Copyright 0 1991 by the Genetics Society of America
Chain Bias in Chi-Stimulated Heteroduplex Patches in the
X
ren
Gene Is
Determined
by
the Orientation of
X
cos
Ann T. Hagemann* and Susan
M.
Rosenbergt*'
*Institute of Molecular Biology, University of Oregon, Eugene, Oregon 97403, tDepartment of Biochemistry, University of Utah Medical Center, Salt Lake City, Utah 84132, and Department of Genetics, university of Alberta, Edmonton, Alberta T6G 2E9
Canada
Manuscript received February 13, 199 1
Accepted for publication July 6, 1991
ABSTRACT
Heteroduplex patch recombinants have received information in one DNA chain but have not recombined flanking markers. Evidence regarding which chain is exchanged bears on the structure
of recombination intermediates. The direction of travel along DNA of RecBCD recombinase, the central enzyme in the Escherichia coli RecBCD pathway of homologous recombination, is determined in phage X by the orientation of the packaging origin, cos. cos is a double-chain cut site which serves
as a preferred entry site for RecBCD. Using partially denaturing gels to resolve heteroduplex molecules, we have examined patch recombinants at the X Ten gene. We report that the transferred information in Chi-stimulated patches at ren can occur on either chain, but is biased to the chain ending 5' at the right of the X map (the X r chain) in phage carrying cos in its normal orientation. The chain bias switches in favor of the chain that ends 3' at the right (the X 1 chain) when RecBCD travel
direction is reversed by inverting cos. We entertain models that accommodate these and other results
pertaining to the structure of RecBCD-mediated recombinants.
R
EcBCD (ExoV) is the central enzyme in the Esch-erichia coli wild-type pathway of homologous recombination. Genetic and biochemical evidence in-
dicates that, when operating on phage X, RecBCD
loads onto the X chromosome at cos, the double-chain cut site that determines the ends of the packaged
chromosome [see THALER and STAHL (1988) for re-
view]. RecBCD can enter only at the right end of h because the packaging enzyme terminase remains bound to the left end after it cuts at cos (FEISS et al.
1983; KOBAYASHI et al. 1982; KOBAYASHI, STAHL and
CRASEMANN 1984). Once in the chromosome,
RecBCD travels unidirectionally, catalyzing a rate- limiting step or steps in recombination (STAHL et al.
1986; YAGIL and SHTROMAS 1985). If a Chi sequence,
5'GCTGGTGG (SMITH et al. 1981a), is present in
proper orientation with respect to the direction of RecBCD travel, stimulation of recombination occurs
near to and downstream from Chi in the enzyme's
travel path [see SMITH and STAHL (1985) for review).
Chi stimulates recombination in both cross partners
even if present in only one (STAHL, LIEB and STAHL
1984).
Heteroduplex patch recombinants contain a region of heteroduplex with flanking markers in a parental
configuration and are thought (in many models since
HOLLIDAY 1964) to arise via an alternative mode of
' Present address: Molecular Genetics, Cross Cancer Institute, 11560
berta T6G 122, Canada.
University Avenue, University of Alberta Medical School, Edmonton, Al- Genetics 129: 61 1-621 (November, 1991)
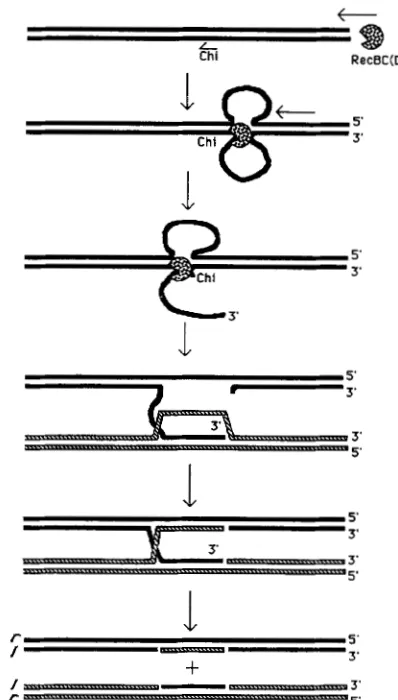
resolution from an intermediate that can also give rise to splice recombinants (those with flanking markers crossed over). T h e structure of a patch ( i . e . , which chain of DNA is exchanged) reveals aspects of the structure of the intermediate that gave rise to it in a way that splice junction structure cannot. This con- cept is illustrated in Figure 1 : Holliday junctions have been formed in which only one of the two chains is
swapped. Resolution of these structures to yield
patches results in patches in the swapped chain, re- gardless of the direction of propagation of the heter- oduplex (products
2
and 4). Resolution of these struc- tures to yield splices, however, produces different heteroduplex structures depending on whether the junction falls to the left or right of the heteroduplexregion (products 1 and 3). It is for this reason that we have chosen to analyze the structure of patches pro- duced by the RecBCD pathway.
We have examined heteroduplex structure in patch
recombinants with a method that utilizes differences
in melting properties of heteroduplex molecules to
resolve them on partially denaturing gels (method of
MYERS, MANIATIS and LERMAN 1987; used to study
yeast recombinants by LICHTEN et al. 1990). Under
612 A. Hagemann and S. M. Rosenberg
7
-
A "branch
I
I
-t
-
,- ( 1 )3
.
(3)"
3.
(2)-
(4)I
o r o r
3
.
FIGURE 1.-Features of the structure of recombination inter- mediates can be inferred from the structure of heteroduplex in patches. The intermediates here are formed by swapping only one chain, the chain ending 3' at the right. The products on the left of the figure result from alternative resolutions of an intermediate that has its Holliday junction on the left side of the heteroduplex region. The products on the right result from resolution of a Holliday junction that lies to the right of the heteroduplex. The patch products (2 and 4) both have heteroduplex that is white in the bottom chain, indicating that the bottom chain (3' at the right) was the one swapped. The splice products (1 and 3) do not reveal which chain was swapped in the intermediate because they have heteroduplex that is either black (splice junction with 3' overhangs) or white (splice junction with 5' overhangs) in the bottom chain, depending on which side of the heteroduplex the Holliday junction was on.
5' at the right end of the X map (designated the X r chain) when cos is in its wild-type (normal) orientation. T h e chain bias switches in favor of the opposite (X 1)
chain when the X cos site is inverted.
A nick initiation model for Chi activity (SMITH et
al. 1981b) (Figure 2) predicts patches biased exclu- sively to the X I chain when cos is oriented normally and to the r chain with inverted cos. We report patches on both chains, with bias opposite to that predicted
by the nick initiation model. We discuss models com-
patible with our results.
MATERIALS AND METHODS
Strain construction: Bacterial and phage strains used are listed in Tables 1 and 2. X Genetic elements are described in Table 3. All phage used are X red gum mutants, causing recombination to occur via the host RecBCD pathway rather than the X Red pathway.
