Copyright 0 1986 by the Genetics Society of America
COMPLEX CIS-ACTING REGULATORS AND LOCUS
STRUCTURE
OF
DROSOPHILA TISSUE-SPECIFIC ADHVARIANTS
L E O N A R D R A B I N O W ' AND W. J. D I C K I N S O N
Department of Biology, University of Utah, Salt Lake City, Utah 841 12
Manuscript received September 13, 1985 Revised copy accepted November 8, 1985
ABS TRA C T
Diverse patterns of tissue-specific expression of alcohol dehydrogenase (ADH) among species of the grimshawi subgroup of Hawaiian picture-winged Drosophila suggests control by complex or multiple, independently acting regulatory ele- ments. T h es e elements act by controlling A d h mRNA accumulation in individual tissue types. Restriction mapping of the A d h loci from these species reveals several insertion/deletion differences, on e of which lies just outside t he 5' en d
of t he structural sequences and correlates with differences in larval patterns of
ADH expression. N o tissue-specific rearrangement of A d h sequences was ob-
served.
NE characteristic of eukaryotic development is the genome-encoded pro-
0
gramming of gene activity in specific tissues and at specific times. Nat- urally occurring variation at genetic loci that determine these patterns of expression, termed temporal loci (GANSCHOW 1975), has been described in a number of experimental systems, including maize, mice and Drosophila (for review, see PAIGEN 1979). Both cis and trans-acting temporal loci have been described for a number of enzyme genes. Cis-acting temporal and other reg- ulators have been found to be closely linked to the affected structural gene whenever the experimental system has been amenable to a detailed genetic analysis (CHOVNICK et al. 1976; DICKINSON 1975; SCHWARTZ 1962; WOODMAN and FREELINC 1981).Many species of the Hawaiian picture-winged Drosophila display dramatic variation in patterns of developmentally programmed tissue and stage-specific expression of a number of enzymes (DICKINSON 1980a). Interspecific hybridi- zation in the laboratory established the existence of both trans- (DICKINSON
1980b) and cis-acting (DICKINSON and CARSON 1979; DICKINSON 1980c) tissue- specific regulatory elements, the properties of which resemble those of the temporal regulatory elzments described within species. Interspecific regulatory variation therefore provides excellent models for the study of developmentally regulated gene function. In addition, studies of this variation may provide clues
524 L. RABINOW AND W. J. DICKINSON
regarding the nature of events that cause changes in gene regulation during evolution.
Alcohol dehydrogenase (ADH) is one of the enzymes demonstrated to be under the control of cis-acting tissue-specific regulators in the Hawaiian Dro- sophila (DICKINSON and CARSON 1979; DICKINSON 1 9 8 0 ~ ) . In a previous report, a recombinant DNA clone of the corresponding locus (Adh) from
D.
melano- gaster was used as a hybridization probe to demonstrate that the accumulation of ADH-specific mRNA is affected by a cis-acting regulatory locus (RABINOW and DICKINSON 1981). T h e species pair in which this study was undertaken,D.
grimshawi andD.
orthofascia, are both members of the grimshawi subgroup of the Hawaiian Drosophila. T h eD.
melanogaster Adh genomic clone has since been used to isolate genomic clones from one of the members of this subgroup,D.
uffinidisjuncta (BRENNAN et al. 1984).We present here several new examples of interspecific variation of ADH tissue-specific expression among four members of the grimshawi species subgroup. Cloned hybridization probes from
D.
afinidisjunctu demonstrate that, in each of the cases examined, the amount of ADH-specific mRNA is affected, although the genomic DNA at Adh is not altered in copy number o r arrangement in any of the tissues examined. T h e existence of discrete regu- latory elements is suggested by the ability of ADH expression patterns to vary independently in different tissues. Finally, analysis of genomic sequences in and around the A d h locus in these species reveals insertion/deletion differences, one of which lies at the 5’ end of the structural gene and correlates with the altered patterns of ADH regulation reported.MATERIALS AND METHODS
Hawaiian Drosophila stocks and culture: The species used in this study (with stock numbers in parentheses) were D. hawaiiensis (J14B8); D. formeEEa (M87G1); D. grimshawi
(Gl); and D. affinidisjuncta (S36G1). The origin and maintenance of these stocks were described previously (DICKINSON 1980a; BRENNAN, ROWAN and DICKINSON 1984).
Tissue preparation, electrophoresis and enzymatic staining for alcohol dehydro- genase: Electrophoresis of ADH in agar noble gels and enzymatic staining were as described elsewhere (DICKINSON and CARSON 1979), with the addition of 0.1 % triton-
X 100 to the staining solution to increase sensitivity approximately two-fold (W. SOFER, personal communication). All tissues for activity analysis, RNA or DNA preparation, were hand dissected in ice-cold Ringer’s solution. To minimize RNA degradation, tis- sues were immediately frozen on dry ice or in liquid nitrogen following dissection. Adult males > I month old were selected for studies of the adult tissue-specific pheno- types reported. Actively feeding third-instar larvae were used for all experiments in- volving larval phenotypes.
