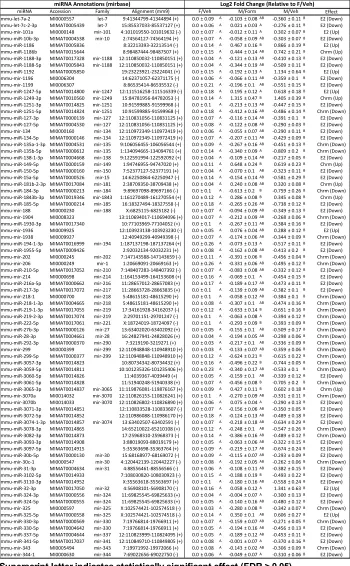
Table 2-1.PN1 brain gene expression assayed by Custom Taqman qRT-PCR Array for
genes important in neurodevelopment……...40
Table 2-2. PN1 brain expression of statistically significant miRNAs assayed by Taqman
qRT-PCR Array……...41
xi
LIST OF ILLUSTRATIONS
Figure 2-1. Second-generation males from the paternal stress lineage (F2-S) show
dysmasculinized brain gene expression and miRNA expression patterns on
postnatal day 1 (PN1) ... 33
!
Figure 2-2. Analyses of physiological and behavioral measures in adult
second-generation males from the paternal stress lineage (F2-S ) show a similar
dysmasculinized physiology and stress-sensitive phenotype as their sires ... 36
!
Figure 2-3. Aromatase inhibition dramatically dysmasculinizes the neonatal brain
miRNA environment during the perinatal sensitive period ... 38
Figure 3-1. The miRNA environment of the neonatal (PN2) hypothalamus is sexually
dimorphic……...65
Figure 3-2. A population of sex-biased miRNAs in the neonatal (PN2) hypothalamus is
dynamically responsive to estrogen...66
Figure 3-3. Estrogen regulation of clustered miRNA genes on chromosome 12...68
Figure 3-4. Ago HITS-CLIP empirically identifies functional miRNA recognition elements
(MREs) across the neonatal (PN2) hypothalamus transcriptome...70
Figure 3-5. Functional miRNA recognition elements (MREs) are present in the mRNAs
xii
Figure 3-6. Estrogen down-regulated miRNAs target a network of genes enriched for
1
CHAPTER 1: General Introduction
Portions of this have appeared in Biology of Sex Differences (2012), 3(1), 22-30 and
Hormones and Behavior (2011), 59(3), 290–295.
What is a Neurodevelopmental Disorder?
“Neurodevelopmental” (and/or “neuropsychiatric”) disorders are inherently
difficult to study because “a lot can go wrong in the development of a human brain”
(Mitchell, 2007). Just trying to define what is and what is not a neurodevelopmental
disorder is difficult. It almost seems that the easiest way to define “neurodevelopmental
disorders” is to say that they are complex. But there does seem to be a subset of
diseases/disorders, including schizophrenia, autism spectrum disorders (ASDs), ADHD,
bipolar disorder, and major depressive disorder (MDD), to which the label is often
applied, and which share two general characteristics: their etiologies are multifactorial,
and these factors interact to give rise to diverse, and often overlapping, phenotypes
(Cristino et al., 2013).Efforts to develop intervention strategies for neurodevelopmental
diseases generally focus on one of these shared characteristics; investigating the
mechanisms of development of a disease (its etiology), or studying the mechanisms
responsible for the expression of the disease phenotype (its pathophysiology). At the
most basic level, the questions explored in this dissertation arise from the first of these
2
The Multifactorial Etiology of Neurodevelopmental Disorders
Recent estimates of the genetic contribution, or heritability, to the etiology of
neurodevelopmental disorders varies from ~90% and ~80% in ASDs and schizophrenia
to ~40% in MDD (Sullivan, Neale, & Kendler, 2000). In the case of schizophrenia, the
lack of any classic Mendelian patterns of inheritance was noted early, and a polygenic
model was proposed (Shields & Gottesman, 1967). At the same time, while studies of
identical twins had demonstrated that though schizophrenia was certainly heritable, this
heritability was incomplete (Shields, Slater, & Gottesman, 1967). Therefore, regardless
of the nature of the genetic contribution, these factors were incompletely penetrant.
Independently, environmental factors, such as social class, had been linked to an
increased prevalence of schizophrenia (Turner & Wagenfeld, 1967). To account for the
fact that no single causal factor was sufficient to account for schizophrenia, Gottesman
and Shields proposed a Genes x Environment (GxE) model that highlighted the
importance of the interaction between a “stressor” (environment) and “stressee”
(genetic) (Gottesman & Shields, 1973). This model is broadly applicable to
neurodevelopmental disorders, and these interactions go both ways; the heritability of
neurodevelopmental disorders is incomplete, and at the same time, there is
heterogeneity in the responses of different individuals to a specific environmental risk
factor (Caspi & Moffitt, 2006).
The developmental origins of adult disease theory, or the Barker hypothesis,
addressed an important remaining question: how do GxE interactions produce disease
phenotypes observed in adults (Barker,!1995)? The core principle of the hypothesis is the
3
specific processes are plastic and responsive to environmental influences (Barker,
2007). This plasticity provides an opportunity for an organism to develop phenotypes
that are most adaptive in a specific environment.For most tissues early development,
and more specifically prenatal development, is the point at which this plasticity is most
apparent. There is a rich epidemiological literature characterizing the association of
maternal stress during periods of pregnancy with premature birth, low birth weight, birth
defects, and altered neurodevelopment. Many retrospective and prospective clinical
reports have linked prenatal stressors including natural and manmade disasters,
bereavement, unwantedness of pregnancy, and reported levels of maternal anxiety or
depression with an increased incidence of neurodevelopmental disorders, including
depression, anxiety, schizophrenia, and autism in the offspring (Beversdorf et al., 2005;
Elias et al., 2004; Khashan et al., 2008; Kinney, Miller, Crowley, Huang, & Gerber, 2008;
Myhrman, Rantakallio, Isohanni, Jones, & Partanen, 1996; Selten, 1999; van Os &
Selten, 1998; Watson, Mednick, Huttunen, & Wang, 1999).
Fetal development is itself highly dynamic, generating a constantly changing set
of tissue targets that can be affected by a prenatal insult. Therefore, it is unlikely that
neurodevelopment would be uniformly susceptible to programming across gestation. For
example, in studies in which maternal stress was initially linked to schizophrenia
development, the importance of the timing of the insult was found to be the major
determinant in disease risk where spousal death during early or late pregnancy
significantly increased risk of offspring schizophrenia (Huttunen & Niskanen, 1978).