Our physical markers are 18-base oligonucleotide inser- tions containing either a XhoI or an XbaI restriction site (see Figure 3 for sequences). These were ligated into SsdI-
linearized pKC31-4" and were then recombined into the various phage by genetic crosses, or were subcloned into pEMBL8'. pKC3 1-4" (gift of D. S. THALER) is a derivative of pKC31 (R. N. RAO, identical with plasmid pRLM4 de- scribed by WOLD et al. 1982) that contains a mutant, uncleav- able EcoRI site replacing the fourth X EcoRI site. (This aspect of its genotype was not important for our experi- ments.) Phage carrying the markers were identified by plaque-blot hybridization using 5' end-labeled oligonucleo- tides as probes. Orientations of markers were determined by producing single-chained versions of pEMBL8+-based plasmids (DENTE, CESARENI and CORTESE 1983) and probing them with the labeled oligonucleotides. All other X genetic elements were introduced by standard genetic crosses (AR- BER et al. 1983). Bacterial genetic elements were introduced by transduction with phage P1, and plasmids by transfor-
&
n
5'
3'
5'
-
3'3'
5'
1
5' 3'
v
-.1 3 3'
5'
J
+
1
-
3'r 5'
FIGURE 2.-A nick at Chi initiation model. In this model pro- posed by SMITH et al. (1981b) RecBC (now known as RecBCD) initiates recombination by unwinding DNA and nicking one chain at Chi. T h e nick at Chi is observed in vitro, and occurs near Chi on the chain ending 3' at the right ( I chain). Transfer of the nicked chain results in patches on the X 1 chain only when cos is oriented normally.
mation. Oligonucleotides used as markers and probes were made by the University of Oregon Biotechnology Labora- tory on an applied Biosystem model 380B DNA Synthesizer and also at Institut Jacques Monod, Universiti. Paris VII.
Growth of phage stocks: High titer X stocks were pre- pared by one of two methods. Those X capable of lysogeny (not carrying Ab1453) were grown by inducing lysogens of FS1576. All phage carry the Sum7 mutation, to allow ex- tended growth without lysis, and a temperature sensitive GI
allele, to allow heat induction of the prophage. Lysogen cultures were grown in supplemented LBK broth (1 % tryp- tone, 0.5% yeast extract, 1% NaCI, 75 mM CaCI2, 4 mM FeCI2, 2 mM MgS04, 10 pg/ml thymine and thiamine, pH 7.5) to an OD550 of 0.8, induced at 43" for 15 min, and incubated with vigorous shaking at 37" for 3 hr. Cells were concentrated by centrifugation and lysed by the method of SPRAGUE, FAULDS and SMITH (1978).
Phages carrying Ab1 453 were grown by infecting cultures of FS1576 using the protocol of SPRAGUE, FAULDS and SMITH (1978), except that aeration was by bubbling with 0 2 .
Chain Bias of RecBCD Pathway in X
TABLE 1
Bacterial strains and plasmids
613
Strain or
plasmid Relevant properties Reference or source
594 FS1576 FS1585 FS 1892 ATH58 ATH69 ATH70 S M R l l l pKC3 1
pAH9 pAH 15 pEMBL8' pAHlO
pAH18 pKC3 1-4"
Su- rec+ supE recDIOO9 supE supF recD1009 594 mutL216::TnlO
594 mutL216::TnlO daml3::Tn9 [pAH15] 594 rnutLPI6::TnlO [pAHlO]
594 mutL216::TnlO [pAH18]
594 mutL216::TnlO daml3::Tn9 [pAH9] Xdv plasmid with KanR marker
pKC3 1 with X's fourth EcoRI site mutated pKC31-4" with XbaI linker in SstII site pKC31-4" with XhoI linker in SstII site
pBR322-based (ColEldriven) plasmid with a phage f l ori pEMBL8+ with AuaI-BamHI fragment (containing XbaI
pEMBL8+ with AuaI-BamHI fragment (containing XhoI linker) subcloned from pAH9
linker) subcloned from DAH 15
WEIGLE ( 1 966) STAHL et al. (1 986) STAHL et al. (1986) D. S. THALER This paper This paper This paper This paper
R N. RAO (see WOLD et al. 1982) D. S. THALER
This paper This paper
DENTE, CESARENI and CORTESE ( 1 983) This paper
This paper
TABLE 2
Phage strains
Strain Genotype
XOH6 XOH 12 XOH29 XOH30 XOH35 XOH36 XOH39 XOH40 XOH46
XOH47
XOH48 XOH49
red3 gam210 imm2lcIts XbaI".' x+D Sam7 red3 gam210 imm2lcIts XhoI'O.' x+DSam7 cos21 Nulam5 red3 gam210 cI857 XhoI'O.' Sam7 cos21 NuIam5 red3 gam210 c1857 XbaI".' Sam7
cos2 COSML b1453 x+Z cI857 XholI'O.' Sam7
cos2 COSML b1453 x+Z cI857 Xbal".' Sam7 cos2 COSMR b1453 13857 XhoI'O.' X'D Sam7
cos2 cosMR b1453 cI857 XbaI".' x+D Sam7 cos2 cosML b1453 cI857 Xh0I'~.'Sam7 cos2 cosML b1453 cI857 XbaI".' Sam7 cos2 cosMR 61453 12857 XhoI'O.' Sam7
cos2 COSMR b1453 cI857 XbaI".' Sam7 All phage strains were generated for this work.
multiplicity of infection (moi) of 5 of each phage parent. For phage by plasmid crosses, the host strain carrying the plas- mid was inoculated into the cross medium from an overnight culture grown in the presence of the antibiotic selecting for the plasmid (kanamycin 50 pg/ml or ampicillin, 100 pg/ml). T h e host strains were SMRl 1 1 for plasmid pAH9, ATH 58 for pAH15, ATH69 for pAH10, and ATH70 for pAHl8. Phage were introduced at a moi of 7. Cross progenies were harvested as described for growth of phage stocks, and DNA was purified from the phage. In crosses with pAHlO or pAH 15, progenies were banded in CsCl equilibrium density gradients to eliminate any phage that had incorporated the plasmid by splicing. For analysis of heteroduplex in plasmid DNA, crosses were identical but supercoiled DNA was pu- rified by CsCI-EtBr equilibrium density gradient centrifu- gation after digestion of the X DNA in vitro with SstII.
Analysis of recombinants: For analysis of heterodu- plexes, DNA from crosses was cut with AluI, yielding a 279 basepair fragment that carries the marker. This fragment was chosen for analysis based on its melting behavior as predicted by the MELT program (LERMAN and SILVERSTEIN
1987), kindly provided by LEONARD LERMAN.
Heteroduplexes of the DNA carrying the marker were prepared in vitro by melting and annealing a mixture of the two AluI-digested parental types by the protocol of LECHNER and RICHARDSON (1983), or by boiling and slow cooling. The optimal denaturant concentration for resolution of these heteroduplex species was determined by running them on perpendicular denaturing gradient gels (MYERS, MAN-
IATIS and LERMAN 1987). The gel conditions used for all
further analyses were 1.05 M urea, 6% formamide (v/v) and 12% acrylamide (w/v), with electrophoresis for 6.5 hr at 150 V and 60". The DNA was transferred by semi-dry blotting (using a Hoeffer TE70 SemiPhor) to Zetabind nylon membrane (CUNO, Inc.) and was probed with a "P 5' end- labeled, 19 base oligonucleotide (5' GCGCCGGACGT- TGCCGCGC; DANIELS et al. 1983). This sequence is present in the restriction fragment containing the markers, outside of the marker sequences. Each heteroduplex species was identified using single-chained oligonucleotide probes to the marker sequences.
