Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Species Identification and Strain Differentiation of Dermatophyte
Fungi by Analysis of Ribosomal-DNA
Intergenic Spacer Regions
COLIN J. JACKSON, RICHARD C. BARTON,*ANDE. GLYN V. EVANS
Department of Microbiology and PHLS Mycology Reference Laboratory, University of Leeds and General Infirmary, Leeds LS2 9JT, United Kingdom
Received 14 October 1998/Returned for modification 11 November 1998/Accepted 13 January 1999
Restriction fragment length polymorphisms (RFLPs) identified in the ribosomal-DNA (rDNA) repeat were used for molecular strain differentiation of the dermatophyte fungusTrichophyton rubrum. The polymorphisms were detected by hybridization ofEcoRI-digestedT. rubrumgenomic DNAs with a probe amplified from the small-subunit (18S) rDNA and adjacent internal transcribed spacer (ITS) regions. The rDNA RFLPs mapped to the nontranscribed spacer (NTS) region of the rDNA repeat and appeared similar to those caused by short repetitive sequences in the intergenic spacers of other fungi. Fourteen individual RFLP patterns (DNA types A to N) were recognized among 50 random clinical isolates ofT. rubrum. A majority of strains (19 of 50 [38%]) were characterized by one RFLP pattern (DNA type A), and four types (DNA types A to D) accounted for 78% (39 of 50) of all strains. The remaining types (DNA types E to N) were represented by one or two isolates only. A rapid and simple method was also developed for molecular species identification of dermatophyte fungi. The contiguous ITS and 5.8S rDNA regions were amplified from 17 common dermatophyte species by using the universal primers ITS 1 and ITS 4. Digestion of the amplified ITS products with the restriction endonuclease
MvaI produced unique and easily identifiable fragment patterns for a majority of species. However, some closely related taxon pairs, such asT. rubrum-T. soudanense andT. quinkeanum-T. schoenlenii could not be distinguished. We conclude that RFLP analysis of the NTS and ITS intergenic regions of the rDNA repeat is a valuable technique both for molecular strain differentiation of T. rubrumand for species identification of common dermatophyte fungi.
Dermatophyte fungi cause a variety of superficial and usual-ly easiusual-ly treated mycoses. However, nail infections (onychomy-coses) due toTrichophyton rubrumare often more intractable, and relapse frequently occurs following cessation of antifungal therapy. Drug resistance is not a primary factor in such epi-sodes, as susceptibility testing of nail isolates pre- and post-therapy usually confirms the strains to be fully sensitive to the chemotherapeutic agent used. We are currently attempting to establish whether recurrence ofT. rubrumonychomycosis fol-lowing an appropriate course of treatment is due primarily to treatment failure or to reinfection with a new strain. This re-quires the development and evaluation of an effective method for strain differentiation inT. rubrum.
Conventional (phenotypic) strain typing of dermatophyte fun-gi is problematic due to a lack of stable characteristics distin-guishing between isolates. MostT. rubrumstrains show unifor-mity in both microscopical and colonial appearance, although variations in colony morphology do exist. However, these ap-parent strain differences are often not stable on subculture or may simply be artifacts due to specific growth conditions or the presence of contaminating bacteria (21). Alternative molecular (genotypic) approaches to the subtyping of dermatophyte fungi have met with limited success. The discrimination achieved by techniques such as arbitrarily primed PCR (AP-PCR) (7, 11), random amplified polymorphic DNA analysis (RAPD) (16, 27), and restriction analysis of mtDNA (15) is generally ade-quate for species identification but insufficiently sensitive for
strain differentiation ofT. rubrum. Zhong et al. examined thirty isolates ofT. rubrumby RAPD and found 22 strains to be indis-tinguishable and 8 to show very minor differences (27), while Liu et al., using AP-PCR, reported no differences between 8 strains ofT. rubrum(11).
