ABSTRACT
CRUMLEY, JENNIFER LEE. Effects of Genotype and Environment on Agronomic
Performance and Phytochemical Profile of Echinacea purpurea and Echinacea angustifolia. (Under the direction of Jeanine M. Davis.)
The genus Echinacea, in the family Asteraceae, is composed of nine species of
herbaceous, perennial flowering plants commonly referred to as coneflowers. Native to North America, a variety of species can be found growing in moist open woods in parts of the United States and in southern regions of Canada. Echinacea has an extensive history of use by Native Americans for its medicinal properties. In modern times, Echinacea is one of the most popular medicinal herbs in today’s market, with domestic sales reaching $28 million in 2013. Echinacea species are still commonly wild-harvested for use in the herbal market; however, in recent years improvements in cultivation practices have reduced dependency on wild plants.
Field studies were conducted to determine the effect of growing environment, plant age, and seed source on growth, yield, and production of several important phytochemicals in
Echinacea purpurea and E. angustifolia. Three year field trials were conducted in the
Effects of Genotype and Environment on Agronomic Performance and Phytochemical Profile of Echinacea purpurea and Echinacea
angustifolia.
by
Jennifer Lee Crumley
A thesis submitted to the Graduate Faculty of North Carolina State University
in partial fulfillment of the requirements for the degree of
Master of Science
Horticultural Science
Raleigh, North Carolina 2015
APPROVED BY:
_______________________________ ______________________________ Chair of Advisory Committee Committee Member
Jeanine M. Davis Chris Gunter
________________________________ ________________________________ Committee Member Committee Member
BIOGRAPHY
Jennifer Lee Crumley was born on March 10, 1989 to Billy and Patti Crumley. Raised in the beautiful Appalachian Mountains in East Tennessee, she developed a love of plants after years of helping out on the family farm. The summer before her senior year at the University of Tennessee in Knoxville, Jennifer worked as a student intern at UT’s botanical gardens. In May 2011, Jennifer received her Bachelor’s of Science degree in Public Horticulture from Knoxville, Tennessee. After a year, during which she worked on the family farm and aided in the care of her grandmother, Jennifer returned to school as a Master’s student in the
ACKNOWLEDGMENTS
I would like to thank my advisor, Dr. Jeanine Davis, for her guidance and overwhelming support during my time as a graduate student at NC State. I feel blessed to have had the opportunity to work alongside her and the amazing crew at the Mountain Horticultural Crops Research Station in Mills River, NC. Without their help, this lengthy experiment would have been impossible. I would like to thank Dr. Lijing Zhou for her wonderful advice and
TABLE OF CONTENTS
LIST OF TABLES ...v
LIST OF FIGURES ... vii
LITERATURE REVIEW ...1
CHAPTER1: Seed Source, Growing Location, and Plant Age Affect Growth, Yield, and Phytochemicals in Echinacea purpurea and Echinacea angustifolia ...18
Introduction ...18
Materials and Methods ...22
Results ...28
Discussion...36
REFERENCES ...42
CHAPTER 2: Effect of Seed Source and Seed Size on Germination and Seedling Growth of Echinacea angustifolia ...75
Introduction ...75
Materials and Methods ...78
Results ...81
Discussion...82
LIST OF TABLES
LIST OF FIGURES
Figure 1.1 Example of vigor rating for Echinacea purpurea ...65
Figure 1.2 Echinacea growth stages and bud development ...66
Figure 1.3 Insect damage on Echinacea plants...67
Figure 1.4 Aster Yellows symptoms on Echinacea ...68
Figure 1.5 Echinacea leaf crinkle rating scale ...69
Figure 1.6 Leaf brown spot scale for Echinacea ...70
Figure 1.7 Widespread brown lesions on E. purpurea foliage ...71
Figure 1.8 Cuprofix Ultra 40D application on Echinacea at Mills River ...72
Figure 1.9 Monthly precipitation at Mills River and Reidsville, NC in 2012 and 2013....73
Figure 1.10 Average daily temperatures in 2012 and 2013 in Mills River and Reidsville, NC ...74
Figure 2.1 Echinacea angustifolia seeds in petri dish study 1 ...90
Figure 2.2 Germination of Echinacea angustifolia achenes ...91
LITERATURE REVIEW
The genus Echinacea, in the family Asteraceae, is composed of nine species of herbaceous, perennial flowering plants commonly referred to as coneflowers (Flagel et al., 2008). Native to North America, a variety of species can be found growing in moist, open woods in the southern United States, west to the dry Midwestern prairies, and north into the lower regions of Canada (McKeown, 1999). Echinacea plants are popular both domestically and overseas for use as cut flowers, landscape ornamentals, and in herbal remedies (Li, 1998). United States herbal supplement sales of Echinacea spp. reached almost $16 million dollars in 2012 (Lindstrom et al., 2013) and $28 million in 2013 (Lindstrom et al., 2014), making it one of the most popular medicinal herbs in today’s market. Echinacea purpurea
and Echinacea angustifolia are the two species most used in the natural products industry and
are the subjects of this review. Plant Descriptions
Echinacea purpurea, commonly referred to as purple coneflower,is usually 60 to 180
Echinacea angustifolia is commonly referred to as narrow leaf purple coneflower. When mature, it is 15 to 60 cm tall with stiff, hairy stems. The leaves are lance-shaped and 5 to 30 cm long and 1.5 to 4 cm wide. The flowers are similar to E. purpurea except the rays are much narrower, droopier, and pale purple to light pink. The achenes are brown to black and four-angled. The plant has a tap root (Stevens, 2006b).
Uses
Traditionally, the roots and aerial parts of Echinacea species were used by some Native American tribes in North America to treat a wide range of ailments (Kindscher, 1989). Presently, roots, foliage, flowers, and seeds of E. purpurea and roots of E.
angustifolia are used to create a large number of herbal preparations. The two species are
often combined in formulations. Echinacea is now well known for its use in preventing and treating symptoms of the common cold and flu (Foster, 1991). Echinacea is commonly consumed in capsule or tincture form.
Wild-harvested and Cultivated Plants
Recognized around the world for their medicinal properties, demand has increased dramatically for both E. purpurea and E. angustifolia. Since much of the Echinacea used in the industry is wild-harvested, this increase in demand has caused a decrease in wild
seed and produces high yields of foliage and roots (Foster, 1991). Echinacea angustifolia is a more difficult plant to propagate primarily because of problems with seed germination. Seedlings and first year plants grow slowly and are very small compared to young E.
purpurea plants. Methods such as in vitro propagation have been suggested as a way to
increase cultivation and reduce dependence on wild populations of Echinacea (Onica and Cachita, 2009). If plants are harvested from the wild, as long as portions of the root are left undisturbed certain species of Echinacea, including E. angustifolia,have the ability to regrow (Kindscher et al., 2008).
One way to encourage more cultivation of Echinacea and reduce dependence on wild-populations is to create cultivars that are superior in medicinal properties or consistency than wild-grown plants. This can be accomplished through breeding. There are different ways to identify genetically superior Echinacea plants for medicinal uses. Murch et al. (2006) experimented with clones of E. purpurea in controlled culture systems to compare the chemical profile of individual plant genetics from a population. Comparing cultivated
populations to wild populations is another way to determine plant superiority. When comparing the young roots from wild populations of E. purpurea to cultivated populations,
E. purpurea roots from the cultivated population were shown to yield the highest amount of
cichoric acid (Binns et al., 2002b). A correlation between root age and cichoric acid
Phytochemicals in Echinacea
Echinacea contains several different phytochemicals which have demonstrated
medicinal properties. Echinacea phytochemicals include alkamides and phenolic
constituents, such as echinacoside, cichoric acid, caftaric acid, chlorogenic acid, cynarin, and caffeic acid (Binns et al., 2002b). Alkamides and cichoric acid (a caffeic acid derivative) are believed to be two of most bioactive constituents in Echinacea (Abdoli, Moieni, and Badi, 2013; Bauer, 1998; Mistríková and Vaverková, 2009). These phytochemicals may be responsible for Echinacea’s immune stimulating, anti-fungal, and anti-inflammatory properties (Kumar and Ramaiah, 2011),
Alkamides
One of the most well-known groups of constituents in the Echinacea species is alkamides. Alkamides can be found in many plant families, resulting in many different alkamide structures. These structure differences can also vary between species in the genus
Echinacea, providing an efficient way to identify plant species via high performance liquid
chromatography (HPLC) which separates compounds from a liquid mixture based on the polarity of the molecules (Harborne and Williams, 2004; Rios, 2012). Alkamides are essentially amine groups attached via amide linkages to an unsaturated fatty acid chain. Amines are derivatives of an amino acid group that undergoes decarboxylation (Kraus et al., 2007; Laval and Golding, 2003).