Preparation of Drosophila DNA and genomic library construction: DNA was pre- pared from frozen adult Drosophila as described by BINGHAM, LEVIS and RUBIN (1981) by lysis of isolated nuclei, followed directly by cesium chloride density centrifugation. Larval tissue-specific DNA was prepared from hand-dissected organs, as described above, or by the method of COEN, THODAY and DOVER (1982). Genomic libraries were constructed in the lambda cloning vector L-47.1 (LOENEN and BRAMMAR 1980), as described by BRENNAN et al. (1 984). The numbers of recombinant phage resulting were as follows: D. grzmshawz, 7.8 X lo4; D. hawaiiensis, 2.6 X lo5.
CIS-ACTING REGULATORS OF ADH 525
juncta subclone pllB-H14 as the probe (see Figure 6). Hybridization was in aqueous solution at 65", using 10% dextran sulfate. Filters were washed to a final salt concen- tration of 0.1 X SSC at 65". At least six phage showing hybridization were recovered from the D. grimshawi and D. hawaiiensis libraries. Only two from each library were
fully characterized.
Subclones were prepared by insertion of phage fragments produced by digestion with HindIII, Sal1 or both enzymes into the corresponding site(s) on plasmid pBR322. Plas- mid DNA was prepared according to CLEWELL and HELINSKI (1969).
Electrophoresis and hybridization of nucleic acids: DNA was size fractionated in gels of from 0.4 to 1.0% agarose. R N A was size fractionated in 1.0% agarose gels containing formaldehyde according to LEHRACH et al. (1977). RNA was prepared from hand-dissected tissues by phenol-SDS extraction as previously described (RABINOW and
DICKINSON 198 1). Transfers of DNA to nitrocellulose were essentially as described by SOUTHERN (1975), and transfers of R N A to nitrocellulose were as described by GOLD-
BERG (1 980). Nick translation was performed according to RICBY et al. (1977). Hybrid- ization was carried out according to the method of WAHL, STERN and STARK'(^^^^) or as described for the isolation of lambda clones. In situ hybridization to polytene chro- mosomes was carried out as previously described (BRENNAN et al. 1984).
RESULTS
Variation in patterns of ADH tissue-specific expression: It has been shown previously that ADH is expressed in the fat body of both larvae and adults in all species examined (DICKINSON 1980a and unpublished results). Expression of ADH in other tissues is highly variable. There are no clear connections among tissues with respect to ADH expression levels. For example, both
D.
affinidisjuncta andD.
grimshawi express ADH at essentially equal levels in larval carcass and midgut (DICKINSON 1980a). In contrast, althoughD.
hawaiiensisexpresses ADH at levels comparable to the former two species in larval carcass, only a trace is found in the midgut, and this is confined to the anterior midgut. As previously reported (DICKINSON 1980a),
D.
formella lacks detectable levels of the enzyme in both larval carcass and midgut, including gastric ceacae. Electrophoretic gels demonstrating these ADH phenotypes are illustrated in Figure 1. T h e qualitatively distinct patterns of ADH activity in midgut were confirmed by histochemical staining of whole dissected digestive tracts. This procedure also reveals that midgut activity inD.
hawaiiensis is localized in the gastric ceacae (data not shown).Similar examples may be derived from patterns of ADH expression observed in mature adult midguts and hindguts.
D.
affinidisjuncta demonstrates substan- tial ADH expression in midgut but weak expression in hindgut, whereasD.
grimshawi displays an essentially reciprocal pattern, with strong expression found in the hindgut and none found in the midgut (Figure2).
These phe- notypes are confirmed by activity gel electrophoresis. Hindgut and midgut from adult males ofD.
formella show essentially the same pattern of ADH expression as that described inD.
grimshawi, whereasD.
hawaiiensis has little or no activity in either location. Thus, there is no obvious correlation between larval patterns of expression and those found in adults of the same species. A summary of all the phenotypes presented may be found in Table 1.526 L. RARINOW AND W. J. DICKINSON
F I G V R E 1 .--Kcpresent;ttive regulatory differences detected by activity gel electrophoresis. In c.;ich case. dissected parts from five individuals were pooled and homogenized in 15 pl of water,
;itid 5 pl of the extract wits fractionated by electrophoresis and stained for ADH enzymatic activity. I’;uiel A, A D H xtivity i n larval car c~s s (body wall musculature and cuticle). Lanes 1-4: serial five- fold dilutions of D. grimshawi extract: lane 5: undiluted D. formella extract: lanes 6-9: serial five- fold dilutions of D. hawaiiensis extract. In the original, activity is detected in the 25-fold dilution
of D. grimshawi and D. hawaiiensis extracts, but not in undiluted D. formella extract. Panel B. ADH activity in regions of larval gut. Lanes 1-4: D. grimshawi: lanes 5-8: D. hawaiiensis. Within each set. the regions are middle and posterior midgut ( 1 and 5): Malpighian tubules (2 and 6): hindgut (3 and 7): and ;interior midgut (4 and 8).