Interstrain polymorphisms in the spacer regions of fungal ribosomal-DNA (rDNA) repeat units have provided practical epidemiological markers for typing a range of clinically impor-tant species. Recently, fragment length polymorphisms present in the rDNA nontranscribed spacer (NTS) regions have been used to type bothCandida krusei(4) andAspergillus fumigatus (19), and nucleotide sequence variations in the internal tran-scribed spacers (ITS I and II) have been shown to differentiate strains of Pneumocystis carinii f. sp. hominis (10). We have examined molecular variation in the rDNA repeats of T. rubrumand other dermatophyte fungi and identified length variations in the NTS region which have been used for strain differentiation.
Additional analysis of the ITS regions has provided a simple and reproducible molecular method for dermatophyte species characterization, utilizingMvaI restriction enzyme patterns of PCR-amplified ITS I and ITS II regions. In this report we show that polymorphism analysis of the rDNA intergenic regions is a valuable technique both for strain typing and species identi-fication in this important group of pathogenic fungi.
MATERIALS AND METHODS
Dermatophyte isolates.Clinical isolates ofT. rubrumand other dermatophyte species were cultured from skin, hair, and nail samples submitted to the Leeds PHLS Mycology Reference Laboratory by general practitioners and hospital dermatology departments in the United Kingdom. Isolates from Iceland, Fin-land, HolFin-land, and Germany were received during the course of a clinical trial from patients with onychomycosis in these countries. Cultures of six
dermato-* Corresponding author. Mailing address: PHLS Mycology Refer-ence Laboratory, Department of Microbiology, The Old Medical School, Thoresby Place, University of Leeds, Leeds LS2 9JT, England. Phone: 44 113 233 5598. Fax: 44 113 233 5640. E-mail: [email protected].
931
on May 15, 2020 by guest
http://jcm.asm.org/
phyte species were provided by Gillian Midgley, Institute of Dermatology, St. Thomas’ Hospital, London, United Kingdom, and three type cultures were obtained from the National Collection of Pathogenic Fungi, PHLS Mycology Reference Laboratory, Bristol, United Kingdom.
All clinical isolates were identified to species level on the basis of standard biochemical tests, microscopy, and colony characteristics. Strains grown from clinical samples were subcultured once to confirm purity, and cultures were maintained in sterile water and on Sabouraud agar slopes.
Isolation of fungal DNA.Strains were cultured in 100 ml of Sabouraud liquid medium (Oxoid; Unipath Ltd., Basingstoke, United Kingdom) and incubated with shaking for up to 7 days at 27°C. Hyphal growth was harvested by filtration and washed twice with 100 ml of sterile saline. Strains which could not be
processed immediately were frozen at280°C prior to extraction. Liquid nitrogen
was added to 2 to 3 g of frozen hyphae in a prechilled mortar, and the cells were ground finely with a pestle. Approximately 200 mg of frozen, ground mycelium
was placed in a 1.5-ml microcentrifuge tube, and 600ml of lysis buffer (400 mM
Tris-HCl, pH 8.0; 60 mM EDTA; 150 mM NaCl; 1% sodium dodecyl sulfate [SDS]; 40 mg/ml proteinase K) was added. Samples were incubated for 1 h at
60°C with occasional mixing, and then 100ml of 5 M sodium perchlorate was
added and incubation continued for a further 15 min at 60°C. Tubes were cooled
on ice, and extraction was performed with 500ml of ice-cold chloroform and then
with equal volumes of phenol-chloroform-isoamyl alcohol (25:24:1; pH 8.0; BDH Lab Supplies, Poole, United Kingdom) and finally with chloroform. Purified nucleic acids were precipitated with 2 volumes of ice-cold 95% ethanol, washed
twice in 500ml of 70% ethanol, air dried, and resuspended in 150 to 200ml of
sterile water.
Detection of rDNA polymorphisms inT. rubrum.Ten micrograms (;10ml) of each DNA sample was digested for 18 h with 15 U of restriction endonuclease
EcoRI (MBI Fermentas, IGI Ltd., Sunderland, United Kingdom) in a total
vol-ume of 20ml. Samples were electrophoresed in 0.8% agarose gels, stained with
ethidium bromide, and photographed. Gel denaturation and neutralization and immobilization of DNA fragments onto nylon membranes (Hybond-N; Amer-sham International plc, Little Chalfont, United Kingdom) by Southern transfer were performed according to standard protocols.