Autoradiographs were analyzed using a Biomed Instru- ments Zeineh SL-504-XL scanning densitometer and Video- phoresis I1 program. Band intensities were quantitated with the INT program for integration written by DAVID C. HAGEN (University of Oregon, Eugene).
Frequencies of recombination were determined by plat- ing progeny of phage crosses on FS 1585 and hybridizing plaque-blots with a probe specific to the marker sequence in the nonpackageable partner. XOH46, XOH47, XOH48 and XOH49 are non-Chi-containing versions of XOH35, XOH36, XOH39 and XOH40, respectively, and were used to determine Chi activity. For X by X crosses, plaque blots were also probed with a labeled oligonucleotide specific to the cos21 sequence in order to determine the maximum frequencies of packaging of cos21 N u l a m 5 and/or of unad- sorbed phage of the cos21 parental type.
RESULTS
614 A. Hagenlann and S. M. Rosenberg
TABLE 3
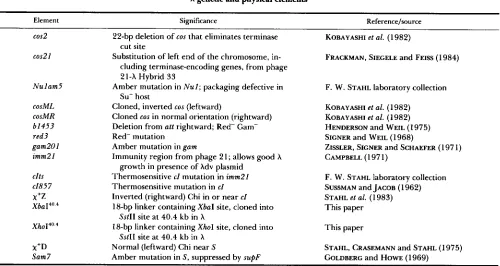
X genetic and physical elements
Element Significance Referencejsource
cos2 22-bp deletion of cos that eliminates terminase KOBAYASHI et al. (1982)
cos21 Substitution of left end of the chromosome, in- FRACKMAN, SIEGELE and FEISS (1 984) cut site
cluding terminase-encoding genes, from phage 21-X Hybrid 33
Su- host
Nu 1 am5 Amber mutation in N u l ; packaging defective in F. W . STAHL laboratory collection
cosML Cloned, inverted cos (leftward) KOBAYASHI et al. (1 982) cosMR Cloned cos in normal orientation (rightward)
b1453
KOBAYASHI et al. (1982)
red3 Red- mutation SIGNER and WEIL (1968)
gam201 Amber mutation in gam ZISSLER, SIGNER and SCHAEFER (197 1) imm21 Immunity region from phage 2 1 ; allows good X CAMPBELL (1 97 1)
clts Thermosensitive c l mutation in imm21 cI857
Xba14'
Deletion from att rightward; Red- Gam- HENDERSON and WEIL (1975)
growth in presence of Xdv plasmid
F. W. STAHL laboratory collection
STAHL et al. (1 983) Thermosensitive mutation in c l SUSSMAN and JACOB (1 962) 18-bp linker containing XbaI site, cloned into This paper
X+Z Inverted (rightward) Chi in or near c l
X ~ O I ~ O . ~ 18-bp linker containing XhoI site, cloned into This paper
x+D Normal (leftward) Chi near S STAHL, CRASEMANN and STAHL (1 975)
SstIl site at 40.4 kb in X
SstII site at 40.4 kb in X
Sam7 Amber mutation in S, suppressed by supF GOLDBERG and HOWE (1969)
paired bases are destabilized in partially denaturing gels, causing their mobility to be retarded relative to that of otherwise identical molecules without mis- matches. Because the extent of retardation depends on the particular mismatches present, some comple-
mentary heteroduplexes (e.g., molecules containing
A-G us. those containing C-T mispairs) can be resolved
from each other as well as from homoduplexes
(MYERS, MANIATIS and LERMAN 1987).
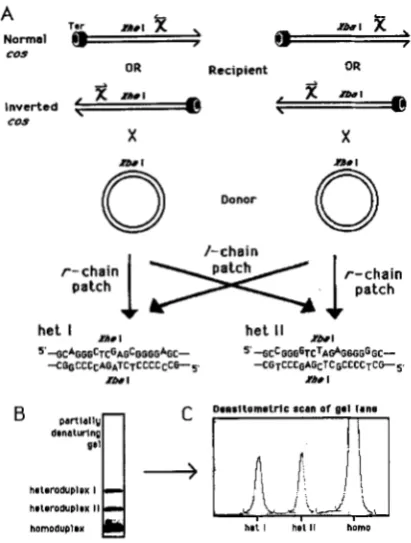
Partially denaturing gels were used to resolve com- plementary heteroduplex patches covering a site in the X ren gene. The markers introduced into ren are 18-base pair (bp) synthetic oligonucleotides. Each par- ent in a cross carries one of the two markers, which differ from each other by five single base changes such that heteroduplex formed across the markers contains five single-base mismatches. The marker se- quences, heteroduplex types produced, and designs
of the crosses are diagrammed in Figure 3. The ren
gene is involved in protecting X from the X Rex-
dependent exclusion system (TOOTHMAN and HER-
SKOWITZ 1980) and has no known phenotype in X infecting nonlysogenic bacteria.
Crosses were conducted in mutL host bacteria to minimize mismatch repair of heteroduplexes. GpmutL is required for a major and a minor mismatch repair
pathway in E . coli [see RADMAN (1 988) and MESELSON
(1 988) for reviews]. T o detect possible marker effects, including any residual mismatch repair, all crosses were done with markers in both possible orientations (see Figure 3). If the marker orientation with the XbaI
site in the donor gives heteroduplex I (het I) patches as the predominant type, then the marker orientation with XhoI in the donor will give het I1 as the majority species, barring marker effects. If one of the hets were preferentially mismatch repaired, this would be re- vealed as an apparent difference in the chain bias of
patches upon comparison of results with the two
marker orientations.
Crosses were either phage by phage, or phage by plasmid. In both procedures only one partner is re- covered, and it is in this partner (defined here as the recipient) that we detect patches. In the phage by
plasmid crosses, only patch recombinants are re-
covered; a phage chromosome that has incorporated the Xdv plasmid is too large to be packaged. The
progeny of crosses of X by the plasmids pAH 10 and
pAH 18 were centrifuged in CsCl equilibrium density gradients to remove the (more dense) phage particles containing a plasmid spliced into the X genome.