CIS-ACTING REGULATORS OF ADH 527
GRIM "m"
GRIM ahq
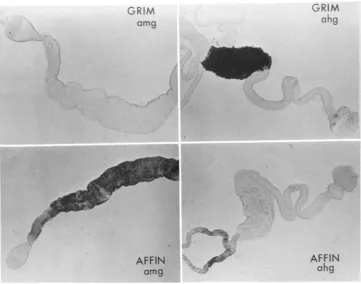
FIGURE 'L.-Representative regulatory differences detected by histochemistry. Whole organs were hand dissected, fixed and stained for A D H activity. Grin) = D. grimshawi; Affin = D. affini- disjuncta: anig = adult midgut; ahg = adult hindgut.
T A B L E 1
Tissuespecific expression of ADH activity
Stage and tissue
Lirval Adulr
Species Ell Ixxly Gill Carcass Midgiit Hindgut
D. affinidisjuncta
++
+
+
+
sf:D. grimshawi
++
+
+
D. formdla
++
+
+
f
-
-
-
-
-
D. hawaiimsis
++
sf:"+
Expression of alcohol dehydrogen;tse in selected larval and adult tissues of four mem- bers of the grimshawi subgroup of Hawaiian Drosophila.
++
= Strong ADH expression;+
= readily detectable expression; f = marginal. or very weak, expression;-
= no detectable expression. See text for more details.a Confined to gastric ceacae.
528 L. RARINOW AND W. J. DICKINSON
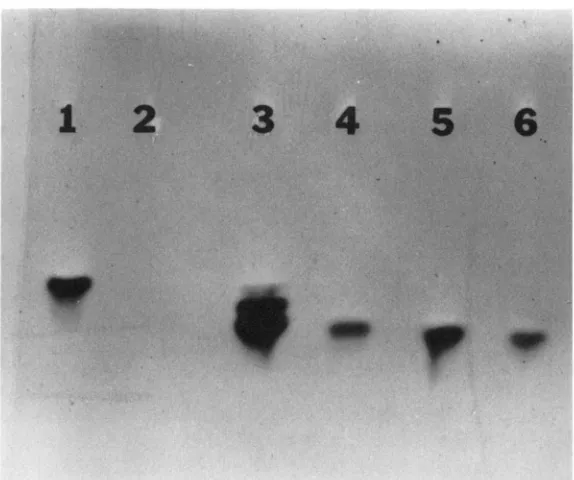
FIGURE 3.--Cis-action o f an element controlling a difference o f the expression o f ADH in D. hawaiiensis and D. formella. T h e following saniples were electrophoresed and stained as for Figure 1. ( I ) D. formella larval fat body, (2) D. formella larval carcass, (3) D. formella X D. hawaiiensis FI
larval fat body, (4) D. formella X D. hawaiiensis FI larval carcass, (5) D. hawaiiensis larval fat body
and (6) D. hawaiiensis larval carcass.
high ADH levels in the male hindgut of some species is related to, but not inseparable from, a sex-specific morphological difference and should not be attributed to a sex-specific difference in ADH expression per se.
Genetic control of tissue-specific ADH expression in interspecific hybrids:
Cis- or trans-action of a regulatory difference can be distinguished if there are suitable markers for the products of alternative structural loci, such as electro- phoretic mobility differences. Among the four stocks considered here, only
D.
hawaiiensis has ADH differentiable from that in other species by activity gel electrophoresis. In
D.
hawaiiensis XD.
formella hybrids, only theD.
hawaiiensis form of ADH is present in carcass, even though both are detected in fat body (Figure 3). Thus, parental differences are preserved in hybrids, indicating con- trol of this regulatory difference by a cis-acting element. Examination of the ADH present in larval carcass from hybrids ofD.
formella andD.
grimshawi yields results consistent with cis-acting regulation of tissue-specific expression of ADH in larval midgut and carcass,i.