An rDNA probe was amplified from template DNA ofT. rubrumNCPF 295
by using universal primers NS 5 (59AACTTAAAGGAATTGACGGAAG 39)
and ITS 4 (59TCCTCCGCTTATTGATATGC 39) (26). The 1,219-bp probe
consisted of a 550-bp fragment from the 39end of the 18S rDNA plus the
ad-jacent ITS I, 5.8S rDNA, and ITS II regions (Fig. 1). The probe was labelled by incorporating digoxigenin (DIG)-labelled dUTP (Boehringer-Mannheim UK Ltd., Lewes, United Kingdom) into a standard PCR mixture. The PCR mixture contained reaction buffer (50 mM KCl, 10 mM Tris-HCl [pH 9.0], 0.1% Triton X-100), 1.5 mM magnesium chloride, a DIG-deoxynucleoside triphosphate (dNTP) mix (0.2 mM each of dATP, dCTP, dGTP; 0.13 mM dTTP; 0.07 mM DIG-11-dUTP, alkali labile, pH 7.0), 30 pmol each of primers NS 5 and ITS 4
(MWG-Biotech, Milton Keynes, United Kingdom), 5 U of Taqpolymerase
(Stratech Scientific Ltd., Luton, United Kingdom), and approximately 10 ng of
diluted template DNA, made up to a total volume of 100ml with pure water.
PCR cycling conditions were 35 cycles of 95°C for 1 min, 55°C for 1 min, and 72°C for 2 min, followed by an extension step of 72°C for 10 min. The probe was gel purified with a GX resin system (Nucleon Biosciences, Coatbridge, United
Kingdom), and the probe concentration was adjusted to;20 ng/ml in a 15-ml
volume of hybridization solution (53SSC [13SSC is 0.15 M NaCl plus 0.015 M
sodium citrate], 0.1%N-lauroylsarcosine, 0.02% SDS, 1% blocking reagent
[for-mulation unknown; Boehringer-Mannheim]). Hybridization was carried out for
18 h at 65°C, followed by four stringent washes (twice for 5 min at 25°C with 23
SSC–0.1% SDS and twice for 15 min at 65°C with 0.53SSC–0.1% SDS). Bound,
labelled probe was conjugated with anti-DIG-AP Fab fragments (Boehringer-Mannheim), and signal was developed by using the chemiluminescent substrate CSPD (Boehringer-Mannheim).
Detection ofMvaI restriction site polymorphisms in the ITS I and II regions of the rDNA.The contiguous ITS I, 5.8S, and ITS II regions were amplified from
a range of dermatophyte species by using the conserved primers ITS 1 (59TC
CGTAGGTGAACCTGCGG 39) and ITS 4 (26). The PCR mixture and
ampli-fication conditions were the same as those used for preparing the labelled probe, but the DIG-dNTP labelling mix was replaced with a standard mix producing a final concentration of 0.2 mM of each dNTP. Species-characteristic restriction site polymorphisms were detected by digestion of the amplified product with the
restriction enzymeMvaI (MBI Fermentas), which recognizes the sequence 59
CC(T/A)GG 39. Digest fragments were separated by electrophoresis in 2%
agarose gels and stained with ethidium bromide prior to photography.
RESULTS
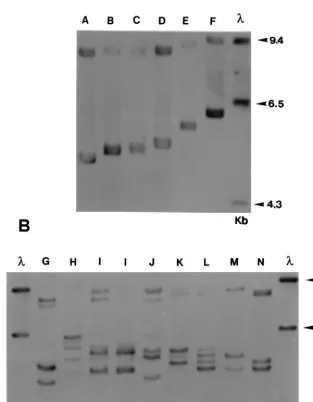
Strain typing ofT. rubrum.From the results of the probe hy-bridizations and using sequence data of theT. rubrum18S and ITS I and ITS II regions (1, 2), a provisional restriction map of the rDNA repeat was constructed (Fig. 1). AnEcoRI fragment comprising ITS I, all of the 18S gene, and an indeterminate length of the NTS region was polymorphic in size between strains. In most isolates, the length of this fragment varied over the range 4.7 to 5.8 kb (Fig. 2A), although a smaller proportion of strains had either two or four polymorphic bands in the same region (Fig. 2B). A second lower-molecular-weightEcoRI frag-ment of approximately 3 kb (not shown) was invariant in size and represented an uncharacterized segment of the 25S gene and the ITS II region proximal to this (Fig. 1). The probe also detected a third, high-molecular-weight band which was ap-proximately equal in length to the sum of the sizes of the first two fragments. This may represent a subpopulation of repeat units from which one of the twoEcoRI sites has been lost.