In the phage by phage crosses, one phage (the
donor) carries cos and a defective terminase (Nul
amber) derived from phage 21. Since phage 21 Nul
is not complemented by X, this phage chromosome is
not linearized and cannot be packaged in the nonsup- pressing cross host. Plaque-blot hybridization of X by X progeny sequentially with probes specific to the phage 21 sequence and the donor marker sequence
revealed that 0.9% and 1.3% (averages for two
Chain Bias of Rec :BCD Pathway in X 615
TABLE 4
Chi stimulation of patches at *cn in X by X crosses
/-chain
FIGURE 3.-A scheme for analyzing heteroduplexes produced with physical markers. A, X chromosomes are represented as du- plexes with one end blocked by Terminase (hex nuts), the other end providing a loading site for RecBCD. Chi ( x ) is present in its active orientation for each cos. For the marker orientation on the left side of the figure ( X b d in donor), het I forms as an r chain patch, whereas for the reverse orientation (XhoI in donor), het I
forms as an 1 chain patch (and vice versa for het 11). B, When run on partially denaturing gels, hets I and I1 are resolved from each other and from homoduplexes (het species were identified using chain-specific oligonucleotide probes to the markers). C, Intensities of het bands are quantified by densitometry.
could be either unadsorbed donor phage, donor phage that leaked through the nonsuppressing cross host, or donor phage that received the cloned cos (and thus the ability to package) by recombination (in an event distant from Chi and therefore not stimulated
by Chi). Of the progeny from crosses with and without
Chi, 2.0% and 1.0%, respectively, carry the cos21 sequence but do not hybridize with the donor marker
sequence at ren. These probably represent double
crossover products. In the DISCUSSION, we will con- sider the possibility that double crossovers contribute to our analysis of heteroduplex structure.
The two types of plasmids used here differ with respect to DNA replication. Xdv plasmids (pAH9,
pAHl5) replicate using the X origin and replication
proteins (MATSUBARA 198 1). pAH 10 and pAH 18 use
a ColEl origin. The extent of homology available for
recombination with X is also different for the two types of plasmids; both have approximately 1.3 kilo- basepairs (kb) of homology to the right of the marker, but to the left, the Xdv plasmids have about 1.8 kb, whereas the ColEI-driven plasmids have about 0.5 kb. It was not known whether the differences in replica-
Recombi-
Pha e fre nation uency Chi lation stimu- Cross Chi+'O yie&
ibm?
(Chi+/Chi")XOH40 X XOH29 Chi+ 49.0 XOH49 X XOH29 Chi" 13.7 hOH39 X XOH30 Chi+ 46.8 XOH48 X hOH30 Chi" 10.0 XOH36 X AOH29 Chi+ 12.2 XOH47 X hOH29 Chi" 6.7 hOH35 X XOH30 Chi+ 21.6 XOH46 X XOH3O Chi" 7.6
5.7 5.7
1
.o
5.3 7.60.7 5.0 5.0
1
.o
5.9 5.91
.o
Phage yields are the number of phage harvested divided by the number of cells in culture at time of infection, averaged for two
crosses.
'
Recombination frequencies are determined by plaque-blot hy- bridization and are the percent of recipient phage that carry the donor marker sequence. These are averages for two to three crosses.tion and extent of homology would affect patch-chain
bias, however, crosses conducted with these plasmids yielded similar results (see Table 5).
Chi stimulates patches at the ren locus: Recombi- nation frequencies for X by X crosses with and without Chi were measured by plaque-blot hybridization with
the donor marker oligonucleotides (Table 4). The
plaques that hybridized with both the donor marker
probe and the cos21 probe (as mentioned above) were
not counted as recombinants. With Chi present, the frequency of patches is at least fivefold greater than in the corresponding crosses without Chi. Since Chi stimulates exclusively RecBCD pathway recombina-
tion (STAHL and STAHL 1977), the patches produced
in the Chi-stimulated crosses analyzed here are ex-
pected to be RecBCD-mediated patches.
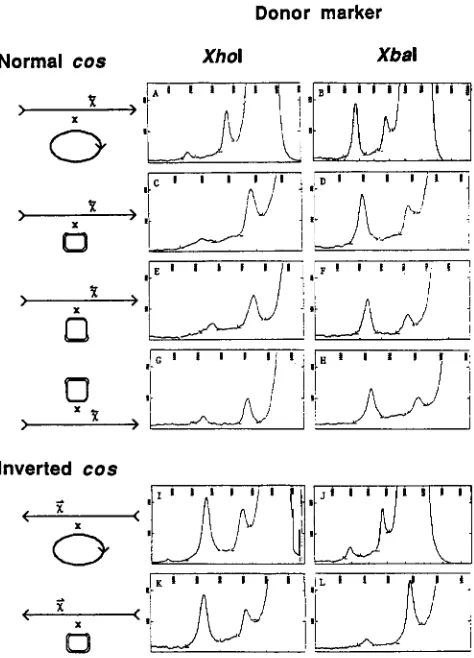
Structure of Chi-stimulated patches at ren with cos
oriented normally: Patches in both the X r chain and the X 1 chain are found, with r-chain patches being the
predominant type recovered from crosses with cos in
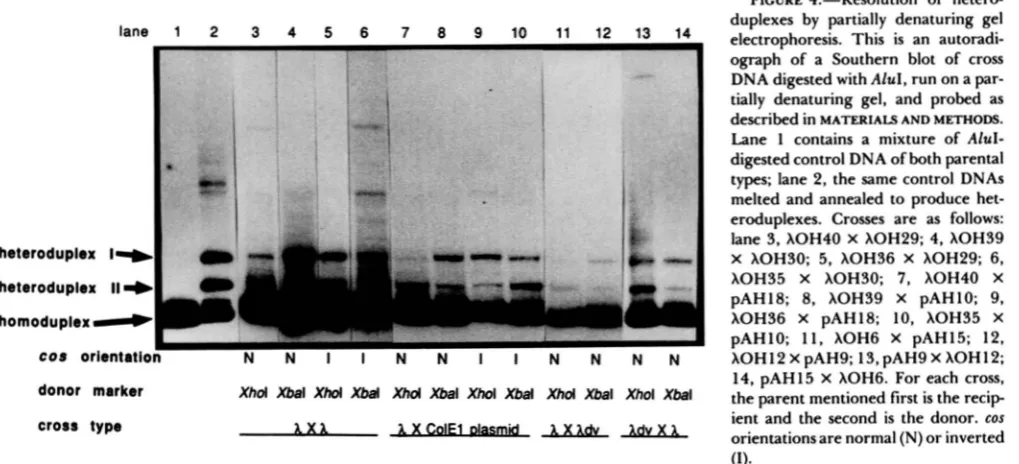
the wild-type (normal) orientation. This is apparent as an excess of het I1 in normal cos experiments with the
XhoI donor marker (Figure 4 lanes 3,
7
and 11).Scanning densitometry of these lanes reveals consist- ently larger peaks for het I1 (Figure 5 scans A, C and E). Similarly, r-chain patches are the majority type produced in crosses with the donor bearing the XbaI marker, as seen by an excess of het I (Figure 4 lanes 4 , 8 and 12; Figure 5 scans B, D and F). Table 5 shows that the r-chain bias is twofold or greater for Chi-
stimulated patches with cos oriented normally.
Dependence of patch chain bias on cos orientation:
Crosses with phage carrying inverted cos and Chi
616 A. Hagemann and S.
M.