e., levels of expression are intermediate in these tissues of hybrids.CIS-ACTING REGULATORS OF ADH 529
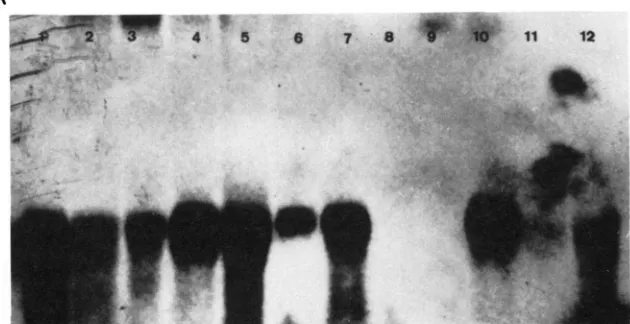
FIGURE 4.-Tissue-specific RNA. '1.otaI R N A froin e;rc.li tissue was electrophoresed, blotted to nitrocellulose and probed with D. affinidisjunrfa subclone 1 I R-S8. Panel A, Larval tissue-specific RNAs derived from four species and three tissues. Lanes 1-3: D. afinidisjuncta; lanes 4-6: D. grimshawi; lanes 7-9: D. formella; lanes 10- 12: D. hawaiiensis. Tissues represented from each species itre fat body (1, 4, 7 and 10); gut (2, 5 . 8 and I I); and carcass (3, 6, 9 and 12). Panel B, Adult tissue-specific RNAs. l an es 1 and 2: D. affinidisjuncta; lanes 3 and 4: D. grimshawi. Hindgut-specific RNA is shown in lanes 1 and 3; midgut-specific RNA is in lanes 2 and 4.
530 L. RABINOW AND W. J. DICKINSON
mologous probe from
D.
afjnidisjuncta also allowed detection of trace amounts of ADH inRNA in the guts of D. hawaiiensis, where enzymatic activity is restricted to the gastric ceacae and is barely detected. This quantitative differ- ence in enzyme activity is thus reflected at the RNA level.Distribution of ADH-specific sequences in RNA prepared from hindguts and midguts of
D.
afjnidisjuncta andD.
grimshawi adult males > I month old is shown in Figure 4B. Tissue-specific expression of ADH is again paralleled by the presence o r absence of the mRNA. An additional example of quantitative control of ADH activity and mRNA is found in D. afinidisjuncta adult male hindgut, which demonstrates a low but reproducible level of both.Chromosomal location: As a first step toward identifying structural corre- lates of altered regulation, w e examined the possibility that gross chromosomal rearrangements have brought the A d h locus into a new context. I n situ hy- bridization procedures were used to visualize the A d h locus on polytene chro- mosomes of the four species considered here, as well as species representing all other major subgroups of picture-winged Drosophila. In every case, hybrid- ization is confined to the same position relative to clearly homologous banding patterns. I t is clear that altered regulation is not a consequence of gross rear- rangement. D. afinidisjuncta,
D.
grimshawi and D. formella are homosequentialin banding pattern throughout chromosome I l l , on which A d h is located, and there are no visible structural alterations in the bands around the A d h gene (Figure 5). Even when an inversion has moved the A d h locus away from the base of chromosome ZZZ, as in
D.
hawaiiensis, the pattern of bands immediately around the locus is unchanged (see Figure 5).Genomic Adh sequences of D. afinidisjuncta, grimshawi, formella and ha- waiiensis: Restriction maps of the A d h loci from the four species are compared i n Figure 6 . These maps are based on data obtained from genomic Southern transfers probed with
D.
afinidisjuncta subclones and on isolated recombinant phage from D. afjnidisjuncta, grimshawi and hawaiiensis.A high degree of sequence conservation at the A d h locus among the four species is indicated by the extensive cross hybridization, as well as by the colinearity and spacing of restriction sites. Although several small fragment- length differences and apparent point changes resulting in gain or loss of some restriction sites are present, no ambiguities in the relationships of comparable restriction sites are found. Repetitive elements localized to the 3’ end of the
D.
afjnidisjuncta A d h locus are present in roughly equal numbers in the ge- nomes of the other species (not shown).Adh structural sequences are almost entirely contained on a 1.65-kb EcoRI fragment that is conserved in all four species (Figure 7A). Only two insertion/ deletion differences greater than approximately 100 base pairs (bp) were found
i n mapping of between 10 and 20 kb from each of the species. Both of these differences result in fragments shorter than the corresponding
D.
afinidisjunctafragments. One difference lies at the 3’ end of the A d h gene and results in a Hind111 fragment of 1.9 kb in
D.
afinidisjuncta, formella and hawaiiensis being shortened to 1.67 kb inD.