4
during the first trimester were 67% more likely to develop schizophrenia then offspring of
unaffected mothers (van Os & Selten, 1998; Khashan et al., 2008).
In addition to the temporal specificity of maternal insult in offspring outcome, fetal
sex has also been identified as an important factor in determination of disease
vulnerability. For instance, an increased risk of schizophrenia has been reported for
offspring of mothers exposed during their first trimester of pregnancy to the 1940
invasion of The Netherlands (van Os & Selten, 1998). However, the study found an
interesting interaction between offspring sex and timing of the stress exposure such that
male but not female offspring exposed in the second trimester to the invasion were also
at an increased risk of schizophrenia as adults. It has been proposed this effect may be
due to the slower rate of cortical development in males leading to an extended window
of sensitivity to the insult (Weinstock, 2007). A similar relationship was identified in
18-year-old students exposed to a severe earthquake in utero where there was found to be
an overall effect of the maternal stress to increase offspring depressive symptoms, but
males exposed to the earthquake during the second trimester showed the greatest
levels of severe depression (Watson et al., 1999). This again supports both a temporal
specificity as well as an involvement of offspring sex in the susceptibility and/or
presentation of disease.
Transgenerational Effects
The heritability of neurodevelopmental disorders may derive not only from
5
important window during which the environment can program changes in
neurodevelopment. It may also serve as a time when the environment can program
phenotypes that persist transgenerationally. A transgenerational impact specific to
maternal stress has not yet been reported in human studies. However, because
enduring malnutrition is inherently stressful, it may be difficult to determine whether
transgenerational metabolic effects that develop as a result of famine are a
consequence of stress or malnutrition. In addition to metabolic phenotypes, studies of
the Dutch Hunger Winters revealed consequences of exposure on neurodevelopment,
including stress sensitivity and risk of schizophrenia (Brown & Susser, 2008).
Interestingly, second generation female offspring had overall poorer health as adults
when their mothers were exposed to famine in utero, and though the statistical power of
the study was insufficient to detect effects on specific phenotypes, outcomes associated
with mental health were included in their definition of “health” (Painter et al., 2008).
These findings highlight two aspects of transgenerational effects that can be influenced
by sex: 1) traits may be transmitted specifically along the maternal or paternal lineage,
and 2) traits may be inherited or expressed by only male or female offspring.
Modeling the Neurodevelopmental Programming Effects of Prenatal Stress
Neurodevelopmental disorders are diagnosed based on a patient’s
symptomology, but individuals with the same disorder often display very different
symptoms. At the same time, there is a high degree of overlap in phenotype between
different disorders; an individual’s diagnosis may even change over their lifetime
6
etiology. Several genetic variants originally associated with schizophrenia were later
linked to ASD or bipolar disorder (Sullivan et al., 2012; Guilmatre et al., 2009). In
addition, there is significant familial comorbidity between schizophrenia, bipolar
disorders, and MDD (Steinhausen et al., 2009). While these characteristics increase the
complexity of designing studies in humans, they suggest that studies in animal models
have broad applicability across neurodevelopmental disease. After all, a mouse cannot
be diagnosed with schizophrenia or ASD. Instead, we investigate intermediate
phenotypes, or endophenotypes, thought to underlie disease pathophysiology, which are
shared across neurodevelopmental disorders (Walters & Owen, 2007).
Animal models allow us to establish causal relationships between specific
aspects of the early-life environment and these endophenotypes. In addition, we can use
animal models to identify the molecular mechanisms through which environmental
factors program disease resilience or vulnerability. In mice, rats, guinea pigs, and
nonhuman primates prenatal stress has been shown to result in stress dysregulation,
anxiety and depressive-like behaviors, and cognitive deficits, all endophenotypes
associated with neuropsychiatric disease (Lemaire et al., 2000; Schneider et al., 2002;
Weinstock, 2001; Darnaudery & Maccari, 2008; Kapoor & Matthews, 2005; Kapoor et al.,
2009; Mueller & Bale, 2007, 2008). As in human studies, specific offspring outcomes
have varied depending upon stressors utilized, timing of the stress event during
pregnancy, and sex of the offspring.
Our lab has utilized a chronic variable stress (CVS) paradigm in mice to compare
the programming effects of maternal stress experience across early, mid or late
7
only defined the temporal specificity of prenatal stress effects, but also identified
sex-specific offspring outcomes. CVS early in pregnancy (EPS) increased immobility in both
the tail suspension and forced swim tests specifically in male offspring (Mueller & Bale,
2008). EPS exposure also affected the programming of offspring stress neurocircuitry,
as prenatally stressed males exhibited a more female-typical elevated peak
corticosterone levels in response to a restraint stress. This effect was associated with
elevated limbic corticotropin-releasing factor, reduced hippocampal glucocorticoid
receptor expression, and with corresponding alterations in CpG dinucleotide methylation
within the promoters of these two genes. There were also sex-specific effects of EPS on
performance in the Barnes maze, a spatial learning and memory task (Mueller & Bale,
2007). Early CVS exposed males took significantly longer then control males to
complete the task, and utilized more female-typical learning strategies. Overall, EPS
males displayed dysmasculinized patterns of stress physiology, behavior, and cognitive
performance, pointing to a potential disruption in normal brain masculinization.
Sexual Differentiation of the Male Brain
Biological sex is a strong predictor of many aspects of neurodevelopmental
disorders, including prevalence, presentation, and therapeutic outcomes. Sex
differences in health outcomes are not limited to neurodevelopmental disease; instead,
the importance of sex has been highlighted in immune-related diseases, many cancers,
and coronary heart disease as examples (Goldstein, Handa, & Tobet, 2014; Ober et al.,
2008; Voskuhl, 2011; Yeh & Chen, 2010). Recognition of these sex-biases in disease is
8
cells and tissues. The male brain is organized in a sex specific manner by a surge of
testes-synthesized testosterone during the perinatal period. In appropriate cell
populations this testosterone is converted to estrogen by a neuronal-specific aromatase
where it acts through estrogen receptors to masculinize and defeminize the neural
substrate. This organized substrate is then activated by adult gonadal hormone levels to
express an appropriate sex-specific behavior and physiology (Arnold & Gorski,1984;
McCarthy et al., 2009b; Phoenix et al., 1959).