One class of alkamides that is of particular interest to researchers is the
act as fungicides inside the plant when exposed to light conditions (Arnason et al., 2002; Binns et al., 2000). When Echinacea is consumed by humans or animals, alkamides show analgesic, immunostimulatory, antiviral, and anti-inflammatory properties (Birt et al., 2008; Cech et al., 2010; Kumar and Ramaiah, 2011). The anti-inflammatory properties are due to alkamides blocking enzymes that are involved in inflammation (Hinz et al., 2007). The ability of alkamides to reduce or block inflammation in vitro was also observed by Cech et al. (2010).
Alkamides are found in both the aerial parts and roots of E. angustifolia and E.
purpurea although the concentrations vary by location within the different species (Bauer,
1998). Qu et al. (2005) determined that the concentration of alkamides in the roots of E.
purpurea was significantly higher than in the aerial parts of the plant. Woelkart and Bauer
(2007) also found alkamides are higher in the roots of E. angustifolia. In a study by Thomsen et al. (2012), seasonal variations were also observed in root alkamide concentrations in E.
purpurea. In their study, root alkamides were observed in lowest concentration in the autumn
and winter months. Cichoric acid
Cichoric acid can be found in both the roots and aerial parts of both species of
Echinacea (Qu et al., 2005). The concentration of cichoric acid can be affected by flower
age, as young buds were shown to contain higher amounts of cichoric acid compared to the more mature flower heads (Letchamo et al., 1999; Qu et al., 2005).When comparing the roots
contains higher levels of cichoric acid than E. angustifolia (Pellati et al., 2005). According to Thomsen et al. (2012), cichoric acid concentration in the roots is highest in the spring. Evidence for Efficacy of Echinacea Alkamides
Despite being important components in the phytochemical profiles of Echinacea species, until recently it was not known how quickly alkamides become available for the body to metabolize. In order to determine the absorption rate of phytochemicals, particularly alkamides, Woelkart et al. (2005) experimented with extracts taken from two year old roots
of E. angustifolia. They reported that while not every alkamide is available for absorption,
the types that are accessible can be detected in human plasma in as little as ten minutes. Still, regardless of the speed at which some phytochemicals can be absorbed by the body, it does not necessarily mean Echinacea is always effective in treating an illness. In a blind study conducted by Turner et al. (2005), data were collected from 399 volunteers to determine if symptom intensity of the Rhinovirus was reduced by the phytochemical constituents in E.
angustifolia root extracts. The results of this study indicate that treatment with E. angustifolia
root extracts provided no significant relief to the subjects afflicted with the Rhinovirus. Studies like these, however, have serious limitations. For example, the extracts that were used for this study were all obtained from a “single lot of E. angustifolia root” (Turner et al., 2005). There is no mention as to the growing environment, variety (if known), or age of the root in question. Another variable which could have influenced the outcome of the study is the dosage administered to the volunteers. According to a letter written to The New England
Journal of Medicine from the American Botanical Council, the dosage of 900 mg of E.
(Blumenthal and Farnsworth, 2005). In order to have produced a more convincing study, the variables mentioned above should have been examined in depth before implying E.
angustifolia’s ineffectiveness against the Rhinovirus. Alternatively, in a double-blind study
by Goel et al. (2005) it was found that the phytochemicals in a standardized extract from E.
purpurea positively impacted the immune system’s ability to fight off symptoms of a cold.
Melchart and Linde (1999) suggest starting treatment at the onset of symptoms to improve
Echinacea’s effectiveness against colds or other upper respiratory illnesses.
Environmental Effects on Phytochemicals in Echinacea
impact on the amount of potassium available, although a high nitrogen/low potassium combination was recommended by Powell et al. (2001) and Dufault et al. (2003) to improve the plant yield of E. purpurea. If too much potassium is applied, Shalaby et al. (1997b) indicated it may lead to a decrease in the synthesis of carbohydrates.
In addition to nutrients, the type of soil also may affect the growth of the plants. In a study by Powell et al. (2001) plants were shown to have increased root yield when grown in sand-based soils. It was suggested that the roots were able to penetrate the sandy soil much easier than clay-based soils. In the study by Gray et al. (2003), cichoric acid was also shown to increase in Echinacea roots when the plants were subjected to drought conditions;
however, root alkamide content was not shown to be significantly affected. This experiment also determined that despite a high concentration of alkamides in the roots, there is a negative correlation between plant age and root alkamide concentration. Therefore, age is another important factor in determining when or how to harvest Echinacea.
The method used to harvest Echinacea roots can also have an effect on the amount of phytochemicals in the plant sample. When Echinacea roots, particularly those of E.
purpurea, are harvested, many times some of the fragile roots are left in the soil as the plant
is removed. Because it has been found that cichoric acid is often found in greater amounts in the fine roots of Echinacea, care should be taken when harvesting in order to ensure a higher concentration of cichoric acid in the plant sample (Letchamo et al., 2002).
root and rhizome yield of E. purpurea. When the researchers compared these trials to another plant density trial in a climate that was 3 to 4°C warmer, it was found that the root yield from the warmer climate trial more than doubled. This indicates that warmer temperatures may be yet another factor influencing on root yield. The other important result of a higher plant density is the increased risk of disease, which can be caused by environmental factors such as wet soils with poor drainage (Parmenter and Littlejohn, 1997). Often lower planting densities improve air flow; reducing the length of time that moisture from rain or dew sits on the foliage and allowing for greater sun exposure.
produced a higher concentration of phytochemicals than their diseased counterparts (Letchamo et al., 2002).
Ultimately, understanding how the environment affects the chemical properties of
Echinacea can help growers identify the seeds that are not only better suited for the region,
but also produce plants with a high medicinal value. This knowledge will not only provide customers with a higher quality product, but can increase awareness of the benefits of natural medicines derived from our native plants.
Echinacea angustifolia Achene Dormancy
Echinacea angustifolia seeds are often more difficult to germinate than other
Echinacea species such as E. purpurea. Many studies have been conducted to determine the
best method for improving germination in Echinacea. Extensive research has shown that light is an important factor in breaking seed dormancy in E. angustifolia (Feghahati and Reese, 1994; Macchia et al., 2001; Romero et al., 2005; Sari et al., 1999; Sari et al., 2001; Smith-Jochum and Albrecht, 1987). Many of these light studies were done in combination with other treatments such as ethephon (2-chloroethylphosphonic acid) or cold stratification. However, when Macchia et al. (2001) compared the germination of seeds in the presence of light or darkness (without any pretreatment), the study showed that the seeds actually had higher germination under dark conditions. Despite this, light is generally considered a requirement for breaking seed dormancy of E. angustifolia.
their spring-sown counterparts (Smith-Jochum and Albrecht, 1987). Dry, cold stratification
of E. angustifolia was shown to be a good method to increase germination (Chuanren et al.,
2004; Sari et al., 1999; Shalaby et al., 1997a). Cold-moist stratification methods have also been examined. Sari et al. (1999), Romero et al. (2005), and Romero et al. (2010)
recommend the cold-moist stratification method in combination with light. With this treatment combination, E. angustifolia seeds showed improved germination when the temperatures were kept at less than 4º C (Romero et al., 2010; Sari et al., 1999). Macchia et al. (2001) also examined cold-moist stratification methods and concluded that cold-moist stratification and light only affected the rate of germination, not the percentage.
In addition to light and cold-stratification, chemical factors have also been examined. Sari et al. (2001) found that applying ethephon to seeds would improve germination under light conditions. This effect was also noted by Feghahati and Reese (1994) when applied to chilled seeds under constant light. When applied to seeds in darkness, Macchia et al. (2001) found that ethephon did not have a significant effect on germination. Other growth regulators such as gibberellic acid (GA3) and 6-benzylaminopurine (BA) are also reported to increase
germination (Chuanren et al., 2004). However, the results provided by Macchia et al. (2001) do not support the use of GA3 since it did not show any significant germination differences
from the control. The differences between these studies may be due to Chuanren et al. (2004) removing the seed coat before applying GA3 or BA, whereas Macchia et al. (2001) kept the
REFERENCES
Abdoli, M., A. Moieni, and H.N. Badi. 2013. Morphological, physiological, cytological and phytochemical studies in diploid and colchicine-induced tetraploid plants of Echinacea
purpurea (L.). Acta. Physiol. Plant. 35:2075–2083.