grimshawi. T h e other difference is found at the 5’CIS-ACTING REGULATORS OF ADH 53 1
FIGURE 5.--Chromoson:al context of the Adh gene. Top: In situ hybridization of plasniid 32RE1 (HRENNAN ef al. 1984) to the third chronlosonie of D. grimshawi. Below: aceto-orcein stained cliromosoiiies of D. grimshawi ( G ) , D. formella (F) and D. hawaiiensis (H). In each case, the arrow indicates the band in or near which the Adh gene is located. Lines connect homologus bands useful ;IS landmiirks. In D. grimshawi and D. formella, which both have the “standard” sequence of
kinds on chronlosonie / I / . the heavy bands at the right are adjacent to centric heterochromatin. I n D. hawaiiensis, inversion Sg (CARSON and STALKER 1968), which has one breakpoint in the centric lieterochrom;itin, has moved the corresponding bands (and the nearby Adh locus) to a position near the center of chromosonie I / / .
formella) or a 180-bp difference
(D.
hawaiiensis), relative to eitherD.
grimshawi o r affinidisjuncta (Figure 7B).DNA of the Adh locus from specific tissues: To examine the possibility of tissue-specific alteration of Adh genomic sequences, DNA was prepared from 100 hand-dissected larval fat bodies, midguts and carcasses of
D.
grimshawi,D.
formella andD.
hawaiiensis. Aliquots were digested with either Hind111 or532 L. RABINOW AND W. J. DICKINSON
D aftinidispncta
I l E - S H l 3
I I t € - S E I
t
-
I l l - H I 4-
ID hawaiiensis
I +
E
..
. . . .
i
I I I I I I 1 I It
-6 -4 -2 -0 + +2 +4
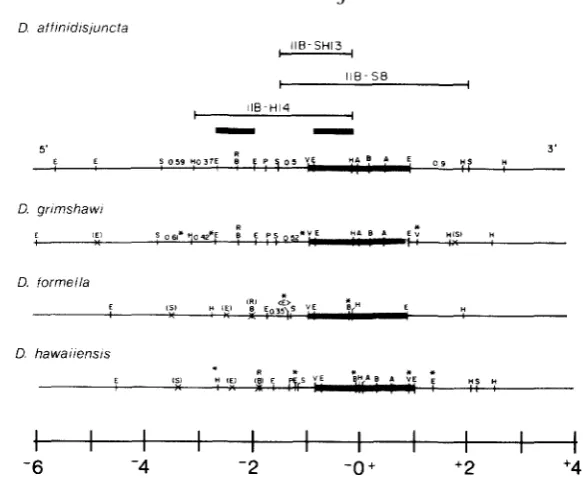
FIGURE 6.--Coniparison of Adh genomic regions by restriction mapping. T h e approximate ex- tent of the transcribed region in indicated by cross-hatching. D. affinidisjuncta subclones (thin lines) and gel-isolated fragments (heavy bars) used as hybridization probes are indicated. T h e maps are aligned at the Hind111 site arbitrarily designated 0.0. Restriction enzyme recognition sites are A = AuaI; B = BglI; E = BcoRI; H = HindIII; P = PstI; R = EcoRV; S = Sall; V = PuuII. An (x) indicates a missing restriction site, and an asterisk (*) indicates a new site or some of the different fragment lengths, all relative to D. afinidisjuncta. Because cloned sequences were not available from D. formella and the presence of repetitive elements near the 3' end of the gene in all species complicated mapping by hybridization to Southern transfers of genomic restriction fragments, some restriction sites present in other species were not mapped in D. formella. Hence, absence of a site on this map does not necessarily mean that the site is missing. T h e ( E ) indicates a possible polymorphism i n D. formella.
have been detected. Additionally, changes in the context of the Adh gene among these tissues would be revealed if any restriction fragments detected by this probe were affected. Only the band(s) expected from mapping of isolated phage o r from adult genomic Southern transfers were observed, thus effec- tively ruling out rearrangement of the gene within 3 kb 5' to the structural gene in these tissues (not shown). T h e intensity of detected bands from differ- ent tissue samples was equal, indicating no gross under o r over replication of
Adh genomic sequences on a tissue-specific basis.