In addition to programming fundamental sex specific behaviors like reproductive
and territorial behaviors in rodents, organizational testosterone establishes sex
differences in stress neurocircuitry. For example, PN1 gonadectomy of male rats results
in adults with elevated corticosterone response to restraint stress, an effect that can be
reversed with testosterone treatment from PN1-PN5 (Bingham & Viau, 2008). Similarly,
a PN1 injection of testosterone in female rats reduces adult corticosterone responses to
a noise stress (Seale et al., 2005). In addition to effects on HPA axis sensitivity, our lab
has shown that perinatal testosterone can masculinize stress coping strategies in female
mice. In the marble burying test males generally show active coping strategies (burying)
while females show more passive strategies (avoidance). Perinatal testosterone
masculinizes female performance leading to a greater number of marbles buried
compared to vehicle treated females (Goel & Bale, 2008b).
NCBI's RefSeq project has currently annotated approximately 20,000 protein
coding genes in both the human and mouse genome. Of these, fewer then 200, or about
1%, are on the Y chromosome and not shared by males and females (Pruitt et al., 2014).
9
component of the sexual differentiation of a tissue. Evidence for this can be found in the
extent to which sex differences exist in the transcriptomes of various tissues, with
55-72% of the active genes in muscle, adipose, and liver tissue displaying sex-biased
expression (Yang, 2006). The ability of miRNAs to regulate a large number of genes with
a high degree of specificity and control makes them perfectly poised to play key roles in
sexually dimorphic programs of gene expression.
microRNA biogenesis and function
miRNAs are small non-coding RNAs that regulate post-transcriptional gene
expression by affecting the stability or translational efficiency of specific mRNA targets.
The majority of miRNAs are organized in clusters within the genome, and are
co-transcribed as single long poly-cistronic primary transcripts (pri-miRNAs) that are
multiple kilobases in length (Bartel, 2004). The transcription of these pri-miRNAs is RNA
polymerase II dependent, and regulated by the same mechanisms as mRNAs (Krol et
al., 2010). Also similar to mRNAs, pri-miRNAs undergo 5’ capping and 3’
polyadenylation (Cai et al., 2004) . Portions of the pri-miRNA folds back on itself to form
a distinct stem-loop structure. In the nucleus, a Drosha-containing microprocessor
cleaves the pri-miRNA at the base of this ‘stem,’ generating a 60–70 bp precursor
(pre)-miRNA (Bartel, 2004). Other (pre)-miRNAs, termed mirtrons, are located within introns of
mRNAs, and are co-transcribed with their host gene (Krol et al., 2010; Ruby et al.,
2007). Mirtrons bypass Drosha processing, and instead use mRNA splicing machinery to
generate pre-miRNAs (Ruby et al., 2007). In the cytosol, Dicer processes pre-miRNAs
10
a duplex (the guide strand) into the Argonaute-containing RNA-induced silencing
complex (known as the RISC complex). Argonaute proteins act at the interface between
miRNAs and their target mRNAs to mediate the functional consequences of these
interactions (Höck & Meister, 2008). There are four different Argonaute proteins found in
mammals, Argo 1–4, though 60% of miRNA-associated RISC complexes contain Argo 2,
the only Argonaute with endonuclease activity (Höck & Meister, 2008; Wang et al.,
2012). Mature miRNAs guide the RISC complex to the 3’ UTR of mRNAs, providing
target specificity through partial sequence homology, and typically resulting in mRNA
destabilization and degradation (Filipowicz et al., 2008; Guo et al., 2010; Bartel, 2009).
Thus, a typical rule is that with an increase in miRNA expression, you see a concordant
decrease in the target mRNA.
miRNAs act as major components of an integrated gene expression regulatory
mechanism (Hah et al., 2011). One genome-wide bioinformatics study annotated more
than 45,000 conserved miRNA binding sites in the 3’ UTR of 60% of human genes
(Friedman et al., 2009). In addition, each miRNA can directly target more than a hundred
different mRNA targets, making this mode of regulation far-reaching and capable of
profound programmatic effects (Baek et al., 2008; Selbach et al., 2008). For example,
more than 600 distinct mRNA targets were identified by Argonaute immunoprecipitation
following miR-124 overexpression in MCF-7 cells, an immortalized breast cancer cell line
(Hendrickson et al., 2009). Interestingly, the presence of a miRNA does not necessarily
result in the complete absence of expression of target transcripts. In fact, in two separate
proteomic studies the transfection or deletion of a single miRNA (including 1,
11
hundreds of proteins. However, the magnitude of these effects on individual proteins
was modest (1–2 fold changes) effects (Baek et al., 2008; Selbach et al., 2008). These
data support a model of miRNA function proposed by Bartel and Chen, in which they
described a functional group of mRNA targets, termed “tuning targets” that have taken
advantage of the miRNA environment and machinery to develop an additional level of
regulation (Bartel & Chen, 2004). This provides a cell with a greater ability to modulate
gene expression in response to a continuously changing environment.
Overview of Dissertation
The work in this dissertation identifies sexual differentiation of the brain as a point
of sex-specific vulnerability to the multigenerational programming effects of early
prenatal stress. Paternal (F1) prenatal stress exposure attenuates the perinatal
testosterone surge, leading to dysmasculinized physiology, including increased stress
sensitivity, in second-generation (F2) male offspring. Further, we reveal a novel role for
the miRNA environment in programming the neurodevelopmental effects of paternal
stress exposure and, more generally, in organizing the sexually dimorphic brain. Finally,
we empirically map miRNA recognition elements across the transcriptome of the
neonatal hypothalamus and identify a network of genes targeted by organizational
estrogen with functional relevance to sexual differentiation of the brain. Together these
findings point to a developmental window of susceptibility during which the programming
effects of early prenatal stress exposure may manifest. As such, identifying sex-specific
12
the miRNA environment we have highlighted, may lead to novel therapeutic targets or
13
CHAPTER 2: Early Prenatal Stress Epigenetically Programs
Dysmasculinization in Second-Generation Offspring Via the Paternal
Lineage
Christopher P. Morgan and Tracy L. Bale
This work was originally published in J Neurosci (2011), 31(33), 11748–11755.
Introduction
Epidemiological studies have linked prenatal stress to increases in the incidence
of neurodevelopmental disorders, including schizophrenia and autism spectrum
disorders, associations that are often sex-dependent (Huttunen and Niskanen, 1978; van
Os and Selten, 1998; Khashan et al., 2008; Kinney et al., 2008). These diseases often
display sex differences in prevalence, presentation, or therapeutic outcomes (Bale et al.,
2010). While many factors likely contribute to these differences, sex-specific responses
to fetal antecedents are likely involved (Weinstock, 2007).