Arnason, J.T., S.E. Binns, and B.R. Baum. 2002. Phytochemical diversity and biological activity in Echinacea phytomedicines: challenges to quality control and germplasm
improvement. p. 9-17. In: M.S. Meskin. (ed.). Phytochemicals in nutrition and health. CRC Press, Boca Raton, FL.
Bauer, R. 1998. Echinacea: biological effects and active principles. p. 140-157. In: L.D. Lawson, R. Bauer. (eds.). Phytomedicines of Europe: chemistry and biological activity. American Chemical Society Symposium Series 691. Washington, DC.
Bauer, R. 2000. Chemistry, Pharmacology and clinical applications of Echinacea products. p. 45-74. In: G. Mazza and B.D. Oomah (eds.). Herbs, botanicals, and teas. CRC Press, Boca Raton, FL.
Binns, S.E., B. Purgina, M.L. Smith, L. Johnson, and J.T. Arnason. 2000. Light-mediated antifungal activity of Echinacea extracts. Planta Med. 66:1-4.
Binns, S.E., J.T. Arnason, and B.R. Baum. 2002a. Phytochemical variation within populations of Echinacea angustifolia (Asteraceae). Biochem. Sys. Ecol. 30:837-854. Binns, S.E., B.R. Baum, J.T. Arnason, and J.F. Livesey. 2002b. Phytochemical variation in
Echinacea from roots and flower heads of wild and cultivated populations. J. Agr. Food
Chem. 50(13):3673-3687.
Birt, D.F., M.P. Widrlechner, C.A. Lalone, L. Wu, J. Bae, A.K.S. Solco, G.A., Kraus, P.A. Murphy, E.S. Wurtele, Q. Leng, S.C. Hebert, W.J. Maury, and J.P. Price. 2008. Echinacea in infection. Amer. J. Clin. Nutr. 87:488–492.
Blumenthal M. and N.R. Farnsworth. 2005. Echinacea angustifolia rhinovirus infections [letter]. New Engl. J. Med. 353(18):1971-1972.
Cech, N.B., V. Kandhi, J.M. Davis, A. Hamilton, D. Eads, and S.M. Laster. 2010. Echinacea and its alkylamides: effects on the influenza A-induced secretion of cytokines, chemokines, and PGE2 from RAW 264.7 macrophage-like cells. Intl. Immunopharmacol. 10:1268–1278. Chuanren, D., W. Bochu, L. Wanqian, C. Jing, L. Jie, and Z. Huan. 2004. Effect of chemical and physical factors to improve the germination rate of Echinacea angustifolia seeds.
Dufault, R.J., G., B. Ward, B.M. Shepard, J. Rushing, and R. Hassell. 2003. Influence of fertilizer on growth and marker compound of field-grown Echinacea species and feverfew. Scientia Hort. 98(1):61-69.
Feghahati, S.J. and R.N. Reese. 1994. Ethylene-, light-, and prechill-enhanced germination of
Echinacea angustifolia seeds. J. Amer. Soc. Hort. Sci.119(4):853-858.
Flagel, L.E., R.A. Rapp, C.E. Grover, M.P. Widrlechner, J. Hawkins, J.L. Grafenberg, I. Alvares, G.Y. Chung, and J.F. Wendel. 2008. Phylogenetic, morphological, and
chemotaxonomic incongruence in the North American endemic genus Echinacea. Amer. J. Bot. 95(6):756–765.
Foster, S. 1991. Echinacea: nature's immune enchancer. Healing Arts Press, Rochester, VT. Goel, V., R. Lovlin, C. Chang, J.V. Slama, R. Barton, R. Gahler, R. Bauer, L.
Goonewardene, and T.K. Basu. 2005. A proprietary extract from the Echinacea plant
(Echinacea purpurea) enhances systemic immune response during a common cold.
Phytother. Res. 19(8):689-694.
Gray, D.E., S.G. Pallardy, H.E. Garrett, and G.E. Rottinghaus. 2003. Acute drought stress and plant age effects on alkamide and phenolic acid content in purple coneflower roots. Planta Med. 69(1):50-55.
Greenfield, J. and J.M. Davis. 2012. Echinacea (Echinacea angustifolia L.). Medicinal Herb Production Guide. N.C. Consortium on Natural Medicines. UNC-Chapel Hill and NC State University. 22 April 2015
<http://www.ces.ncsu.edu/fletcher/programs/herbs/crops/medicinal/E%20angustifolia%20fin al%20article%202012.pdf>
Harborne, J.B. and C.A. Williams. 2004. Phytochemistry of the genus Echinacea. p. 55-71. In: S.C. Miller, He-ci Yu (eds.). Echinacea: The genus Echinacea. CRC Press, Boca Raton, FL.
Hinz, B., K. Woelkart, and R. Bauer. 2007. Alkamides from Echinacea inhibit
cyclooxygenase-2 activity in human neuroglioma cells. Biochem. Biophys. Res. Commun. 360:441-446.
Kindscher, K. 1989. Ethnobotany of purple coneflower (Echinacea angustifolia, Asteraceae) and other Echinacea Species. Econ. Bot. 43(4):498-507.
Kraus, G.A, J. Bae, L. Wu, and E. Wurtele. 2007. Synthesis and natural distribution of anti-inflammatory alkamides from Echinacea. National Institute of Health. Molecules
11(10):758–767.
Kumar, K.M. and S. Ramaiah. 2011. Pharmacological importance of Echinacea purpurea. Intl. J. Pharma. Bio. Sci. 2(4):304-314.
Laval, G. and B.T. Golding. 2003. One-pot sequence for the decarboxylation of α -amino acids. Synlett. 4:542–546.
Letchamo, W., J. Livesey, T.J. Arnason, C. Bergeron, and V.S. Krutilina. 1999. Cichoric acid and isobutylamide content in Echinacea purpurea as influenced by flower developmental stages. p. 494-498. In: J. Janick (ed.). Perspectives on new crops and new uses. ASHS Press, Alexandria, VA.
Letchamo, W., L.V. Polydeonny, N.O. Gladisheva, T.J. Arnason, J. Livesey, and D.V.C. Awang. 2002. Factors affecting Echinacea quality. ASHS Press, Alexandria, VA. 514-521. Li, T.S.C. 1998. Echinacea: cultivation and medicinal value. HortTechnology 8(2):122-129. Lindstrom, A., C. Ooyen, M.E. Lynch, and M. Blumenthal. 2013. Herb supplement sales increase 5.5% in 2012: herbal supplement sales rise for 9th consecutive year; turmeric sales jump 40% in natural channel. HerbalGram 99:60-65.
Lindstrom, A., C. Ooyen, M.E. Lynch, M. Blumenthal, and K. Kawa. 2014. Sales of herbal dietary supplements increase by 7.9% in 2013, marking a decade of rising sales; turmeric supplements climb to top ranking in natural channel. HerbalGram 103:52-56.
Macchia, M.M., L.L. Ceccarini, and L.G. Angelini. 2001. Methods to overcome seed dormancy in Echinacea angustifolia. DC. Scientia Hort.89(4):317-324.
McKeown, K.A. 1999. A review of the taxonomy of the genus Echinacea. p. 482-489. In: J. Janick (ed.), Perspectives on new crops and new uses. ASHS Press, Alexandria, VA.
Melchart, D. and K. Linde. 1999. Clinical investigations of Echinacea phytopharmaceuticals. p. 105-118. In: H.Wagner (ed.). Immunomodulatory Agents from Plants. Birkhäuser Verlag, Basel.
Mistríková, I. and S. Vaverková. 2007. Morphology and anatomy of Echinacea purpurea, E.
Mistríková, I. and S. Vaverková. 2009. Patterns of variation in lipophilic and hydrophilic constituents in flower developmental stages of Echinacea purpurea (L.) Moench cultivated in Slovakia. Plant Soil Environ. 55(2):70–73.
Murch, S.J., S.E. Peiris, W.L. Shi, S.M.A. Zobayed, and P.K. Saxena. 2006. Genetic diversity in seed populations of Echinacea purpurea controls the capacity for regeneration, route of morphogenesis and phytochemical composition. Plant Cell. Rep. 25(6):522-532. Onica, D. and D. Cachita. 2009. The Echinacea species. J. Hort. For. Biotechnol. 13:268-271.