DISCUSSION
T h e new regulatory patterns reported here, when considered with previously reported examples (DICKINSON 1980a, 1980b, 1983; RABINOW and DICKINSON
198 l ) , show that evolutionary changes in patterns of gene expression can affect different tissues and stages independently. This suggests the existence of mul- tiple and/or complex regulatory sites that can be altered independently or in
CISACTING REGULATORS OF ADH 533
2.8
1.01 1.00 0.61
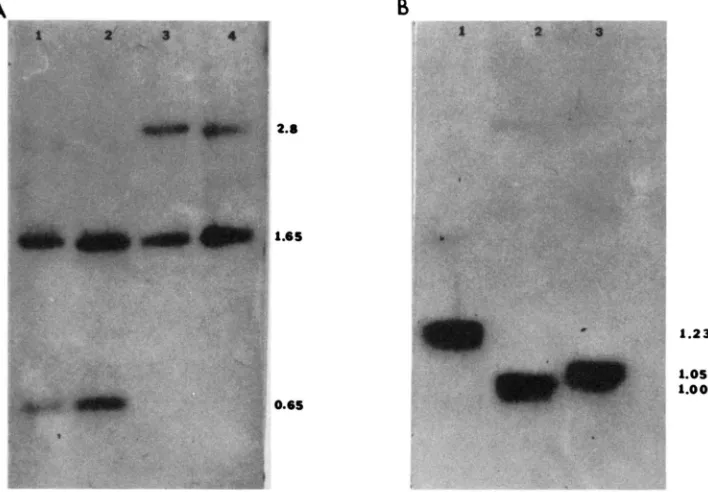
FIGURE 7.-lnterspecific comparisons of restriction fragment lengths. Genomic DNA digested as specified below was electrophoresed, blotted and hybridized to cloned sequences from D. a& idisjuncta as indicated. Panel A. EcoRl digested DNA from D. afinidljuncta (lane l), D. gtimshawi
(lane 2), D. formella (lane 3) and D. hawaiiensl (lane 4) hybridized to t w o gel-purified fragments from 11B-HI4 (Figure 6). T h e first fragment extends from the Hind111 site (0.0) near the middle of the coding region to the EcoRl site (-0.65) near the 5’ end and detects the conserved 1.65-kb fragment that carries most of the coding sequences in all species. T h e second probe is an EcoRl fragment extending from -1.65 to -2.3 in D. affinidljuncta. I t detects the corresponding 0.65- kb fragment in that species and in D. grimshawi, but detects a 2.8-kb fragment in the other species, documenting the absence of the -2.3 site (Figure 6). Panel B HindlII, Sal1 doubledigested DNA from D. grimsham. (lane 1). D. formella (lane 2 ) and D. hawaiiensl (lane 3 ) hybridized to 11B- SH13 (Figure 6). This documents length differences from the central Hindlll site to the Sal1 site 5’ to the coding region. Conservation of the 1.65-kb EcoRI fragment (above) confines the observed length differences to the EcoRISalI fragment adjacent to the 5‘ end of the coding region. T h e length of this fragment in D. afFnidisjuncta (not shown) is almost identical to that in D. grimshawi.
T h e apparent separation of cis-acting sites responsible for expression of Dro- sophila yolk proteins in t w o different tissues recently reported by GARABEDIAN, HUNG and WENSINK (1 985) supports a similar conclusion.
T h e one case for which an appropriate test was possible clearly showed cis-
acting control of the regulatory difference. All other cases gave a result (in- termediate expression) consistent with cis-acting control. We believe that the multiple o r complex regulatory sites implicated by the diversity of regulatory patterns are most likely to be located in a cis-acting regulatory region adjacent to the structural gene. However, w e cannot exclude the formal possibility that multiple truns-acting regulators interact with a single cis-acting site in a species- specific or allele-specific manner (i.e., pattern differences are due to different truns-acting regulators, but they appear as cis-acting controls in hybrids, be-
U
534 L. RABINOW AND W. J. DICKINSON
cause each species’ trans-acting factors interact only with a cis-acting component from the same species).
Results at the RNA level extend the conclusion that the regulatory sites under consideration act at or near transcription to several new cases. T h e low but distinctly detectable presence of both ADH enzyme activity and mRNA in anterior midgut of
D.
hawaiiensis larvae and hindgut ofD.
affinidisjuncta adult males indicates that similar mechanisms apply to quantitative as well as quali- tative (on/off) regulation. We find no evidence for loss, amplification or rear- rangement of the Adh locus in any of the tissues affected by regulatory variants. We also find no evidence that differences between expression patterns result from gross chromosomal rearrangements.Analysis of the structural and flanking sequences of the Adh locus in D.
affinidisjuncta, grimshawi, formella and hawaiiensis reveals a high degree of co- linearity at the level of restriction sites and hybridization to subclones. Com- parison of the A d h loci of the four species does not define sequences respon- sible for the tissue-specific control of Adh expression, but certain regions are circumstantially implicated and bear further study.
D. afinidisjuncta and
D.
grimshawi express ADH reciprocally in adult midguts and hindguts. Comparison of the Adh locus restriction maps of these two species reveals no major differences between them at the 5’ end or within the structural gene. There is a relatively large insertion/deletion difference at the 3’ end (a Hind111 fragment of 1.9 kb inD.
afinidisjuncta, compared to a 1.67- kb fragment in D. grimshawi), but there is no obvious correlation with pheno- type, becauseD.
formella andD.
hawaiiensis, which resemble D. grimshawi in lacking ADH in adult midguts, have the 1.9-kb restriction fragment.Correlation between Adh restriction maps and patterns of expression is found with respect to larval phenotypes. Comparison of the A d h restriction maps of either
D.
formella orD.
hawaiiensis with those ofD.