We have previously identified early gestation as a specific window of sensitivity
during which male mice were susceptible to the programming effects of maternal stress.
These males exhibited physiological and behavioral stress-sensitivity and cognitive
deficits, which are endophenotypes associated with human neuropsychiatric disease. In
addition, these changes reduced or disrupted established sex-differences by
dysmasculinizing male offspring measures of stress responsivity (Mueller and Bale,
2007, 2008). Similar disruptions of sex differences in behavior, morphology, and gene
14
paradigms across multiple species (Ward, 1972; Meisel et al., 1979; Reznikov et al.,
1999; Biala et al., 2010; Kapoor and Matthews, 2005). The organizational/activational
hypothesis of brain development suggests that a surge of gonadal hormones organize
the brain in a sexually dimorphic manner during the perinatal sensitive period. Then in
adulthood, gonadal hormones can activate this organized neurocircuitry to express
appropriate sex-specific behavioral phenotypes, including stress axis responsivity
(Phoenix et al., 1959; Arnold and Gorski, 1984; Seale et al., 2005; Bingham and Viau,
2008). The disruption of sex-differences identified in our model suggests that early
prenatal stress alters the trajectory of neurodevelopment during the perinatal period.
Fetal antecedents likely contribute to adult disease through programming
changes in the epigenome. Examples of this phenomenon are emerging in human
studies. For example, infants with prenatal exposure to maternal depression or anxious
mood exhibited increased glucocorticoid methylation, which was associated with a
heightened cortisol response to a mild stressor (Oberlander et al., 2008). Such
programming effects may transmit to subsequent generations, predisposing offspring to
disease. Animal models have clearly established a role for epigenetics in
transgenerational phenotypic inheritance following exposure to environmental factors
such as maternal stress, diet, and endocrine disruptors (Dunn and Bale, 2009; Franklin
et al., 2010; Guerrero-Bosagna et al., 2010; Skinner, 2011). Importantly, in rodent
models, inheritance of a phenotype through the paternal lineage excludes confounding
effects of the maternal intrauterine environment and postnatal rearing behavior, and
indicates gametic epigenetic transmission (Youngson and Whitelaw, 2008). These
15
neurodevelopmental diseases could be the product of both classic genetic and
non-mendelian, or epigenetic, mechanisms (Maher, 2008; Slatkin, 2009). Therefore, we
utilized our mouse model to determine epigenetic programming effects and the
transmission of a dysmasculinized phenotype to second-generation (F2) male offspring
via the paternal lineage, eventually focusing mechanistically on analysis of the miRNA
environment as a form of transcriptional regulation with broad potential to impact
developmental processes.
Materials and Methods
Animals
All dams bred for gestational stress studies were virgin, experimentally naïve
C57Bl/6:129 F1 hybrid 5 wk old mice purchased from the Jackson laboratory. Offspring
from these breedings were used to generate the second-generation (F2) litters for
transgenerational studies. Justification for using a hybrid background strain in these
studies is related to stress responsivity phenotypes and physiology. C57Bl/6 are
extremely low stress responders and display low levels of maternal care, making them
poor choices for studies focusing on neurodevelopment. While 129 mice are great stress
responders and show high levels of quality maternal care, they frequently lack a fully
formed corpus callosum and are poor performers in behavioral tests, especially learning
and memory tasks. However, the combination of these two strains produces a hybrid
vigor that has served our research well with predictable stress responses, behavioral
16
Bale, 2006, 2007, 2008). Pregnancy was established by confirmation of a copulation
plug (checked for each morning between 7-8 am). Presence of a copulation plug
denoted experimental day 1 for early prenatal stress exposure. The pregnant female
was individually housed, given a cotton nestlet, and randomly assigned to a stress
treatment or control group. Food (Purina Rodent Chow; 28.1% protein, 59.8%
carbohydrate, 12.1% fat) and water was provided ad libitum throughout the study. All
studies were performed according to experimental protocols approved by the University
of Pennsylvania Institutional Animal Care and Use Committee, and all procedures were
conducted in accordance with institutional guidelines.
Early Prenatal Stress (EPS)
Administration of chronic variable stress was performed as described previously
(Mueller and Bale, 2006). Briefly, pregnant dams were randomly assigned to either an
experimental treatment group to receive chronic variable stress during gestation days
1-7, or to a control non-stressed treatment group. Pregnant dams assigned to the stress
group experienced a different stressor on each of the seven days. Stressors included: 36
hrs constant light, 15 min of fox odor exposure (1:10,000 2,4,5-trimethylthiazole, Acros
Organics), novel objects in cage overnight (eight marbles of similar size and color), 5
min restraint stress in a 50 ml conical tube, novel white noise overnight (Sleep machine,
Brookstone), multiple cage changes, and water saturated bedding overnight. These mild
stressors were selected to be non-habituating and to not induce pain. We previously
demonstrated that this chronic variable stress paradigm does not affect maternal food or
water intake, weight gain, gestation length, litter size, or post-partum maternal behaviors
17
Offspring
To determine if the dysmasculinized stress-sensitive phenotype identified in
first-generation (F1) prenatally stressed males could be transmitted through the paternal
lineage to second-generation (F2) offspring, F1 control or prenatally stressed males
were bred with F1 control females to generate F2 control litters (F2-C n = 9 litters, n = 69
total animals) or F2 stress litters (F2-S n = 9 litters, n = 59 total animals). Molecular,
physiological, morphological, and behavioral correlates of the dysmasculinized
stress-sensitive phenotype were then tested in these F2 offspring. All experimental n’s
described throughout the manuscript refer to litter n’s
Postnatal day one (PN1) brain expression analysis
To exploremechanisms through which the F1 dysmasculinized stress-sensitive
phenotype may be programmed in the F2 generation, we examined whole brain gene
expression in male and female neonates at birth, a period critical for the organization of
the sexually dimorphic brain by gonadal hormones. One male and one female neonate
per litter were sacrificed on the day of parturition. Whole brains were dissected, frozen in
liquid nitrogen, and stored at -80°C prior to assay. Neonate sex was confirmed by SRY
genotyping of tail genomic DNA as previously described (Mueller and Bale, 2008).