Parmenter, G.A. and R.P. Littlejohn. 1997. Planting density effects on root yield of purple coneflower (Echinacea purpurea (L.) Moench). N. Z. J. Crop Hort. Sci. 25(2):169-175. Pellati, F., T. Lasseigne, M. Melegari, and S. Benvenuti. 2005. Variability in the composition of anti-oxidant compounds in Echinacea species by HPLC. Phytochemical Anal. 16(2):77-85.
Powell, E.E., M.R.L. Bantle, T.G. Crowe, and D.J. Carrier, D.J. 2001.
Echinacoside and alkamide distribution in Echinacea angustifolia root: root depth and growing condition. J. Nutraceut. Function. Med. Foods 3(3):95-106.
Qu, L.P., Y. Chen, X.P. Wang, R. Scalzo, and J.M. Davis. 2005. Patterns of variation in alkamides and cichoric acid in roots and aboveground parts of Echinacea purpurea (L.) Moench. HortScience 40(5):1239-1242.
Ražić, S., A. Onjia, and B. Potkonjak, 2003. Trace elements analysis of Echinacea purpurea - herbal medicinal. J. Pharm. Biomed. Anal. 33(4):845-850.
Rebek, E.J. 2008. Be on the lookout for aster yellows. 22 April 2015 <http://entoplp.okstate.edu/pddl/2008/PDIA7-15.pdf>
Rios, M.Y. 2012. Natural alkamides: pharmacology, chemistry and distribution. p. 107-144. In: O. Vallisuta (ed.). Drug discovery research in pharmacognosy. InTech. 30 May 2013 <http://www.intechopen.com/books/drug-discovery-research-in-pharmacognosy/natural-alkamidespharmacology-chemistry-and-distribution>
Romero, F.R., Y. Liu, P. Murphy, K. Delate, and D.J. Hannapel. 2010. Horticultural and biochemical variations due to seed source and production methods in three Echinacea spp. J. Herbs Spices Med. Plants 16(3-4):167-192.
Sari, A.O., J.E. Simon, and M.R. Morales. 1999. Echinacea angustifolia: an emerging medicinal. p. 490-493. In: J. Janick (ed.). Perspectives on new crops and new uses. ASHS Press, Alexandria, VA.
Sari, A.O., J.E. Simon, and M. R. Morales. 2001. Ethephon can overcome seed dormancy and improve seed germination in purple coneflower species Echinacea angustifolia and E.
pallida. HortTechnology 11(2):202-205.
Seemannová, Z., I. Mistríková, and S. Vaverková. 2006. Effects of growing methods and plant age on the yield, and on the content of flavonoids and phenolic acids in Echinacea
purpurea (L.) Moench. Plant Soil Environ. 52(10):449-453.
Shalaby, A.S., A.S. El-Khayat, S.F. Hindawy, E.A. Agina, and S.E. El-Gengaihi. 1997a. Response of Echinacea to some agricultural practices. J. Herbs Spices Med. Plants 4(4):59-67.
Shalaby, A.S., A.S. El-Khayat, S.F. Hendawy, S.E. El-Gengaihi, and E.A. Agina. 1997b. Growth and yield of Echinacea purpurea L. as influenced by planting density and
fertilization. J. Herbs Spices Med. Plants 5(1):69-76.
Smith-Jochum, C.C. and M.L. Albrecht. 1987. Field establishment of
three Echinacea species for commercial production. Acta Hort. 208:115-120. Stevens, M. 2006a. Eastern purple coneflower. Echinacea purpurea (L.) Moench. USDA/NRCS Plant Guide. 22 April 2015.
<http://plants.usda.gov/plantguide/pdf/cs_ecpu.pdf>
Stevens, M. 2006b. Purple coneflower. Echinacea angustifolia D.C. USDA/NRCS Plant Guide. 22 April 2015. <http://plants.usda.gov/plantguide/pdf/cs_ecan2>
Thomsen, M.O., X. Fretté, K.B. Christensen, L.P. Christensen, and K. Grevsen. 2012. Seasonal variations in the concentrations of lipophilic compounds and phenolic acids in the roots of Echinacea purpurea and Echinacea pallida. J. Agric. Food Chem. 60:12131-12141. Turner, R.B., R. Bauer, K. Woelkart, T.C. Hulsey, and J.D. Gangemi. 2005. An evaluation of
Echinacea angustifolia in experimental rhinovirus infections. N. Engl. J. Med.
Woelkart, K., C. Koidl, A. Grisold, J.D. Gangemi, R.B. Turner, E. Marth, and R. Bauer. 2005. Bioavailability and pharmacokinetics of alkamides from the roots of Echinacea
angustifolia in humans. J. Clin. Pharmacol. 45(6):683-689.
Woelkart, K. and R. Bauer. 2007. The role of alkamides as an active principle of Echinacea. Planta Med. 73(7):615-623.
CHAPTER 1
Seed Source, Growing Location, and Plant Age Affect Growth, Yield, and Phytochemicals in Echinacea purpurea and Echinacea angustifolia. Abstract
Three year field trials were initiated in 2012 in the southern mountains (Mills River) and upper piedmont (Reidsville) regions of North Carolina to evaluate the effect of seed source, plant age, and environment on the growth, yield, and phytochemical profiles
of Echinacea purpurea and Echinacea angustifolia. The studies were established as three
year studies; the results from the first two years are reported here. Location had a significant effect on flower number for both species. Dried aerial yields for E. purpurea were greater in the mountains than in the piedmont in both years, but there was no difference between locations for dried root yields. Echinacea angustifolia did not exhibit a clear response to location. Seed source had no effect on E. purpurea root yield. For E. angustifolia, however, one seed source provided significantly higher root yields, but DNA analysis revealed it also contained genetic material from E. pallida. Plants grew bigger and matured faster in the second growing season. Aerial cichoric acid content in E. purpurea decreased with plant age while root cichoric acid increased. Root alkamide levels also decreased as the plants aged. Introduction
The genus Echinacea, in the family Asteraceae, contains nine species native to North America(Flagel et al., 2008). Plants from the genus Echinacea are commonly used as
range of ailments including snake bites and inflammation, and as an eyewash and local anesthetic (Kindscher, 1989). Now Echinacea is commonly found in a capsule or tincture form and used for treating symptoms associated with the common cold or flu. According to Lindstrom et al. (2013), domestic sales of Echinacea spp. reached almost $16 million in 2012, making it one of the most popular herbs in today’s herbal supplement market.
The two Echinacea species most often used as dietary supplements are E. purpurea
and E. angustifolia.Echinacea purpurea is 60 to 180 cm tall with serrated ovate to lanceolate
shaped leaves, reddish-purple to lavender flowers with large raised disks, and a fibrous root system. Echinacea angustifolia tends to be slightly smaller with narrower leaves and pale-purple to pink flowers with long narrow rays, and a taproot (Foster, 1991; Kindscher, 1989; Mistríková and Vaverková, 2007; Seemannová, Mistríková. and Vaverková, 2006). Both species tend to be drought tolerant once established (Li, 1998).
The aerial parts and roots of both E. purpurea and E. angustifolia contain phytochemicals that are the compounds that make the plants valued for their medicinal properties. The concentrations of these phytochemicals vary between the different species and are affected by the environment and location where the plants are grown (Bauer, 1998). Alkamides are in both the aerial parts and roots of E. angustifolia and E. purpurea. While cichoric acid is found in the roots and aerial parts of many Echinacea species, according to Letchamo et al. (1999), cichoric acid is in greatest concentration in the young buds of E.
purpurea plants. As the flower heads reach maturity, the level of cichoric acid starts to
roots of E. angustifolia, the roots of E. purpurea contain greater amounts of cichoric acid (Pellati et al., 2005).
The environment where the plants are grown can greatly affect the quality of the chemical constituents (Gray et al., 2003). A study conducted on E. purpurea looked at the effect of different rates of K fertilizer on plant growth (Dufault et al., 2003). A low N-P-K fertilizer was recommended when growing E. purpurea for its roots, as a high rate reduced root growth. Wu et al. (2007) determined that Echinacea roots produce higher amounts of cichoric acid when grown in an environment with limited nutrition availability. In addition to nutrients, the type of soil also may affect the growth of the plants. Echinacea can be found growing naturally in rocky soils that are generally considered undesirable for any crop production (Kindscher et al., 2008). In one study, Echinacea plants had increased root yield when grown in sand-based soils (Powell et al., 2001). It was suggested that the roots were able to penetrate the sandy soil much easier than clay-based soils.