affinidisjuncta or grimshawireveals a large insertion/deletion difference at the 5’ end of the structural gene. Recall that
D.
affinidisjuncta andD.
grimshawi are identical in their larval patterns of ADH expression, whereasD.
hawaiiensis expresses low levels of ADH in the larval gastric ceacae, shows none in midgut and shows levels comparable to the former two species in larval carcass. T h e restriction map of theD.
hawaiiensis A d h locus reveals a deletion relative to the former two species of approximately 180 bp, immediately 5’ to the A d h coding sequences.D.
formella lacks all ADH expression in larval gut and carcass, and it reveals a 230-bp deletion relative to
D.
affinidisjuncta orD.
grimshawi at the 5‘ end ofthe Adh coding sequences. T h e organization of the
D.
affinidisjuncta Adh gene is similar to that seen inD.
melanogaster (BENYAJATI et al. 1983), including transcription from two promoters (BRENNAN et al. 1984; R . ROWAN and W. J. DICKINSON, unpublished results). In both cases, the proximal promoter is used primarily in larvae, and the distal promoter is used primarily in adults. T h e length differences inD.
formella andD.
hawaiiensis relative toD.
afjnidisjunctaCISACTING REGULATORS OF ADH 535
both
D.
formella andD.
hawaiiensis. T h e additional 50 bp in the hawaiiensis Adh allele relative to D. formella might then contain information required for ADH expression in larval carcass and gastric ceacae. This is consistent with the presence of multiple, cis-acting elements tightly linked to the structural gene, as suggested above.One further correlation can be made concerning levels of ADH in larvae of D. formella relative to
D.
grimshawi. We noted previously that D. formella larval fat body expresses only about one-fourth the ADH found in the same tissue of D. grimshawi (RABINOW and DICKINSON 1981). It is interesting thatD.
for- mella bears the larger deletion relative to D. grimshawi o r aflanidisjuncta. There- fore, information determining levels of transcription may also be contained in the fragment missing in D. formella, o r such information may be inseparable from that required for expression in multiple tissues.These hypotheses are, of course, tentative. Indeed, other studies of Droso- phila Adh locus structure, using strains of D. melanogaster as well as those of several other species, have described DNA polymorphisms without attendant dramatic effects on ADH expression (LANGLEY, MONTGOMERY and QUATTLE- BAUM 1982; BODMER and ASHBURNER 1984; KREITMAN 1 9 8 3 ) . However, tis- sue- and stage-specific ADH levels were not examined in these studies; thus, the variation in expression patterns described in this report would probably have gone undetected.
Although correlations between ADH expression patterns and Adh locus re- striction maps of the grimshawi subgroup variants provide preliminary and tentative evidence concerning the location of tissue-specific regulatory infor- mation, unambiguous results await a direct functional demonstration that can only be provided by DNA-mediated transformation.
D.
melanogaster P-element germline transformation is effective in the Hawaiian Drosophila, although noAdh transformants have so far been recovered (BRENNAN, ROWAN and DICK- INSON 1 9 8 4 , and unpublished results). Transient expression assays following injection of DNA into embryos as described by W. SOFER (unpublished results) have, however, recently begun to provide encouraging results (P. THORPE, unpublished results).
This work was supported by National Institutes of Health grant HD10723 to W.J.D. and National Institutes of Health Predoctoral Fellowship GM07464 to L.J.R. PAULA FEY provided technical assistance in some of these experiments. We would also like to acknowledge the critical comments and continual encouragement of RAY GESTELAND.
LITERATURE CITED
BENTON, W. D. and R . W. DAVIS, 1977 Screening lambda gt recombinant clones by hybridization to single plaques in situ. Science 196: 180-182.
T h e messenger RNA for alcohol dehydrogenase in Drosophila melanogaster differs in its 5’ end in different develop- mental stages. Cell 33: 125-133.
Cloning o f DNA sequences from the white BENYAJATI, C., N. SPOEREL, H . HAYMERIE and M. ASHBURNER, 1983
BINGHAM, P. M . , R. LEVIS and G. M . RUBIN, 1981
536
BODMER, M. and M. ASHBURNER, 1984
L. RABINOW AND W. J. DICKINSON
Conservation and change in the DNA sequences coding
Introduction of a functional P element for alcohol dehydrogenase in sibling species of Drosophila. Nature 3 0 9 425-430.
into the germline of Drosophila hawaiiensis. Cell 38: 147-151.
Isolation and initial charac- terization of the alcohol dehydrogenase gene from Drosophila aflnidisjuncta. J. Mol. Appl. Genet. 2: 436-446.
BRENNAN, M., R. ROWAN and W. J. DICKINSON, 1984
BRENNAN, M., R. ROWAN, L. RABINOW and W. J. DICKINSON, 1984
CARSON, H. L. and H. D. STALKER, 1968 Polytene chromosome relationships in Hawaiian species of Drosophila.1. T h e D. grzmshawi group. Stud. Genet. (University of Texas Publication 6818): 335-354.