Whole brains were sonicated in TRIzol reagent (Invitrogen) and total RNA was isolated
according to manufacturer’s protocol.
Custom Taqman qRT-PCR Array
250 ng of total RNA from PN1 brains was reverse transcribed to cDNA using the
18
93 genes important in neurodevelopment were determined using a custom Taqman
array (F2-C n = 4, F2-S n = 3, and F2-C n = 4) (Applied Biosystems). Analysis
was performed using the comparative Ct method. 18S rRNA was used as an
endogenous loading control. Expression levels of each sample were normalized to the
average F2-C expression level.
miRNA Taqman qRT-PCR Array
500 ng of total RNA from PN1 brain was reverse transcribed to cDNA using
Megaplex RT pool A primers and Multiscribe reverse transcriptase (Applied biosystems).
Expression levels of 239 miRNAs were determined using theTaqman Array MicroRNA
card A Array (F2-C n = 5, F2-S n = 3, and F2-C n = 5) (Applied Biosystems).
Analysis was performed using the comparative Ct method. For each sample, the
average of the Ct values of sno135 and sno202 was used as an endogenous loading
control. Expression levels of each sample were normalized to the average F2-C
expression level.
qRT-PCR of predicted miRNA target transcripts
Predicted miRNA targets were identified using the web-based algorithm miRDB
(Wang, 2008; Wang and El Naqa, 2008). Expression of mRNAs that were shared
predicted targets of miRNAs identified as significantly changed by early prenatal stress,
and with potential relevance to disease mechanisms, were assayed by Taqman
qRT-PCR (Applied Biosystems). GAPDH was used as an endogenous loading control.
19
method. Expression levels of each sample were normalized to the average F2-C
expression level.
F2 physiological and behavioral measures
For each test, one female and one male per litter were examined to avoid litter
effects. An investigator blind to animal treatment group and sex performed scoring of
behavioral tests.
Tail Suspension Test (TST)
The TST was performed as previously described to examine active versus
passive stress coping strategies (Steru et al., 1985). Mice were secured to a rod by
adhesive tape placed approximately 1 cm from the tip of the tail, and suspended 50 cm
from the bench-top in a visually isolated area (F2-C n = 7, F2-S n = 7, F2-C n = 8,
and F2-S n = 6). Immobility time, defined as the absence of all movement except for
whisker movement or respiration, was scored for the 6 min test session.
Barnes Maze
As a test with predictable sex-differences in outcome measures of learning and
memory, we conducted the Barnes maze (F2-C n = 7, F2-S n = 7, F2-C n = 9,
and F2-C n = 9) as previously described (Mueller and Bale, 2007). Briefly, the maze
consists of a black circular disk (90 cm in diameter) with 24 holes evenly spaced around
its perimeter. An escape box is located under one of the holes. The location of the
escape box remains constant throughout training. The disk is elevated 70 cm above the
20
perimeter of the maze. All mice were trained on the maze 2 trials/day for 3 days. Trials
within each day were separated by 4 hrs. To begin each trial, the mouse was placed
under a glass beaker in the center of the maze for 15 sec prior to trial start. Latency to
identify the target escape box was determined. Each trial was terminated when the
mouse located and entered the target escape box or after 4 min elapsed. If the mouse
did not successfully locate the target box, the investigator guided the mouse to the target
and a latency to target of 240 sec was assigned.
HPA response to restraint stress
The HPA axis response to acute stress was measured by exposing adult F2 mice
to a 15 min restraint in a 50 ml conical tube (F2-C n = 7, F2-S n = 7, F2-C n = 7,
and F2-C n = 9). Testing occurred 2-5 hrs following lights on. Blood samples were
collected from a tail nick at four time points: 1) time 0, immediately upon removal from
the cage, 2) time 15, immediately after the restraint stress, 3) time 30, following 15 min
recovery in the home cage, 4) time 90, following 75 min recovery in the home cage.
Samples were collected into EDTA-treated tubes, centrifuged, and plasma was stored at
-80°C until corticosterone levels were measured by radioimmunoassay (MP
Biomedicals). The minimum detection limit of the assay was 7.7 ng/ml, and the intrassay
coefficient of variation was 7.1%.
Morphology
One experimentally naïve male and female per litter were sacrificed as adults
(F2-C n = 7, F2-S n = 7, and F2-C n = 9). Anogenital distances and left testis
21
Effects of an aromatase inhibitor on the neonatal brain miRNA
environment
Formestane administration
To examine the influence of organizational estradiol on the neonatal brain miRNA
environment, a separate cohort of C57Bl/6:129 control pups were treated with an
aromatase inhibitor or vehicleon the morning following parturition. Male pups were
randomly assigned to receive 20 µg of formestane (Sigma-Aldrich) in 20 µl sesame oil
with 10% ethanol (n = 7 from 6 litters) or vehicle injections (n = 7 from 6 litters). This
dose, after adapting for differences in rat versus mouse neonate weight, was previously
shown to reduce male hypothalamic estrogen to female levels (Amateau et al., 2004). All
female pups received vehicle injections (n = 8 from 5 litters). Injections were
administered subcutaneously between the shoulders, and the injection site was treated
with New Skin liquid bandage to prevent leakage.
miRNA expression analysis
Pups were sacrificed 24 hrs after treatment (PN2). Whole brains were dissected,
frozen in liquid nitrogen, and stored at -80°C prior to assay. Neonate sex was confirmed
by SRY genotyping of tail genomic DNA as previously described (Mueller and Bale,
2008). Whole brains were sonicated in TRIzol reagent (Invitrogen) and total RNA was
isolated according to manufacturer’s protocol. 500 ng of total RNA from PN2 brain was
reverse transcribed to cDNA using Megaplex RT pool A primers and Multiscribe reverse
transcriptase (Applied Biosystems). Expression levels of 239 miRNAs were determined
using theTaqman Array MicroRNA card A Array (Applied Biosystems). Analysis was
22
values of sno135 and sno202 was used as an endogenous loading control. Expression
levels of each sample were normalized to the average expression level of
vehicle-treated males.