Cichoric acid was shown to increase in Echinacea roots when the plants were subjected to drought conditions; however, root alkamide content was not shown to be
significantly affected (Gray et al., 2003). This experiment also determined that despite a high concentration of alkamides in the roots, there is a negative correlation between plant age and root alkamide concentration. Therefore, age is another important factor in determining when or how to harvest Echinacea.
When the researchers compared these trials to another plant density trial in a climate that was 3° to 4°C degrees warmer, it was found that the root yield from the warmer climate trial was more than doubled. This indicates that growing temperatures also influence root yield. However, an increase in root growth does not necessarily indicate increased alkamide concentration, as observed by Romero et al. (2009).
Another important result of using higher planting densities is the increased chance of disease, which can be caused by environmental factors such as wet soils with poor drainage (Parmenter and Littlejohn, 1997). Often a lower planting density provides improved air flow around the plants, which can help keep moisture from sitting on the foliage and allows for greater sun exposure.
In addition to disease, insect damage can be an issue for a variety of reasons, such as destroying Echinacea crops grown for cut flowers or opening the plant up to infection
forming a “witch’s broom”, or green flower rays and cones (Rebek, 2008; Romero et al., 2010).
Recognized around the world for their medicinal properties, demand has increased for both E. purpurea and E. angustifolia. Since Echinacea is often still wild-harvested in parts of its native range, this increase in demand can cause a decrease in wild populations (Li, 1998), particularly that of E. angustifolia (Kindscher et al., 2008). However, due to improvements made in cultivation of E. angustifolia, dependency on wild plants has been greatly reduced in recent years (Greenfield and Davis, 2012). Adoption of such methods as in vitro propagation to increase efficiency of propagation has been suggested to help reduce dependence on wild populations (Onica and Cachita, 2009). More sustainable wild-harvest methods could also reduce the negative effects on wild populations. For example, if portions of the root are left undisturbed certain species of Echinacea have the ability to regrow (Kindscher et al., 2008).
The objective of this research was to determine the effect of growing environment, plant age, and seed source on growth, yield, and production of several important
phytochemicals in E. purpurea and E. angustifolia. Materials and Methods
Location
Field studies on E. purpurea and E. angustifolia were conducted over the summer and fall of 2012 and 2013 at two different locations in North Carolina, USA. One field site was located at the Mountain Horticultural Crops Research and Extension Center in Mills River in the western mountains of North Carolina at longitude -82.557 and latitude 35.4267 at
average precipitation is 128 cm of rain and 28 cm of snow (USClimateData.com). The other field site was located at the Upper Piedmont Research Station in Reidsville in the piedmont in north-central North Carolina at longitude -79.6947 and latitude 36.3825 at approximately 271 meters in elevation. The annual temperature is 14.4°C and annual precipitation is 118 cm of rain and 23 cm of snow (USClimateData.com).
Experimental Design
Two identical field studies were established, one for E. purpurea and one for E.
angustifolia. For each species, six seed sources were evaluated with the plants being
harvested for phytochemical analysis after one, two, or three years of growth. Each species was in a nested design consisting of a split-plot study at the two locations. The main plots were harvest year and the subplots were seed sources. There were three replications. Each subplot consisted of four rows of 12 plants each with the outer two rows serving primarily as guard rows. To accommodate cultivation equipment at the different sites, the subplots at the Mills River site were 4.3 m x 3 m with rows 107 cm apart. At the Reidsville site the subplots were 4.9 m x 3 m with rows 122 cm apart. Plants were 25 cm apart in the row.
Seed Sources
MN, USA), Native American Seed (Junction, TX, USA), Pharmasaat (Artern, Germany), S&S Seeds (Carpinteria, CA, USA), and Stock Seed Farm (Murdock, NE, USA). Echinacea
angustifolia seeds were ordered from the commercial seed companies Prairie Moon Nursery
(Winona, MN, USA), Native American Seed (Junction, TX, USA), S&S Seeds (Carpinteria, CA, USA), Wind River Seed (Manderson, WY, USA), and Stock Seed Farm (Murdock, NE, USA).
Growing Methods
Transplants of both of the Echinacea species were grown by a local nursery (Gardens of the Blue Ridge, Inc., Pineola, NC, USA). Seedswere sown on April 4, 2012 in 200 cell flats containing Fine Germinating Mix (Fafard®, Agawam, MA, USA) before being
transferred to 72 cell flats containing Fafard 3b Mix (Fafard®, Agawam, MA, USA) on May 14, 2012. Fertilizer (15-30-15, Grow More Inc., Gardena, CA, USA), proportioned at a rate of 15,000:1 was provided through the irrigation during early growth. No pretreatment methods were used to break seed dormancy. The approximate germination percentage was recorded by the grower (Table 1.1).
Soil samples were collected from the fields to determine initial soil fertility, soil type, and texture (A & L Laboratories, Inc., Memphis, TN, USA). The Mills River site was a clay loam soil and the Reidsville site had sandy clay loam and sandy loam soils for the E.
purpurea and E. angustifolia fields, respectively. The preplant fertilizers applied were
soil fertility for both sites. The target soil fertility was a pH of 6 to 7, 56 kg N/ ha, and 74 kg K2O/ha.
Echinacea purpurea was transplanted by hand to the field in Mills River on June 12,
2012 and in Reidsville on June 14, 2012, E. angustifolia was transplanted by hand to the field in Mills River on July 6, 2012 and in Reidsville on July 11, 2012. The transplants were watered in during the planting process to improve transplant success. Overhead irrigation was used as needed throughout the growing season at both sites. To prevent deer herbivory, electric wire fencing was erected around the fields at the Reidsville location. Mechanical cultivation was used for weed control early in the growing season. Once the Echinacea plants became too tall for the tractor to pass without resulting in crop damage, the weeds were removed by hand. Paraquat (Firestorm®, Chemtura Corporation, Middlebury, CT, USA) and Surflan (United Phosphorus, Inc., King of Prussia, PA, USA) were used to control weeds on the field perimeters in Mills River, NC.
Cuprofix Ultra 40D (United Phosphorus, Inc., King of Prussia, PA, USA) was applied on August 1, 2012 to control a foliar blight which affected both Echinacea species in Mills River. The blight could not be identified at the Plant Disease and Insect Clinic (North Carolina State University, Raleigh, NC, USA) and did not reoccur.
Plant Rating
1.6). Vigor was rated for the entire plot, rather than individual plants. Plant height was measured from soil level to the top of the tallest flower or leaf. Growth stages were determined using the diagrams in Figure 2 and the descriptions by Brovelli et al (2005) (Table 1.2). Damage caused by insects consisted of holes and leaf blemishes caused by chewing and leaf tunneling insects. Leaf deformities, such as “leaf curl”, occurred when the leaf was curled in on itself but was not caused by insect activity. Leaf “crinkle” was the term given to plants that produced leaves with a wrinkled surface. The term “brown spots” was given to indicate the presence of brown blemishes on the leaf surface.
A rating scale of 1 to 4 was used for insect damage, leaf crinkle, curl, and brown spot. Rating of “1” means none, “2” means <25% of the total plot leaf area is affected, “3” means 25% to 50% of the total plot leaf area is affected, and “4” means 50% to 100% of the total plot leaf area is affected. A rating scale of 1 to 3 was used for plot vigor where “1” was used to indicate plots that are considered poorly adapted to the field, “2” indicates average growth and appearance, and “3” indicates a plot which contains healthy and vigorous plants which are well adapted to field conditions.
