Trigger factor assisted self assembly of canine parvovirus VP2 protein into virus like particles in Escherichia coli with high immunogenicity
7
0
0
Full text
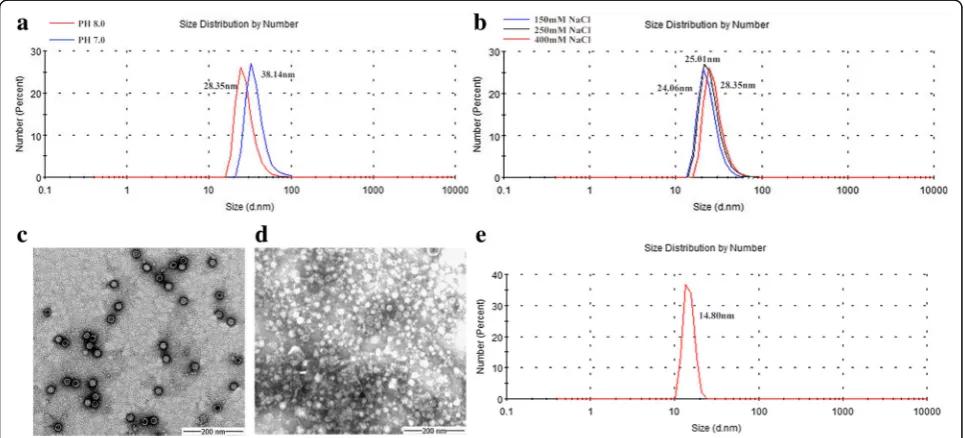
Figure
Related documents