Statistics
An investigator blind to animal treatment group and sex conducted all studies
and analyses. Either only one male and female from a litter was used for a test or assay,
or values were averaged across littermates to control for litter effects where identified in
methods. We tested the null hypothesis that early prenatal stress exposed males could
not transmit their dysmasculinized stress-sensitive phenotype to their second-generation
offspring. Morphological measures and immobility in the TST were analyzed by
one-tailed student’s t-test, testing for effects identified in early prenatal stress exposed (F1)
sires. Barnes maze performance and corticosterone response to restraint was analyzed
by one-way ANOVA for paternal (F1) prenatal stress exposure, and trial or time as
repeated measure, respectively. miRNA and gene target expression in F2-C , F2-S ,
and F2-C PN1 whole brains was analyzed by one-way ANOVA, using student’s t-tests
for post hoc analysis. miRNA expression data in control or formestane-treated brains
were analyzed by hierarchical clustering of samples, using Pearson correlation as a
metric. Treatment effects on individual miRNAs were identified by two-tailed student’s
t-tests. Heatmaps and hierarchical clustering was performed using MultiExperiment
23
Results
F2 gene expression analysis during the perinatal sensitive period
F2 Postnatal day one (PN1) brain gene expression analysis
To exploremechanisms through which the F1 dysmasculinized stress-sensitive
phenotype may be programmed in the F2 generation, we examined brain gene
expression in male and female neonates at birth, a period critical for the organization of
the sexually dimorphic brain by gonadal hormones. Expression data for individual genes
are presented inTable 2-1. These data are also displayed in a heatmap to highlight the
broad similarities in gene expression that exist between F2-S♂ and F2-C♀ relative to
F2-C♂ (Fig. 2-1A). Mean F2-S♂ expression of the 17 genes that display significant
basal sex differences in PN1 whole brain are plotted on a continuum between average
F2-C♂ and F2-C♀ expression (Fig. 2-1B). In F2-S♂, 13 of these 17 genes displayed
expression levels closer to F2-C♀ than to F2-C♂ levels.
To identify potential mediators of the program of dysmasculinized gene
expression, we assayed the F2 PN1 brain expression of aromatase, ERα, and ERβ,
known effectors of masculinization during the perinatal sensitive period (Fig. 2-1C).
While there was no significant effect of group on aromatase [F(2, 10)=0.69; p=0.70] or
ERα levels [F(2, 10)=1.51; p=0.27], ERβ expression was elevated in F2-S♂ compared to
F2-C♂ [F(2, 10)=4.23; p=0.05].
24
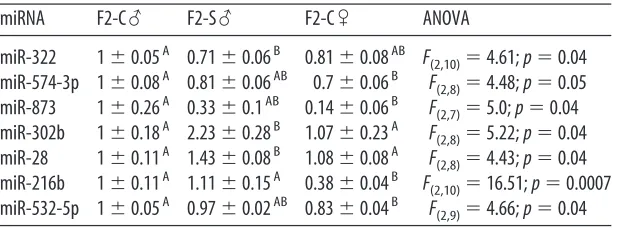
To identify potential alternative mediators of a program of dysmasculinized gene
expression in F2-S males, the F2 PN1 brain miRNA environment was assayed. miRNAs
with expression levels that displayed a statistically significant effect of group are
displayed in Table 2-2. The expression of three of these miRNAs: miR-322, miR-574-3p,
and miR-873, appeared dysmasculinized in F2-S♂(Fig. 2-1D).
To determine if miRNA changes in F2 PN1 brains were associated with altered
expression of target transcripts, expression of genes that were shared predicted targets
of miRNAs identified as significantly changed by early prenatal stress, and with potential
relevance to disease mechanisms, were assayed by Taqman qRT-PCR. β-glycan was
the only predicted target of all three dysmasculinized miRNAs (miR-322, miR-574-3p,
and miR-873). There was significantly greater expression of β-glycan in F2-S♂ then in
F2-C♂ [F(2, 10)=4.99; p=0.03] (Fig. 2-1D). There was also significantly greater expression
of Reep3, the shared predicted target of miR-302b and miR-28, in F2-S♂ (1.23±0.11)
compared to F2-C♀ (0.91±0.06) [F(2, 10)=4.64; p=0.04]. There were no significant
differences in expression of the additional predicted targets of miR-322 and miR-873,
Plxna2 [F(2, 10)=0.72; p=0.51] and Prkar2a [F(2, 10)=0.23; p=0.23], or of the predicted
targets of miR-28 and miR-302b, Unk [F(2, 10)=0.37; p=0.70] and Hif1an [F(2, 10)=0.68;
p=0.53].
Analysis of adult F2 behavior and physiology
25
To assess the degree to which paternal (F1) prenatal stress exposure affects
morphological measures of masculinization, we examined male anogenital distances
and testis weights (Fig. 2-2A and 2-2B). F2-S♂ had reduced anogenital distances
[one-tail t=2.7; p=0.01] and reduced testis weights [one-[one-tail t=1.97; p=0.04].
F2 Adult behavior
To determine if prenatal stressed males (F1) could transmit their
dysmasculinized stress-sensitive phenotype to F2 offspring, we examined F2 adult
performance in the TST and Barnes maze. F2-S♂ spent significantly more time
immobile then F2-C♂ (one-tailed t(12)=1.85, p=0.04) (Fig. 2-2C). There was no
corresponding increase in F2-S♀ immobility relative to F2-C♀ (one-tailed t(12)=0.17,
p=0.44) (Fig. 2-2D). Analyzing Barnes maze performance, there was no statistically
significant between subjects effect of paternal (F1) prenatal stress exposure in males
[F(1, 9)=0.09; p=0.40]. As expected, there was a significant within subjects effect of time
[F(5,5)=6.91; p=0.03] (Fig. 2-2E). In females, there was also no significant
between-subjects effect of paternal (F1) prenatal stress exposure [F(1, 11)=0.03; p=0.57], and again
there was a significant within-subjects effect of time [F(5,7)=4.34; p=0.02] (Fig. 2-2F).
F2 Adult HPA stress axis
To examine the impact of paternal (F1) prenatal stress exposure on F2 offspring
HPA axis sensitivity, we examined corticosterone levels in response to a 15 min restraint
stress. In males, there was no significant between-subjects effect of paternal prenatal
26
significant within-subjects effect of time [F(3,10)=51.0; p<0.0001] (Figure 2-2G). In
females, as in males, there was no significant between-subjects effect of paternal
prenatal stress exposure on corticosterone response [F(1,14)=0.89; p=0.36], though there
was a significant within-subjects effect of time [F(3,12)=33.5; p<0.0001] (Figure 2-2H).