Ratings were taken seven times during the season in 2012 but only twice during 2013 because record setting rains created excessive mud in the Mills River fields. To prevent damage to the plants and root systems, unnecessary entry into the fields was avoided. Harvest
place on November 5 and November 14 for Mills River and Reidsville, respectively. In 2013, the aerial parts of E. purpurea plants were harvested in Reidsville, NC on June 19 and in Mills River, NC on June 25 and roots of both species were harvested in Reidsville on October 31 and in Mills River on November 12. The aerial parts of E. purpurea were harvested once buds were produced but before the flowers matured. Ideal flower bud stages for harvest ranged from bud enlargement in stage 4 to stage 6 buds with erect green or white ligules (Figure 1.2). Once at least 50% of the data row plants reached bud stage 4, the plot was considered ready for harvest. The plants were harvested both by hand with hand shears and with the use of a gas powered weed eater (various makes and models) with a blade attachment. Plants were cut approximately 15 cm above the ground and the stems, leaves, and flower cones were cut into smaller pieces to facilitate even drying. The methods used for aerial harvest of E. purpurea were used to simulate the commercial mechanized harvesting of
E. purpurea at Trout Lake Farms in Washington, USA. Plants were gathered into brown
paper bags, weighed, and dried in a forced-air dryer (custom built) at 43°C, and reweighed. In the fall, roots of both species were harvested by digging with garden forks. Very large roots were cut to facilitate washing. The roots were tumbled in a KPRO model concrete mixer (Kushlan Products, LLC, Sugar Land, TX, USA) with a small amount of water and then rinsed with a high pressure hose. The roots were air dried till surface moisture was gone, gathered in brown paper bags, weighed, dried in a forced-air dryer at 43°C, and reweighed. Phytochemical Analysis
phytochemical analysis was performed using HPLC. The phytochemicals cichoric acid and alkamides were analyzed according to methods described by Rana and Chandra, (2006). Statistical Analysis
Data were analyzed using the Proc Mixed and Proc GLM Procedures of the SAS statistical package 9.3 (SAS Institute, Cary, NC, USA). Means separations were conducted using PDMIX800 with significance defined as alpha = 0.05 (Saxton, 1998.). All data were considered normal and did not have to be transformed for statistical analysis.
Study Years Included
The overall study was designed to be three years long; the plants were harvested after one, two, and three years of growth. Only the first two years of harvest are included in this Master of Science degree thesis project. All three years of data will be included for the final publications on the project.
Results
First Year Growth in 2012:
On July 30, 2012, brown lesions were observed on the foliage of the plants at the
mountain site in Mills River, NC. The lesions were widespread across the field, affecting
both Echinacea species (Figure 1.7). Plant samples were taken from both species and sent to
the North Carolina Plant Disease and Insect Clinic in Raleigh, NC but a positive diagnosis
could not be made. The problem was assumed to be fungal in nature, so in an attempt to
prevent further spread of the lesions a copper fungicide product was applied on August 1and
put out healthy new growth and outgrow the affected tissue. The brown lesions were not
considered the same as the “brown spots” recorded over the two year study.
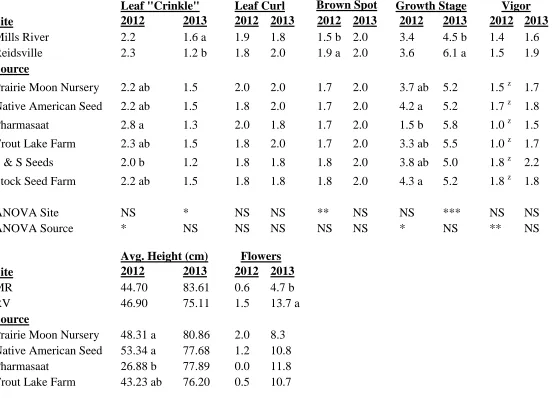
Echinacea purpurea
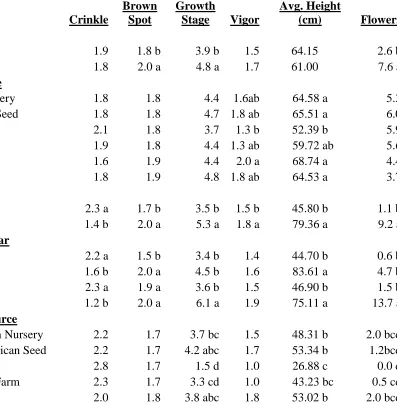
The September 7 ratings of the various factors observed on E. purpurea in 2012 are
presented here.Growing location and seed source had no effect on the amount of insect
damage or Aster Yellows symptoms sustained by the plants (data not shown). The
appearance of the foliar deformity, leaf curling, was also not related to site or seed source
(Table 1.3). Leaf “crinkle”, another leaf anomaly, was not different between the two growing
locations, but plants grown from Pharmasaat seed had the most “crinkled” leaves and
significantly more than plants grown from seeds from S&S Seeds. All other sources were
statistically similar. There was more leaf brown spot on plants grown in Reidsville compared
to their counterparts in Mills River with a rating of 1.9 and 1.5 out of 4, respectively (Table
1.3). Seed source, however, had no effect on brown spot occurrence in E. purpurea.
Seeds from Prairie Moon Nursery, Native American Seed, S & S Seeds, and Stock
Seed Farm produced the tallest plants with average heights of 48.31 cm, 53.34 cm, 53.02 cm,
and 50.01 cm, respectively (Table 1.3). The shortest plants were from Pharmasaat seed with
an average height of only 26.88 cm. Plants grown from seed from Trout Lake Farm and
Pharmasaat were the least vigorous (Table 1.3). Growing location and seed source had no
effect on flower number.
On September 7 there were significant differences in growth stage among plants from
respectively. Plants from Pharmasaat seed were significantly less mature with a growth stage
of 1.5. All other sources were similar, ranging from a growth stage of 3.3 to 3.8.
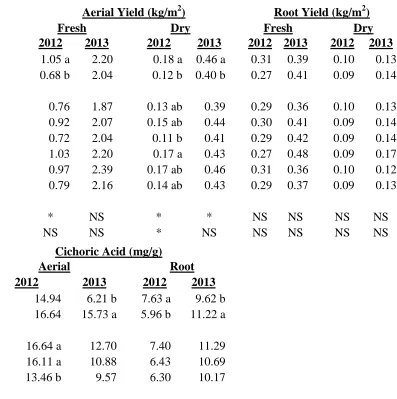
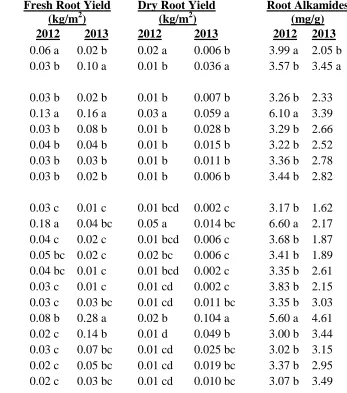
Fresh and dry aerial yield were affected by growing location. Plants grown in Mills River produced higher aerial yields than those in the Reidsville location (Table 1.4). Dry aerial yield was also affected by source, with plants grown from Trout Lake Farm seeds producing significantly more than plants grown from Pharmasaat seed. Plants from other seed sources produced similarly. Neither site nor source significantly affected dry root yield.
The phytochemicals in E. purpurea are presented on a milligram per gram dried tissue basis. Aerial cichoric acid levels were not affected by growing location, but plants grown from Pharmasaat seed contained significantly less aerial cichoric acid than plants from any of the other seed sources except for Trout Lake Farm (Table 1.4). In contrast, site had a significant effect on root alkamides and root cichoric acid. Plants grown in Mills River
produced more root cichoric acid but less root alkamides than their counterparts grown in
Reidsville.
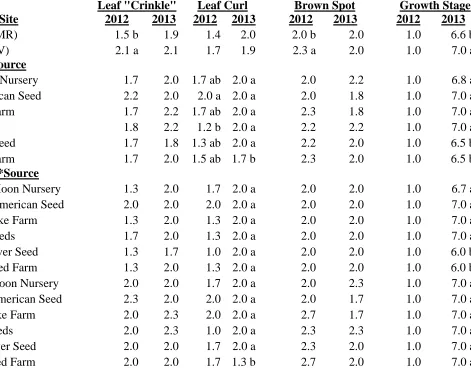
Echinacea angustifolia:
The September 7 ratings of the various factors observed on E. angustifolia in 2012
are presented here. Neither growing location nor source had any effect on the amount of
Aster Yellows symptoms or insect damage (data not shown). Leaf “crinkle” was significantly
influenced by growing location (Table 1.5) with Reidsville grown plants having more
“crinkled” leaves than their counterparts in Mills River. Leaf curling, however, was affected
by seed source (Table 1.5). Plants grown from seed from Native American Seed were
different from any of the other seed sources. Plants grown in Reidsville had more leaf brown
spot than plants grown in Mills River (Table 1.5). Echinacea angustifolia grew very slowly
the first season in the field and on September 7 all the plants were young with no flower buds
present. There was no difference in growth stage among any of the treatments.