Rosenberglane 1 2 3 4 5 6 7 8 9 10 11 12 13 14
heteroduplex 1 3 )
heteroduplex II
+
homoduplex-
Cod orisntatlon N N I I N N I I N N N N
"
donor marker Xhd Xbal Xhd Xbal Xhd Xbel Xhd Xbal Xhd Xbel Xhol Xbel cross type
Ix1
dx Co'FIolasma
AxWLA u L
apparent as an excess of het I from crosses with the XhoI marker in the donor (Figure 4 lanes 5 and 9;
Figure 5 scans I and
K)
and as an excess of het I1 from crosses with the XbaI marker in the donor (Figure 4 lanes 6 and 10; Figure 5 scans J and L). The bias toward l-chain patches is twofold or greater for all of the cross types examined (Table 5). The similarity of the results obtained with each of the two marker orientations rules out marker effects as the source of the observed patch-chain bias.Do
patches onto both partners in a cross have the same structure? Supercoiled plasmid DNA was iso- lated from cells following infection with phage and was analyzed for heteroduplexes. In Xdv plasmids that were crossed by phage with normal cos, the chain patched most often was the same as for the phage, the r chain (Figure 4, lanes 13 and 14; Figure 5 , scans Gand H). The bias was approximately twofold (Table 5). Thus the chain swapped most often was the same for both the Chi-containing partner and the non-Chi- containing partner in X by Xdv plasmid crosses, and the same extent of bias was observed. It was not possible to recover quantities of heteroduplex
SUE-
cient for analysis from crosses with the ColEldriven plasmids.DISCUSSION
Summary of results: We report that Chi-stimulated heteroduplex patches in the X ren gene are present on either chain of DNA, but display at least a twofold chain bias that is determined by the orientation of cos.
With normal cos, Chi-stimulated patches are predom- inantly in the X r chain; with cos inverted, Chi-stimu- lated patches in the X 1 chain predominate. Because the orientation of cos determines the direction of
FIGURE 4.-Resolution of hetero- duplexes by partially denaturing gel electrophoresis. This is an autoradi- ograph of a Southern blot of cross DNA digested with A M , run on a par- tially denaturing gel, and probed as described in MATERIALS AND METHODS.
Lane 1 contains a mixture of AluI- digested control DNA of both parental types; lane 2, the same control DNAs melted and annealed to produce het- eroduplexes. Crosses are as follows: lane 3, XOH40 X XOH29; 4, XOH39
X XOH30; 5, XOH36 X hOH29; 6, XOH35 X XOH30; 7, XOH40 X
pAHl8; 8, XOH39 X pAH10; 9, XOH36 X pAHl8; 10, XOH35 X
pAHlO; 1 1 , XOH6 X pAH15; 12, XOHl2 X pAH9; 13, pAH9 X XOH12; 14, pAH15 X XOH6. For each cross, the parent mentioned first is the recip ient and the second is the donor. COS orientationsare normal (N) or inverted (1).
travel of RecBCD along DNA (STAHL et al. 1986), this patchthain bias likely reflects some difference in treatment of the two chains by RecBCD. For both orientations of cos, the chain to which patches at ren
are biased is the chain that ends 5' at the end of the DNA that RecBCD enters.
The observation of COS orientationdependent bias bears on models for RecBCD-mediated recombina- tion. A previous report of patchthain bias independ- ent of cos orientation (ROSENBERC 1988) argued strongly against models in which RecBCD acts as a chain-specific initiator or a chain-biased resolvase, be- cause such models predict either no bias or cos orien- tationdependent bias. These findings at Ten allow reconsideration of possible roles for RecBCD in re- combination.
Are patches the products of two close splices? T o
relate heteroduplex patch structures to current models for recombination, we will assume that a ge- netic patch arises in a single chain-swapping event
(e.g., see Figure 1). However, it is not possible for us to distinguish a single chain transfer from a pair of close double chain transfers, or splice events. Because the possibility of the latter complicates interpretation of our results, we will first review evidence supporting the assumption that heteroduplex patches arise from single chain transfers.
Chain Bias of RecBCD Pathway in X 617
Donor marker
I
c
I
i S
FIGURE 5.-Densitometric scans of complementary heterodu- plex bands resolved in partially denaturing gels. The cross type for each pair of crosses is represented diagrammatically at the left; the upper partner in each couple is the recipient, the one that is recovered. X Chromosomes with cleavable cos sites (discussed in text) are represented as linear, with arrowheads representing the orientation of cos. Plasmids and X chromosomes with uncleavable
cos sites are represented as small and large circles, respectively. The marker present in the donor is indicated above each column. Crosses in the XbaI column are isogenic to those in the XhoI column, but have the markers reversed. Cross genotypes follow: A, XOH40 X
XOH29; B, XOH39 X XOH30; C, XOH40 X pAH18; D, XOH39 X
pAH10; E, XOH6 X pAH15; F, XOH12 X pAH9; G , pAH9 X
XOH12; H, pAHl5 X XOH6; I, XOH36 X XOH29; J. XOH35 X
XOH30; K, XOH36 X pAH18; L, XOH35 X pAHlO.
Chi, the partner lacking Chi is not expected to acquire
patches through a double-splice mechanism. If the
Chi+ (with Chi) and Chi" (without Chi) patch recom- binants are formed by different mechanisms, different patch structures might be expected. We observe that both have the same structure.
Chi stimulates patches onto the Chi+ partner (taken patches) as well as onto the Chi" partner (given
patches) when only one carries Chi (STAHL, LIEB and
STAHL 1984). We report that in crosses of X by Xdv, both given and taken patches are r-chain biased when cos is oriented normally. [Given and taken patches stimulated by Chi and found at the X P gene were
both also r-chain biased (ROSENBERG 1987, 1988).] In
the context of evidence that the partner lacking Chi
enjoys a single recombination event (STAHL et al.
1990), our results imply that the heteroduplex patches we observe reflect the primary, single event and are not likely to result from double splices, since both partners in the cross show the same magnitude and chain of bias (see Figure 6). We will consider that the production of Chi+ recombinants, both splices and patches, does involve a pair of events, but that the second event may be spatially separated from and
need not impinge upon the first (see example in Figure
6B). In forming a packageable patch recombinant in the RecBCD pathway it is probably always necessary to engage in a splice event, as well as a patch event, in
order to satisfy the apparent dimer requirement for X
packaging [see FEISS and BECKER (1 983) for review].
It has been reported that in RecBCD-mediated
crosses lacking Chi, heteroduplex DNA at splice junc- tions (sites of recombination events exchanging flank- ing markers) in the X cZ and P genes has predominantly
3' overhangs (as in Figure 6A; SIDDIQI, STAHL and STAHL 199 1). If patches result from two consecutive splices as in Figure 6A, the structure of the splice
junctions (3' overhangs) would demand that the plas-
mid in a phage by plasmid cross would receive pri- marily l-chain patches, with normal cos. We find that patches in such plasmids are r-chain biased, with the same extent of bias as in the phage partner, strength- ening the argument against double splices as the main route to formation of patches.