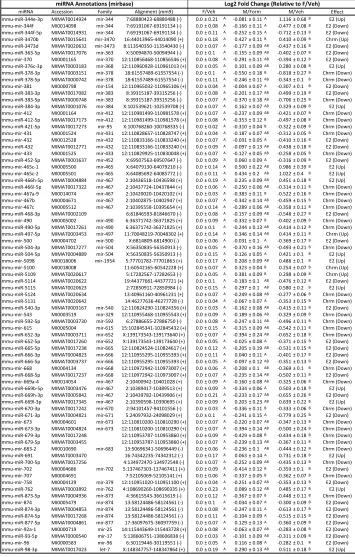
Effects of formestane treatment on the neonatal miRNA environment
To determine the role of organizational gonadal hormones in the regulation of the
neonatal brain miRNA environment, we assayed changes in brain miRNA complement
24 hrs following a PN1 injection of the aromatase inhibitor, formestane. These data are
displayed in a heatmap (Fig. 2-3). Hierarchical clustering analysis using Pearson
correlation as a metric successfully segregated male vehicle samples from
formestane-treated male and female vehicle samples, while it was unable to distinguish between
formestane-treated male and vehicle treated female samples (Fig. 2-3). Formestane
significantly increased expression of miR-143 [t(12)=3.0, p=0.02], miR-152 [t(12)=3.69,
p=0.005], miR-18a [t(12)=2.35, p=0.04], miR-298 [t(12)=2.24, p=0.05], miR-301b [t(12)=2.29,
p=0.04], miR-34a [t(12)=2.65, p=0.03], miR-362-3p [t(12)=2.79, p=0.02], miR-365
[t(12)=2.75, p=0.02], miR-384-3p [t(12)=3.34, p=0.007], miR-448 [t(12)=2.47, p=0.03],
miR-451 [t(12)=2.57, p=0.03], and miR-674 [t(12)=2.38, p=0.04]. Formestane significantly
reduced expression of miR-133b [t(12)=4.16, p=0.002], miR-15a [t(12)=2.23, p=0.05],
27
Discussion
Epidemiological studies have linked sex-biased neurodevelopmental disorders,
including autism and schizophrenia, with prenatal stress (Huttunen and Niskanen, 1978;
van Os and Selten, 1998; Khashan et al., 2008; Kinney et al., 2008). Animal models of
prenatal and postnatal stress have provided insight into sensitive periods and
sex-specific vulnerabilities related to neurodevelopmental disorder etiology (Champagne and
Meaney, 2007; Mueller and Bale, 2007; Kapoor and Matthews, 2008; Mueller and Bale,
2008; Cottrell and Seckl, 2009; Biala et al., 2010; Eiland and McEwen, 2010; Ivy et al.,
2010; Korosi et al., 2010). We previously identified early gestation as a period sensitive
to the sex-specific programming effects of prenatal stress in which male offspring
showed a dysmasculinized phenotype in behavioral and physiological stress measures
as adults (Mueller and Bale, 2007, 2008). As certain disease outcomes persist into
subsequent generations, we examined the paternal transmission and programming of
the prenatal stress induced dysmasculinized phenotype in second-generation (F2)
offspring.
F2 brain gene expression was examined during the perinatal sensitive period to
identifymechanisms of a disruption in masculinization in the F2 male brain. This period
is critical for the organization of the sexually dimorphic brain by gonadal hormones.
Using a custom Taqman qRT-PCR Array for genes involved in neurodevelopment, we
observed a broad shift in expression from a male-typical to a more female-typical pattern
in the F2 male offspring of prenatally stressed sires (F2-S). In F2-S male PN1 brains, 13
of 17 genes with statistical sex differences displayed expression levels closer to F2-C
28
differences previously reported in adult hippocampal gene expression of prenatally
stressed rats, supporting the hypothesis that disrupted masculinization during the
perinatal sensitive period may be a mechanism through which paternal (F1) prenatal
stress exposure impacts F2 offspring development (Biala et al., 2010).
Sex differences in gene expression result from combinations of chromosomal
and hormonal effects. The male brain is organized in a sex-specific manner by a surge
of testosterone during the perinatal sensitive period (Phoenix et al., 1959; McCarthy et
al., 2009a).Testosterone is converted to estradiol by a neuronal-specific aromatase
where it alters gene expression to masculinize and defeminize neurocircuitry through the
estrogen receptors ERα and ERβ. We examined the expression of these primary
effectors to determine if their dysregulation was associated with the broad shift in gene
expression observed in F2-S males. While aromatase expression was unchanged, both
ERα and ERβ appeared upregulated, an effect suggestive of reduced ligand availability
supporting a hypothesis for decreased perinatal testosterone in F2-S males. To identify
potential alternative mediators of the dysmasculinized gene expression in F2-S males,
we examined the PN1 brain miRNA environment. miRNAs are small noncoding RNAs
involved in the post-transcriptional regulation of genes (Bartel, 2009). Interestingly, a
single miRNA may interact with up to a hundred target transcripts, potentially regulating
critical gene families involved in early neurodevelopment. We identified 3 miRNAs
whose expression appeared dysmasculinized in F2-S males, and 2 miRNAs that showed
a significant effect of paternal (F1) prenatal stress. Several of these miRNAs have
known functions in peripheral tissues (Caruso et al., 2010; Ghosh et al., 2010; Qin et al.,
29
we identified predicted gene targets utilizing the web-based algorithm miRDB (Wang,
2008; Wang and El Naqa, 2008). Only one gene, β-glycan (TGFβr3), was a shared
predicted target of all 3 dysmasculinized miRNAs. As would be predicted based on the
reduced expression of miR-322, miR-574-3p, and miR-873, β-glycan expression was
significantly increased in the F2-S male PN1 brain. Beta-glycan is a member of the
TGFβ superfamily expressed in adult brain, pituitary, and gonadal tissues where it acts
as an accessory protein, binding other TGFβ isoforms, such as inhibin A, and increasing
their receptor affinity (Lewis et al., 2000; MacConell et al., 2002). Interestingly, in
pituitary gonadotrophs and gonadal leydig or theca cells, β-glycan is involved in
regulating the release of gonadal hormones (MacConell et al., 2002; Chapman and
Woodruff, 2003; Wiater et al., 2009). As a role for β-glycan in neurodevelopment has not
been identified, our data suggest that it may serve an unappreciated role in the
organization of the sexually dimorphic brain.
As an additional physiological marker programmed by perinatal testosterone,
adult male anogenital distances were measured (Scott et al., 2008). As predicted, F2-S
males showed a significantly reduced anogenital distance and adult testis weights,
supportive of decreased testosterone exposure during the perinatal sensitive period.