Plant vigor was the only factor that had a significant site x source interaction (Table
1.5). Plants grown from Native American Seed at both sites were the most vigorous. The
least vigorous were Prairie Moon Nursery and Stock Seed Farm plants from Mills River, and
Trout Lake Farm plants from Reidsville. Seed source had an effect on plant height with the
tallest plants produced from Native American Seed seed and the shortest plants from Trout
Lake Farm seed (Table 1.5). There was neither location nor seed source effects on the
number of flowers per plot (Table 1.5).
The phytochemicals for E. angustifolia are presented on a milligram per gram dried tissue basis. There was a significant site x seed source interaction for E. angustifolia fresh and dry root yield per unit land area as well as root alkamides (Table 1.6). The highest fresh
and dry root yield was obtained from plants grown in Mills River from Native American
Seed seeds. They were significantly higher than any other treatment. The second highest
fresh and dry root yields were from plants grown in Reidsville from Native American Seed
seeds, but they were not significantly different from the Mills River grown plants produced
from S&S Seeds and Wind River Seed. The lowest yields at Mills River were obtained from
plants grown from seed from Prairie Moon, Trout Lake Farm, and Stock Seed. At Reidsville,
Alkamides were produced in much higher amounts in both locations in roots of plants
grown from seed from Native American Seed plants. All the others were not different from
the other regardless of site or seed source.
Second Year Growth in 2013:
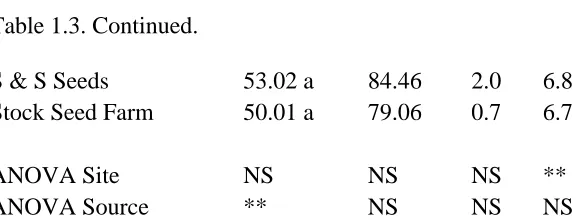
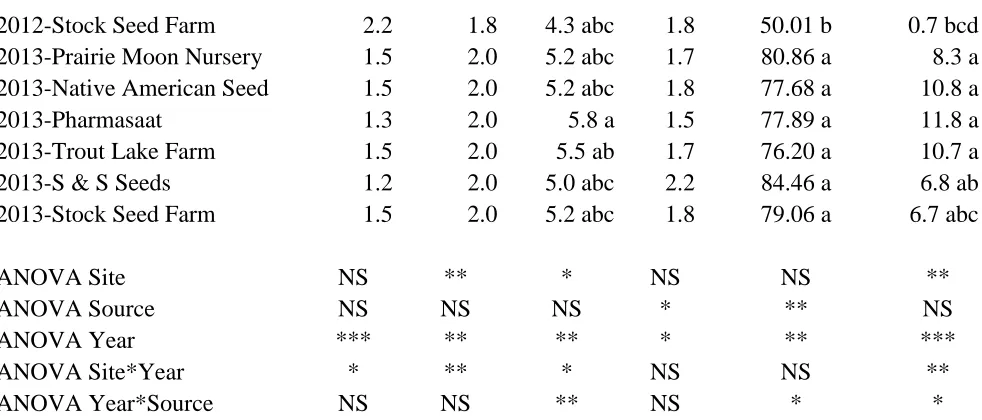
Echinacea purpurea:
The growth rating data presented here are from June 6, 2013 in Mills River and June
10, 2013 in Reidsville. There was no effect of growing location or seed source on symptoms
of Aster Yellows or insect damage (data not shown) or on leaf curl, brown spot, vigor, or
plant height (Table 1.3). Plants grown in Mills River had leaves with a more “crinkled”
appearance than their counterparts in Reidsville. In early June, the average growth stage was
more advanced and there were more flowers in Reidsville than Mills River
Echinacea angustifolia:
The data presented are from June 6, 2013 in Mills River and June 10, 2013 in
Reidsville. Growing location and source did not affect the incidence of Aster Yellows or
insect damage (data not shown), or leaf “crinkle” or brown spot (Table 1.5). The F-test
indicated there was a significant site x source interaction for leaf curl, but the simple effect
means and the Tukey mean separation indicate that all the treatment means were the same
except for the Reidsville Stock Seed Farm treatment which was significantly less (Table 1.5).
There was a highly significant site x source interaction for growth stage in early June. The
plants grown from seed from Wind River Seed and Stock Seed Farm at Mills River were
significantly less mature than all other treatments (Table 1.5). For plant vigor, plants grown
from Native American Seed were the most vigorous. Plants grown at Reidsville from seed
obtained from Native American Seed were significantly taller than from any other treatment.
For flower number, plants at Reidsville produced more flowers than plants at Mills River and
plants grown from seed from Native American Seed produced significantly more flowers
than any other seed source (Table 1.5).
There was a significant site x source interaction for fresh and dry root yield per unit
land area (Table 1.6). In both instances, plants grown from the Native American Seed
produced significantly higher yields than the other treatments which were all statistically
similar. Root alkamides were produced in higher amounts in Reidsville compared to Mills
Year to Year Comparison
Echinacea purpurea:
Leaf “crinkle” was more prevalent in the field in 2012 compared to 2013 with a rating of 2.3 and 1.4, respectively (Table 1.7). No differences were observed for the presence of Aster Yellows or leaf curl (data not shown). Plants grown in 2013 experienced more brown leaf spots than plants grown during the 2012 season (Table 1.7). Overall, plants grown in Reidsville were more affected by brown spot than those from Mills River with a rating of 2.0 and 1.8, respectively (Table 1.7). Plants matured significantly faster in 2013 compared to 2012, reaching a growth stage of 5.3 by early June in 2012 compared to a growth of stage of only 3.5 in early September in 2012 (Table 1.7). Plant vigor was affected by year with 2013 plants being slightly but significantly more vigorous than 2012 with a rating of 1.8 compared to 1.5, respectively (Table 1.7).
Across both locations, the two year old plants were all taller than the first year old plants with an average height of 79.36 cm compared to 45.80 cm, respectively (Table 1.7). There were also more flowers per plot in the two year old plots (9.2) compared to the one year old plants (1.1) (Table 1.7). There was no difference in height of plants between locations when years were combined, but average number of flowers per plot was significantly higher at Reidsville (7.6) than Mills River (2.6) (Table 1.7).
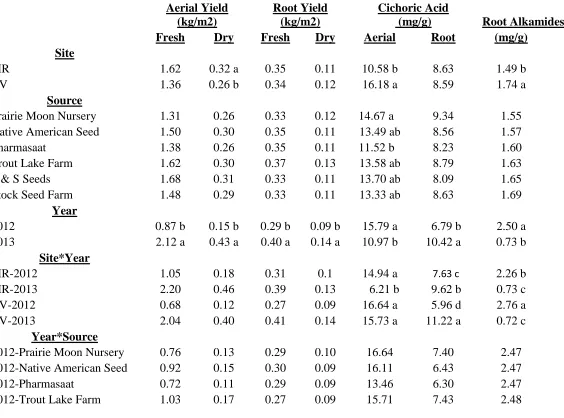
different between years but not as dramatically so as the aerial yields. Fresh root yields increased 1.4 fold (0.29 kg/m2 in 2012 and 0.40 kg/m2 in 2013) and dry root yields increased 1.6 fold (0.09 kg/m2 in 2012 and 0.14 kg/m2 in 2013) (Table 1.8). There was not a significant difference in fresh or dried root yield between locations when years were combined.
Aerial cichoric acid levels across all sources and both sites were significantly reduced from 2012 to 2013, 15.79 mg/g compared to 10.97 mg/g, respectively (Table 1.8). Root cichoric acid levels were just the opposite with 6.79 mg/g in 2012 and 10.42 mg/g in 2013 (Table 1.8). Root alkamide levels were also severely and significantly reduced from 2012 to 2013, with 2.50 mg/g and 0.73 mg/g, respectively (Table 1.8).
Echinacea angustifolia
Native American Seed plants produced the most flowers with 76.3, and were significantly more than Stock Seed Farm which plants produced the least with 15.4 (Table 1.9).
There was a significant difference observed for the interaction of growing location, year, and seed source for fresh root and dry root yield. Native American Seed plants
produced the greatest fresh and dry root yield in Reidsville in 2013 (Table 1.10). Trout Lake Farm plants produced the second greatest dry root yield in Reidsville in 2013. Native
American Seed plants also produced the most root alkamides in Mills River in 2012, in Reidsville in 2012, and in Reidsville in 2013 with 6.60 mg/g, 5.60 mg/g, and 4.61 mg/g, respectively (Table 1.10).