Nick at Chi initiation model: In a nick at Chi initiation model for RecBCD-mediated recombination (SMITH et al. 198 1 b) chain bias in heteroduplex for- mation is determined by exchange of the X
I
chain, the chain that suffers the nick at Chi in vitro (see Figure2).
This model predicts an absolute chain bias that is dependent on the direction of RecBCD travel. In contrast with that model's predictions, we find patches on both chains, with bias opposite to that predicted. Several other aspects of the nick at Chi initiation model conflict with recent genetic and bio-chemical observations regarding RecBCD-mediated
recombination (reviewed by ROSENBERG and HAS-
What causes patch chain bias? Heteroduplex patch chain bias could result from bias in initiation or reso- lution steps of recombination (or both), and must reflect the polarity of RecBCD, although polarities of other recombination enzymes and DNA ends, as well as locus-specific factors (as discussed below for the X
P locus) may play roles in determining chain bias. The twofold bias we report for heteroduplex in the X ren gene could reflect different frequencies of transfer
for the two chains, or different lengths of transferred
segments on the two chains. A model in which
RecBCD initiates recombination by providing single- stranded DNA ends has been proposed (ROSENBERG
and HASTINGS 1991). This model achieves chain bias
618 A. Hagemann and S. M. Rosenberg
TABLE 5
Patchchain bias with different orientations of cos
Recipient
parent parent Donor
~~~~ ~~
cos orien- Donor r-chain patches Gchain patches tation marker l-chain patches r-chain patches
XOH40 XOH39 AOH36 XOH35 XOH40 XOH39 XOH36 XOH35 XOH6
XOH 12 pAH9 DAH^^
hOH29 XOH30 XOH29 XOH30 pAH18 pAH 10 pAHl8 pAHlO pAHl5 pAH9 XOH 12 XOH6
Normal Normal Inverted Inverted Normal Normal Inverted Inverted Normal Normal Normal Normal
XhoI XbaI XhoI XbaI XhoI XbaI XhoI XbaI XhoI XbaI XhoI XbaI
3.8 f 1.8 (5) 2.0 f 0.1 (2)
2.8 f 1.1 (4) 2.4 f 0.3 (7) 5.0
*
3.7 (3)3.4 f 1.5 (3)
3.0
*
0.5 (3) 6.0*
3.8 (2) 2.1 f 0.7 (20)2.2 f 0.3 (9) 2.7 f 1.6 (2) 2.6 f 0.1 (2)
Numerical values given are means with standard deviations. The number of gel lanes quantitated, representing a minimum of two crosses of each type. is indicated in parentheses. For data from t w o repetitions, f values are calculated as for standard deviations.
A B
. .
I
I
L
1
FIGURE 6.-Nonreciprocal models for generating patches. A, Chistimulated recombination of X with a plasmid (open lines) involves loss
of the right arm of the chromosome up to the crossover, necessitating a second round of recombination with another X to produce a packageable recombinant carrying Chi. By this model, patches into X necessarily arise as double splices. In the plasmid a double event is not needed and patch structure reflects the chemistry of the initial interaction. This model is incompatible with our demonstration that patches onto the Chicontaining (A) partner have the same structure as patches onto the (plasmid) partner lacking Chi (discussed in the text). B, In this model, degradation occurs up to Chi, leaving an incomplete X recombinant that must splice with another X in order to be packaged. Patches may be produced by swapping a single chain of DNA, and patch structure in both partners reflects the chemistry of the initial interaction. Model B is compatible with our results, with those of STAHL et al. (1990). and with the observed degradation of DNA in vitro
between the RecBCD loading site and Chi (DIXON and KOWALCZYKOWSKI 1991).
by
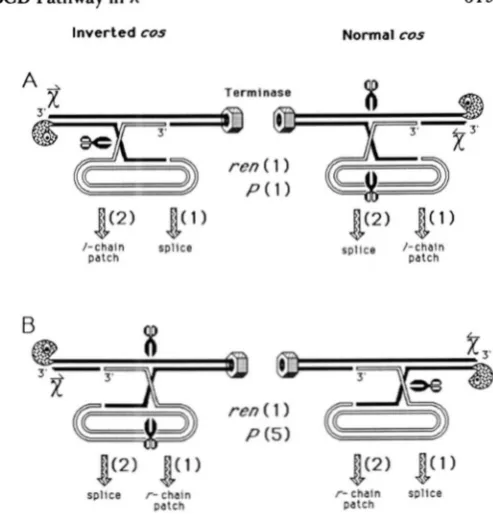
allowing formation of longer heteroduplex from 5’ ends than 3’ ends. We will entertain a model for RecBCD-mediated recombination in which RecBCD exerts its influence on chain bias in a resolution step.A
biased resolution model for formation of patches: In this model (Figure 7) Holliday junctions are formed independently of RecBCD, and result from 3’ end invasion only (possibly due to a 3’ end requirement for strand transfer by RecA; KONFORTI and DAVIS 1987). RecBCD degrades the engagedchromosome up to Chi, and is transformed from an exonuclease into a Holliday junction resolvase upon recognition of Chi. This transformation could be caused by loss of the RecD subunit. (THALER et al.
Chain Bias of RecBCD Pathway in X 619
Inverted cos Normal cos
Normal cos
m Inverted cos
A
I
I-chain patch SPllCR
D
u m
b
*
3 -d 3'B
aD
b
SDIICB r-chuln putch
FIGURE 7.-Biased resolution model. Initiation of recombination is assumed to be independent of RecBCD and occurs on either chain by RecA-catalyzed 3' end invasion. RecBC (having lost RecD and its exonuclease activity as a consequence of passing Chi) is proposed to resolve Holliday junctions by cutting the chain that ends 5' where it enters, producing 3' overhangs at splice junctions, r-chain patches (those on the chain ending 5' at the right) with normal cos, and I-chain patches (on the chain ending 3' at the right) with inverted cos. Notched circles represent RecBC; nuts represent terminase; scissors indicate the chains cut in Holliday junction resolution.
FIGURE 8.-Biased resolution model amended to explain patch structure at both the ren and P loci. The model in Figure 7 is altered as follows: RecBC is proposed to resolve Holliday junctions by cutting the chain that ends 5' at its loading site twice as often as it cuts the chain ending 3' at the loading site. This gives the twofold bias observed at ten. Now suppose that the single-chain breaks that must occur in order to initiate (or complete) the Holliday junction intermediate have a nonuniform distribution. This would allow the formation of the two types of junctions (A, top; B, bottom) at different relative frequencies in different parts of the chromosome (the 3' end invasion rule still applies here). An abundance of nicks near any locus will cause an abundance of either the A type or the B type structure at the locus in question depending on whether the concentration of nicks (or intermediates that are nicked to form Holliday junctions) falls to the left or right of that locus. At P, if the replication origin (600 bp left of the P marker) provides such a concentration of initiation sites, a majority of heteroduplexes at P
will arise from intermediate B. If B is five times as abundant as A, an r-chain bias in patches of 2.5-fold for inverted cos and 10-fold for normal cos will result. The r-chain bias observed at P was at least 9-fold for normal cos, and at least 3-fold for inverted cos (ROSENBERG 1987. 1988). Symbols are the same as in Figure 7. chain with normal cos and to the 1 chain with inverted
cos. This model is compatible with results presented here for patches at the ren locus and with the nonre- ciprocal model of Figure 6B. It also gives the 3' overhangs reported for splice junctions formed in the RecBCD pathway (SIDDIQI, STAHL and STAHL 199 1).