Interestingly, studies examining prenatal stress during late pregnancy have also reported
decreased perinatal testosterone, adult anogenital distance, and testes weight in rats
(Dahlof et al., 1978; Ward and Weisz, 1980). It is important to note that we are
examining these measures in F2 animals that were not themselves exposed to any
prenatal manipulation. Thus, these data suggest that reduced exposure to organizational
30
experience, and that this effect can be transmitted along the paternal lineage to F2 male
offspring. Of translational importance, male schizophrenics have been reported to
display reduced circulating testosterone and disruptions in brain masculinization (Gur et
al., 2004; Goldstein et al., 2007). Further, boys with prepsychotic prodromal symptoms
had significantly lower testosterone levels during adolescents, a period of increased
psychotic disorder onset (van Rijn et al., 2011).
To examine F2-S male adult dysmasculinized and stress-sensitive phenotypes,
we measured their stress responsivity in the tail suspension test, Barnes maze, and HPA
stress axis. These tests were selected as they measure predictable sex differences in
stress-provoking environments, and performance in these tests was previously found to
be significantly dysmasculinized in F1 prenatally stressed males (Mueller and Bale,
2007, 2008). In the tail suspension test, F2-S males spent significantly more time
immobile than F2-C males. No effect of paternal prenatal stress was detected in
females. These results are similar to those reported in the first generation (Mueller and
Bale, 2008). While there was not a statistically significant effect of F2-S on overall
performance in the Barnes maze or HPA axis sensitivity, males did show a general trend
for a pattern of stress responsivity similar to that identified in F1 prenatally stressed
males (Mueller and Bale, 2008). Thus, it appears that aspects of the adult
dysmasculinized stress-sensitive phenotype were transmitted from F1 prenatally
stressed sires to their F2 male offspring. It is also possible that increased numbers of
litters may have provided sufficient statistical power to identify significant effects in
31
As our data point to a likely reduction in testosterone-mediated developmental
organization in F2-S male brains, we hypothesized that miRNAs in the brain are
responsive to organizational gonadal hormones. Therefore, in a subsequent study
examining the influences of estradiol on the neonatal brain miRNA environment, we
administered the aromatase inhibitor, formestane, to PN1 male neonates. miRNA
expression was then assayed using a miRNA Taqman qRT-PCR Array. Aromatase
inhibition dramatically dysmasculinized the brain miRNA environment where statistical
hierarchical clustering was unable to distinguish between formestane-treated males and
control females based on miRNA expression patterns, while completely segregating
control males from these groups. Thus, these data confirm the dynamic response of the
miRNA environment during this critical window. Gonadal hormones have previously
been shown to regulate miRNAs in peripheral target tissues (Klinge, 2009; Delic et al.,
2010; Narayanan et al., 2010). However, our data appear to demonstrate a novel impact
of organizational hormones on brain miRNA expression during the perinatal sensitive
period. Epigenetic mechanisms have been attributed to gonadal hormone status and
shown to influence brain sexual differentiation and may intersect with miRNAs to
program the sexually dimorphic brain (McCarthy et al., 2009b; Auger and Auger, 2011;
Auger et al., 2011).
Our studies provide intriguing evidence for the paternal transmission of prenatal
stress effects on neurodevelopmental processes including programming of the miRNA
environment and adult stress responsivity. Transmission through the paternal lineage
excludes confounds associated with maternal transmission, such as the intrauterine
32
(Youngson and Whitelaw, 2008). However, we cannot completely discount paternal
experience effects that, while unlikely, could occur during the brief time the males are in
the cage with females where stress-sensitive F1 males may impart some aspect of their
behavior upon the pregnant dam. A recent report examining a postnatal stress model
has also demonstrated the ability of early-life maternal separation to alter adult behavior
and methylation patterns of several genes in the germ line of male mice, with effects
persisting into second-generation offspring (Franklin et al., 2010). In addition, the
dysmasculinization we observed in F2-S male offspring importantly points to a
developmental window of susceptibility during which the programming effects of early
prenatal stress exposure may manifest. As such, identifying developmental processes
affected during this window, such as the dynamic changes in miRNAs detected, may
lead to critical therapeutic targets or biomarkers predictive for neurodevelopmental
diseases, particularly in at-risk pregnancies. Overall, these data support an early
gestational period vulnerable to prenatal stress epigenetic programming of the male
germline, permitting paternal transmission into subsequent generations.
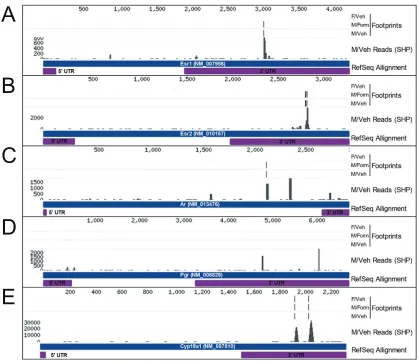
33
A
34 miR-322 miR-574-3p miR-873 0.0 0.5 1.0 1.5 * * * Re la ti ve e xp re ss io n Aromatase
ERα ERβ
35
Figure 2-1. Second-generation males from the paternal stress lineage (F2-S) show
dysmasculinized brain gene expression and miRNA expression patterns on postnatal
day 1 (PN1). A, Heatmap illustration of custom Taqman qRT-PCR Array results
demonstrating a broad shift in gene expression in the PN1 brain of F2-S male mice from
a male-typical (F2-C♂) to a more female-typical (F2-C♀) pattern. B, Statistical analyses
for sex differences detected 17 genes in the PN1 brain from our custom Taqman Array.
In S male PN1 brains, 13 of these 17 genes displayed expression levels closer to
F2-C females than to F2-F2-C male levels. C, As F2-S males show a reduced organizational
masculinization, we examined gene expression for central estrogen programming
targets: aromatase, estrogen receptor alpha (ERa) and beta (ERb). ERb was
significantly increased in the F2-S male PN1 brain compared to F2-C male. D,
Examination of the miRNA environment in F2 PN1 brain was examined using a miRNA
Array. miR-322, miR-574-3p, and miR-873 expression were dysmasculinized in F2-S
male mice. A single predicted shared gene target of these 3 miRs, β-glycan (TGFbr3),
was identified by the database miRDB.org and examined in F2 PN1 brain. Where we
found a reduction in miR expression in F2-S male mice, we detected an expected
increase in expression of β-glycan. All data are mean per group ± SEM, n = 3 - 5