Precipitation and Daily Temperatures
Precipitation affected field operations, data collection, and plant growth at the Mills River location in 2013. That site received over double the rainfall in 2013 compared to 2012, 222.5 cm compared to 102.1 cm, respectively. In contrast, precipitation at the Reidsville site was similar both years with 138.8 cm in 2012 and 123.5 cm in 2013 (Figure 1.9).
There were also differences in average daily temperatures between the two locations. Although not as dramatic as the precipitation, they might have had an effect on plant growth and flowering. The average daily temperature for the year was slightly less in the southern mountain location of Mills River, 13.6ºC in 2012 and 12.5ºC in 2013 compared to the piedmont location of Reidsville, 15.7ºC in 2012 and 14.2ºC in 2013 (Figure 1.10). Discussion
angustifolia grown in North Carolina. This information will be helpful to the dietary supplement industry as they source raw material from different regions. Predictable yields and consistency in phytochemical composition helps a manufacturer make financial decisions concerning purchasing of raw materials and simplifies the standardization of products.
In this study, location had a large effect on many factors. In addition to the expected environmental differences between a mountain site and a piedmont site, the second year brought rainfall to the mountain site that was more than double that of the first year. This had a negative effect on the production of E. angustifolia in particular. This species is a prairie species with a long tap root that is adapted to drier climates and well-drained soil. E.
purpurea with its large fibrous root system withstood the excessive wet weather much better
than E. angustifolia.
Both E. purpurea and E. angustifolia produced more flowers in Reidsville than in Mills River during the second year of growth. Since Echinacea is primarily a prairie species and prairies tend to be dry, the extensive rains in Mills River during 2013 (Figure 1.9) may have had a detrimental effect on flower production (Foster, 1991; Li, 1998). In our study, seed source had no influence on the number of flowers from two year old E. purpurea plants. Romero et al. (2010) reported similar results for both E. purpurea and E. angustifolia flower production.
Surprisingly, seed source had no effect on E. purpurea dried root yield. This is also beneficial for growers, suggesting that they can purchase seed from a variety of sources and be fairly certain that they will result in good yields. For E. angustifolia, however, there was one seed source, Native American Seed that provided significantly higher dried root yields than any other seed source.
Echinacea angustifolia did not exhibit a clear yield response to location. In general,
dried root yields were higher in the mountains than the piedmont in the first year of growth, but this was complicated by an interaction which was mostly attributable to the extremely high yields for plants from one seed source, Native American Seeds. In year two, not
unexpectedly, dried root yields were significantly higher in the piedmont location than in the mountain location. We attribute that mostly to the extremely wet soil conditions in the mountains for the entire 2013 growing season. A study by Gray et al. (2003) observed an increase in dry root weight in E. purpurea when subjected to controlled drought stress.
Both species of Echinacea grew taller, were more vigorous, reached flowering stage earlier, and produced more overall plant biomass in the second year of growth compared to the first year. This is similar to the data provided by Gray et al. (2003) and Seemannová et al, (2006), who reported a positive correlation between plant age and increased dry root weight in E. purpurea.
acid content in E. purpurea decreased with plant age. In contrast, root cichoric acid content increased with plant age. The increase in root cichoric acid with plant age was also observed by Gray et al. (2003). In our study, the amount of root cichoric acid found in one year old E.
purpurea plants was slightly less than reported by Binns et al. (2002). In contrast to root
cichoric acid, root alkamides decreased from the first year of growth to the second. The decrease in root alkamides with plant age was similar to that reported by Gray et al. (2003). Aside from plant age, root alkamides from both Echinacea species were also produced in greater quantities in plants grown at Reidsville compared to Mills River. These results may be due the soil structure differences between the two locations. A study by Powell et al. (2001) noted a significant increase of root alkamides in plants growing in sandy loam soils compared to those growing in soils with higher clay content.
The accumulation of phytochemicals by location may also be influenced by
differences in soil moisture. Plants grown in Reidsville experienced better soil drainage and less rainfall than those growing in Mills River particularly during the second season of growth. Although an increase in irrigation or rainfall may encourage plant growth, as observed by Yousef et al. (2013), periods of drought stress can increase cichoric acid
concentration in the roots (Gray et al., 2003). Irrigation was observed by Kreft (2005) to have no effect on aerial cichoric acid production in E. purpurea. Rainfall may have also influenced the results by ultimately leading to widespread loss of E. angustifolia across the field in Mills River. Because the taproots of E. angustifolia were subjected to a wet environment for
negative effect of increased precipitation on E. angustifolia roots was also observed by Letchamo et al. (2002).
Some seed sources were very different from the others in growth and yield. For E.
purpurea, in the first growing season, plants grown from seed obtained from Pharmasaat
were short, exhibited poor plant vigor, were slow to mature, had poor yields, and had low foliar and root cichoric acid levels. By the second growing season, however, the plants produced much like those from the other seed sources. Similarly, a study by Romero et al. (2010), found that seed source did influence plant height for first year E. purpurea plants. However, seed source did not have an effect on plant height of two year old plants (Romero et al., 2010). The most striking difference, however, was in the E. angustifolia plants produced from seed from Native American Seed. In both years, these plants were more vigorous, taller, had more flowers, produced higher root yields, and had higher alkamide content than plants from the other seed sources. The physical appearance of the Native American Seed plants also differed from the other five sources in floral ray length and the direction of the ray corolla. The differences were striking enough that we submitted samples for DNA analysis (Authen Technologies, Richmond, CA). The results indicate that the plants contained E. pallida and E. angustifolia genetic material. Further testing is being conducted on these plants that is beyond the scope of this project. Even though these plants looked superior to the other plants growing in the E. angustifolia fields, this is not a positive outcome. The presence of E. pallida genetic material in a product being sold as E.
angustifolia would invalidate that material as being E. angustifolia. It is of interest to us,
Conclusion
In conclusion, this study has demonstrated that when choosing seed for commercial production, seed source, growing location, and plant age are important factors to consider for producing high quality plants. Echinacea purpurea was more consistent across locations and seed sources than E. angustifolia. Plant age is something that needs to be carefully evaluated as biomass yields and phytochemical concentrations do not necessarily increase with plant age. Care should be taken to confirm species identify when growing for the dietary
REFERENCES
Bauer, R. 1998. Echinacea: biological effects and active principles. p. 140-157. In: L.D. Lawson, R. Bauer. (eds.). Phytomedicines of Europe: chemistry and biological activity. American Chemical Society Symposium Series 691. Washington, DC.
Binns, S.E., J.F. Livesey, J.T. Arnason, and B. R. Baum. 2002. Phytochemical variation in
Echinacea from roots and flowerheads of wild and cultivated populations. J. Agric. Food
Chem. 50:3673-3687.
Brovelli, E.A., D. Rua, H. Roh-Schmidt, A. Chandra, E. Lamont, and G.D. Noratto. 2005. Human gene expression as a tool to determine horticultural maturity in a bioactive plant
(Echinacea purpurea L. Moench). J. Agric. Food Chem. 53(21):8156-8161.
Dufault, R.J., G. McCutcheon, B. Ward, B.M. Shepard, J. Rushing, and R. Hassell. 2003. Influence of fertilizer on growth and marker compound of field-grown Echinacea species and feverfew. Scientia Hort. 98(1):61-69.
Flagel, L.E., I. lvarez, J.L. Grafenberg, J.F. Wendel, G.Y. Chung, C.E. Grover, R.A. Rapp, J. Hawkins, and M.P. Widrlechner. 2008. Phylogenetic, morphological, and chemotaxonomic incongruence in the North American endemic genus Echinacea. Amer. J. Bot. 95(6):756– 765.
Foster, S. 1991. Echinacea: nature's immune enchancer. Healing Arts Press, Rochester, VT. Gray, D.E., S.G. Pallardy, H.E. Garrett, and G.E. Rottinghaus. 2003. Acute drought stress and plant age effects on alkamide and phenolic acid content in purple coneflower roots. Planta Med. 69(1):50-55.
Greenfield, J. and J.M. Davis. 2012. Echinacea (Echinacea angustifolia L.). Medicinal Herb Production Guide. N.C. Consortium on Natural Medicines. UNC-Chapel Hill and NC State University.http://www.ces.ncsu.edu/fletcher/programs/herbs/crops/medicinal/E%20angustifo lia%20final%20article%202012.pdf
Kindscher, K. 1989. Ethnobotany of purple coneflower (Echinacea angustifolia, Asteraceae) and other Echinacea species. Econ. Bot. 43(4):498-507.