Why is patch-chain bias cos orientationdependent at rets but orientation-independent at
P?
A previous genetic analysis of patches at a marker in the XP
gene, 630 bp left of the marker used here in Ten, revealed patches biased to the X r chain, regardless of the orientation of cos. The bias reported at P was stronger; within the limits of detection, chain bias favored ther chain by at least ninefold with normal cos (ROSEN- BERG 1987) and at least threefold with inverted cos
(ROSENBERG 1988). A possible basis for the stronger bias is discussed in Figure 8.
Three important differences between experiments at
P
and ren follow: (1) two different experimental methods were used;(2)
two different loci were ex- amined using two different markers; (3) with cos in- verted, the condition under which chain bias differed atP
and Ten, the crosses designed to detect patches atP were not Chi stimulated, whereas those detecting patches at Ten were Chi stimulated. The difference between results at the two loci could reflect the dif- ferent methods of analysis, and attempts to assay both loci with a single method are in progress. Because we
can identify no fault in either method, we will cur- rently consider other explanations for the different results.
The difference between results at ren and
P
might reflect the fact that in the P experiments recombina- tion was not stimulated by Chi when cos was inverted. Chi" experiments could not be examined with the physical method used at ren due to insufficient yield of recombinants, and inverted Chi+ experiments were620 A. Hagemann and S. M. Rosenberg
DIQI, STAHL and STAHL (1 99 1) for discussions of such evidence]. We think it likely that, rather than reflect- ing a difference in the mechanism of recombination
with and without Chi, the difference between the ren
and P results reflects a location-specific factor. Figure 8 illustrates how a locus-specific factor can cause recombination proceeding by the model in Fig- ure
7
to yield cos orientation-dependent bias at most loci, but predominantly r chain-biased patches at one locus ( P ) , regardless of the orientation of cos. In this model the rules for resolution are relaxed to allow for production of the minority type of heteroduplex ob- served at Ten, and initiation is postulated to occur with different frequency at different sites. By this model, frequency of initiation at a particular site will influ-ence which chain is exchanged, but is not expected to
affect the rate of recombination, because the rate-
limiting step in RedBCD-mediated recombination is
the step catalyzed by RecBCD and stimulated by Chi, here imagined to be resolution.
A greater understanding of the mechanism of re- combination in the RecBCD pathway will be gained
by determining whether patch-chain bias is a conse- quence of more frequent or longer heteroduplex on a given DNA chain, and whether this difference in treatment of the two chains is manifest during for-
mation or resolution of a recombination intermediate.
We wish to thank LEONARD LERMAN, TIM BULLOCK, DAN GRA- HAM, MICHAEL LICHTEN, DAVID HAGEN, MARY STAHL, and MIR-
JANA PETRANOVIC for technical advice and CATAYANA Hsu for assistance. We enjoyed and benefited from discussions with all our colleagues, especially FRANK STAHL, IMRAN SIDDIQI, RIK MYERS, MIRO RADMAN and GISELA MOSIG. We are grateful to R. MYERS, J.
SAWITZKE, F. STAHL, P. J. HASTINGS, L. CALAVETTA, L. THOMASON,
J. HABER, M. LICHTEN, M. RADMAN, G. R. SMITH, D. S. THALER,
S. BURGESS and 0. HUGHES for thoughtful criticisms of the manu- script. FRANK and MARY STAHL gave invaluable advice and encour- agement. This work was supported by National Institutes of Health (NlH) GM3367 and National Science Foundation PCM8905310 to FRANKLIN STAHL (University of Oregon) and NIH GM41747-01 to S.M.R. and D. CARROLL (University of Utah). A.T.H. was s u p
ported by NIH Genetics Training Grant GM07413-09 (University
of Oregon); S.M.R. was supported by NIH Postdoctoral Fellowship GM12406 (University of Utah and University of Alberta). S.M.R. is grateful to M. RADMAN and the Association pour le Recherches sur le Cancer for supporting her visit to his laboratory at Institut Jacques Monod, Universiti Paris VII.
LITERATURE CITED
ARBER, W., L. ENQUIST, B. HOHN, N. E. MURRAY and K. MURRAY, 1983 Experimental methods for use with lambda, pp. 433- 466 in Lambda I I , edited by R. W. HENDRIX, J. W. ROBERTS, F. W. STAHL and R. A. WEISBERG. Cold Spring H a r b o r Labo- ratory, Cold Spring Harbor, N.Y.
CAMPBELL, A., 197 1 Genetic structure, pp. 13-14 in The Bacteri- ophage Lambda, edited by A. D. HERSHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
CHENG, K. C., and G. R. SMITH, 1984 Recombinational hotspot activity of Chi-like sequences. J. Mol. Biol. 1 8 0 371-377. DANIELS, D., J. SCHROEDER, W. SZYBALSKI, F. SANGER, A. COUISON,
G. HONG, D. HILL, G. PETERsoN and F. BLATTNER, 1983 The
complete annotated lambda sequence, pp. 51 9-676 in Lambda
II, edited by R. W. HENDRIX, J. W. ROBERTS, F. W. STAHL and R. A. WEISBERG. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
DENTE, L., G. CESARENI and R. CORTESE, 1983 pEMBL: a new family of single stranded plasmids. Nucleic Acids Res. 11:
DIXON, D. A., and S. C. KOWALCZYKOWSKI, 1991 Homologous pairing in vitro stimulated by the recombination hotspot, Chi. Cell 6 6 1-20.
FEISS, M., and A. BECKER, 1983 DNA Packaging and cutting, pp. 305-350 in Lambda II, edited by R. W. HENDRIX, J. W. ROBERTS, F. W. STAHL and R. A. WEISBERG. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
FEISS, M., W. WIDNER, G. MILLER, G. JOHNSON and S. CHRISTIAN-
SEN, 1983 Structure of the bacteriophage X cohesive end site:
location of the sites of terminase binding ( c o d ) and nicking (cosN). Gene 24: 207-2 18.
FRACKMAN, S . , D. A. SIEGELE and M. FEISS, 1984 A functional domain of bacteriophage X terminase for prohead binding. J. Mol. Biol. 1 8 0 283-300.
GOLDBERG, A., and M. HOWE, 1969 New mutations in the S cistron of bacteriophage Lambda affecting host cell lysis. Vi- rology 38: 200-202.
HENDERSON, D., and J. WEIL, 1975 Recombination-deficient dele- tions in bacteriophage X and their interactions with Chi muta- tions. Genetics 7 9 143- 174.
HOLLIDAY, R., 1964 A mechanism for gene conversion in fungi. Genet. Res. 5 282-304.