CHOVNICK, A., W. GELBERT, M. MCCARRON, B. OSMOND, E. P. M. CANDIDO and D. L. BAILLIE, 1976 Organization of the rosy locus in Drosophila melanogaster: evidence for a control ele- ment adjacent to the xanthine dehydrogenase structural element. Genetics 84: 232-255. CLEWELL, D. B. and D. R. HELINSKI, 1969 Supercoiled circular DNA-protein complex in E. Coli:
purification and induced conversion to an open circular DNA form. Proc. Natl. Acad. Sci. USA 62: 1159-1163.
Rate of turnover of structural variants in the COEN, E. S., J. J. THODAY and G. DOVER, 1982
DICKINSON, W. J., 1975
DICKINSON, W. J., 1980a
DICKINSON, W. J., I980b
rDNA gene family of Drosophila melanogaster. Nature 295: 564-568.
A genetic locus affecting the developmental expression of an enzyme in
Evolution of patterns of gene expression in Hawaiian picture-winged
Complex cis-acting regulatory genes demonstrated in Drosophila hybrids.
Tissue specificity of enzyme expression regulated by diffusible factors: evidence from Drosophzla hybrids. Science 207: 995-997.
Tissue-specific allelic isozyme patterns and cis-acting developmental reg- ulators. Isozymes 9: 107-1 22.
Regulation of the tissue specificity of an enzyme by a czs-acting genetic element: evidence from Drosophila hybrids. Proc. Natl. Acad. Sci. USA 76:
Simultaneous genetic control of the structure and rate of synthesis of murine glucuronidase. pp. 633-647. In: Isozymes: Genetics and Evolution, Edited by C. L. MARKERT. Academic Press, New York.
Independent control elements that determine yolk protein gene expression in alternative Drosophila tissues. Proc. Natl. Acad. Sci. USA 82: 1396-1400.
Isolation and partial characterization of the Drosophila alcohol dehydrogen- ase gene. Proc. Natl. Acad. Sci. USA 77: 5794-5798.
Nucleotide polymorphism at the alcohol dehydrogenase locus of Drosophila melanogaster. Nature 304: 4 12-4 17.
Restriction map variation in the Adh region of Drosophila. Proc. Natl. Acad. Sci. USA 7 9 5631-5635.
RNA molecular weight determination by gel electrophoresis under denaturing conditions: critical reexamination. Bio- chemistry 16: 4743-475 1 .
A bacteriophage lambda vector for cloning large DNA fragments made with several restriction enzymes. Gene 10: 249-259.
Drosophila melanogaster. Dev. Biol. 47: 206-2 14.
Drosophila. J. Mol. Evol. 16: 73-94.
Dev. Genet. 1: 229-240. DICKINSON, W. J., 1980c
DICKINSON, W. J., 1983
DICKINSON, W. J. and H. L. CARSON, 1979
4559-4562.
GANSCHOW, R. E., 1975
GARABEDIAN, M. J., M-C. HUNG and P. C. WENSINK, 1985
GOLDRERG, D., 1980
KREITMAN, M., 1983
LANGLEY, C. H., E. MONTGOMERY and W. F. QUATTLEBAUM, 1982
LEHRACH, H., D. DIAMOND, J. M. WOZNEY and H. BOEDTKER, 1977
CIS-ACTING REGULATORS OF ADH
537
PAIGEN, K., 1979 Genetic factors in developmental regulation. In: Physiological Genetics, Edited by J. G. SCANDALIOS. Academic Press, New York.
A cis-acting regulator of enzyme tissue specificity in
Drosophila is expressed at the RNA level. Mol. Gen. Genet. 183: 264-269.
Labeling DNA to high specific activity in vitro by nick translation with DNA polymerase I. J. Mol. Biol. 113: 239-251.
Genetic studies on mutant enzymes in maize: control of gene action in synthesis of p H 7.5 esterase. Genetics 47: 1609-1615.
Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98: 503-518.
Efficient transfer of large DNA fragments from agarose gels to DBM paper and rapid hybridization using dextran sulfate. Proc. Natl. Acad. Sci. USA 7 6 3683-3687.
Identification of a genetic element that controls the RABINOW, L. and W. J. DICKINSON, 1981
RICRY, P. W. J., M. DIECKMANN, C. RHODES and P. BERG, 1977
SCHWARTZ, D., 1962
SOUTHERN, E. M., 1975
WAHL, G. M . , M. STERN and G . P. STARK, 1979
WOODMAN, J. C. and M. FREELING, 1981
organ-specific expression of ADH 1 in maize. Genetics 98: 357-378.