KOBAYASHI, I., M. M. STAHL and F. W. STAHL, 1984 The mech- anism of Chi-cos interaction in RecA-RecBC-mediated recom- bination. Cold Spring Harbor Symp. Quant. Biol. 4 9 497- 506.
KOBAYASHI, I., H. MURIALDO, J. M. CRASEMANN, M. M. STAHL and F. W. STAHL, 1982 Orientation of cohesive end site cos de- termines active orientation of Chi sequences in stimulating recA. recBC-mediated recombination in phage X lytic infec- tions. Proc. Natl. Acad. Sci. USA 7 9 5981-5985.
KONFORTI, B. B., and R. W. DAVIS, 1987 3' Homologous free ends are required for stable joint molecule formation by the RecA and single-stranded binding proteins of Escherichia coli. Proc. Natl. Acad. Sci. USA 8 4 690-694.
LECHNER, R. L., and C. C. RICHARDGON, 1983 A preformed, topologically stable replication fork. J. Biol. Chem. 2 5 8
LERMAN, L. S., and K. SILVERSTEIN, 1987 Computational simu- lation of DNA melting and its application to denaturing gra- dient gel electrophoresis. Methods Enzymol. 1 5 5 482-501. LICHTEN, M., C. GOYON, N. P. SCHULTES, D. TRECO, J.
w.
SZOSTAK,J. E. HABER and A. NICOLAS, 1990 Detection of heteroduplex DNA molecules among the products of Saccharomyces cereuisiae meiosis. Proc. Natl. Acad Sci. USA 87: 7653-7657.
MATSUBARA, K., 1981 Replication control system in lambda dv. Plasmid 5: 32-52.
MFSELSON, M., 1988 Methyl-directed repair of DNA mismatches, pp. 91-1 13 in The Recombination of Genetic Material, edited by K. B. Low. Academic Press, New York.
MYERS, R. M., T. MANIATIS and L. S. LERMAN, 1987 Detection and localization of single base changes by denaturing gradient gel electrophoresis. Methods Enzymol. 1 5 5 501-527. RADMAN, M., 1988 Mismatch repair and genetic recombination,
pp. 169-192 in Genetic Recombination, edited by R. KUCHER-
LAPATI and G. R. SMITH. American Society for Microbiology,
Washington, D.C.
ROSENBERG, S. M., 1987 Chi-stimulated patches are heteroduplex, with recombinant information on the phage X r chain. Cell 4 8 855-865.
ROSENBERG, S. M., 1988 Chain-bias of Escherichia coli Rec-me- 1645-1655.
Chain Bias of RecBCD Pathway in X 62 1
diated X patch recombinants is independent of the orientation of X cos. Genetics 1 1 9 7-2 1.
ROSENBERG, S. M., and P. J. HASTINGS, 1991 The split-end model for homologous recombination at double-strand breaks and at Chi. Biochimie 73: 385-399.
SIDDIQI, I . , M. M. STAHL and F. W. STAHL, 1991 Heteroduplex chain-polarity in recombination of phage X by the Red, RecBCD, RecBC(D-) and RecF pathways. Genetics 128: 7-22. SIGNER, E. R., and J. WEIL, 1968 Recombination in bacteriophage
X. I. Mutants deficient in general recombination. J. Mol. Biol.
SMITH, G. R., and F. W. STAHL, 1985 Homologous recombination promoted by Chi sites and RecBC enzyme of Escherichia coli. Bioessays 2: 244-249.
SMITH, G. R., S. M. KUNES, D. W. SCHULTZ, A. TAYLOR and K. TRIMAN, 1981a Structure of Chi hotspots of generalized re- combination. Cell 24: 429-436.
SMITH, G. R., D. W. SCHULTZ, A. TAYLOR and K. TRIMAN, 1981b Chi sites, RecBC, and generalized recombination. Stadler Genet. Symp. 13: 25-37.
SPRAGUE, K. U., D. H. FAULDS and G. R. SMITH, 1978 A single base-pair change creates a Chi recombinational hotspot in bacteriophage X. Proc. Natl. Acad Sci. USA 75: 6182- 6186. STAHL, F. W., J. M. CRASEMANN and M. M. STAHL, 1975 Rec-
mediated recombinational hotspot activity in bacteriophage lambda. 111. Chi mutations are site mutations stimulating Rec- mediated recombination. J. Mol. Biol. 94: 203-212.
STAHL, F. W., M. LIEB and M. M. STAHL, 1984 Does Chi give or take? Genetics 1 0 8 795-808.
STAHL, F. W., and M. M. STAHL, 1977 Recombination pathway specificity of Chi. Genetics 8 6 715-725.
STAHL, M. M., I. KOBAYASHI, F. W. STAHL and S. K. HUNTINGTON, 1983 Activation of Chi, a recombinator, by the action of an endonuclease at a distant site. Proc. Natl. Acad. Sci. USA 8 0 3 4 256-271.
2310-2313.
STAHL, F. W., I. KOBAYASHI, D. S. THALER and M. M. STAHL, 1986 Direction of travel of RecBC recombinase through bac- teriophage lambda DNA. Genetics 113: 2 15-227.
STAHL, F. W., L. C. THOMASON, I. SIDDIQI and M. M. STAHL, 1990 Further tests of a recombination model in which x removes the RecD subunit from the RecBCD enzyme of Esch-
erichia coli. Genetics 1 2 6 519-533.
SUSSMAN, R., and F. JACOB, 1962 Sur un systi.me de repression thermosensible chez le bactkriophage X d’ Escherichia coli. C. R. Acad. Sci. (Paris) 2 5 4 1517-1519.
THALER, D. S., and F. W. STAHL, 1988 DNA double-chain breaks in recombination of phage X and of yeast. Annu. Rev. Genet.
THALER, D. S . , E. SAMPSON, I. SIDDIQI, S. M. ROSENBERG, L. C.
THOMASON, F. W. STAHL and M. M. STAHL, 1989 Recombination of bacteriophage X in recD mutants of Escherichia coli. Genome 31: 53-67.
TOOTHMAN, P., and I. HERSKOWITZ, 1980 Rex-dependent exclu- sion of lambdoid phages. 11. Determinants of sensitivity to exclusion. Virology 102: 147-160.
WEIGLE, J., 1966 Assembly of phage lambda in vitro. Proc. Natl. Acad. Sci. USA 55: 1462-1466.
WOLD, M. S., J. B. MALLORY, J. D. ROBERTS, L. H. LE BOWITZ and R. MC MACKEN, 1982 Initiation of bacteriophage X replica- tion in vitro with purified X replication proteins. Proc Natl. Acad Sci USA 79: 6176-6180.
YAGIL, E., and I . SHTROMAS, 1985 Rec-mediated recombinational activity of two adjacent Chi elements in bacteriophage lambda. Genet. Res. 4 5 1-8.
ZISSLER, J., E. SIGNER and F. SCHAEFER, 1971 The role of recom- bination in the growth of bacteriophage lambda. I . The gamma gene, pp. 455-468 in The Bacteriophage Lambda, edited by A. D. HERSHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
22: 169-197.