Six different single band types (types A to F [Fig. 2A]) and eight multiple band patterns (types G to N [Fig. 2B]) were rec-ognized, making a total of 14 patterns overall among the 50 isolates examined. One pattern, DNA type A, was found for 19 of 50 (38%) of strains and was present in isolates from all geographic locations. Three other pattern types were preva-lent: DNA types B (9 of 50 strains; 18%), C (5 of 50 strains; 10%), and D (6 of 50 strains; 12%). All other types were rep-resented by single isolates only, except for two strains of DNA type I (Table 1; Fig. 2B). There was no obvious correlation between any pattern type and the geographical or clinical ori-gin of an isolate, although such associations may not have been apparent due to the relatively small number of strains exam-ined. Ten isolates with different rDNA profiles (rDNA types A, B, C, D, E, F, J, K, L, and M) were evaluated by RAPD us-ing the primer OPK-17 (27). No clearly evident variations in RAPD pattern were found between any of these strains (data not shown).
Species identification of dermatophytes. The ITS I, 5.8S, and ITS II region amplified fromTrichophyton rubrumby using primers ITS 1 and ITS 4 was 692 bp in length (1). The ampli-fication products obtained from 10 otherTrichophytonspp. and fromMicrosporum persicolorwere of approximately equivalent size (Fig. 3A). The ITS regions amplified fromEpidermophyton floccosum and Microsporum caniswere larger than that of T. rubrumby about 50 bp, while those fromTrichophyton ter-restre,Microsporum gypseum, andMicrosporum audouiniiwere smaller, at approximately 670, 610, and 590 bp, respectively. Amplified products from all species had between two and four recognition sites forMvaI, except forM. audouinii, which had none. Thirteen differentMvaI restriction patterns were pro-duced from 17 dermatophyte species. Nine patterns were unique to one species only (M. audouinii,M. canis,M. gypseum, M. persicolor,E. floccosum,Trichophyton mentagrophytes,T. ter-restre,Trichophyton verrucosum, andTrichophyton violaceum), while four patterns were shared between each of two Tricho-phytonspecies (T. quinkeanumandT. schoenlenii,T.
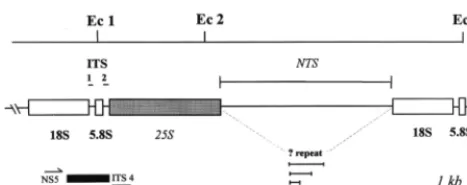
soudane-FIG. 1.EcoRI restriction map of the rDNA repeat unit ofT. rubrum. The
fragment between restriction sites Ec 1 and Ec 2 may encompass the whole of the
25S gene and is of constant length (;3 kb) in all strains. The fragment between
sites Ec 2 and Ec 3 represents the NTS region and the 18S gene and shows
fragment length polymorphisms in severalT. rubrumstrains. One hypothesis to
account for these length variations is the presence of a repetitive element located in the NTS region.
932 JACKSON ET AL. J. CLIN. MICROBIOL.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.612.54.288.71.163.2]nseand T. rubrum,T. equinumand T. tonsurans, andT. con-centricumandT. erinacei) (Fig. 3B).
Three clinical isolates with atypical or equivocal species characteristics could be unambiguously assigned to specific taxa by the ITS PCR method (data not shown).
Stability and reproducibility of strain typing and species identification.Five separate DNA extractions were made over a period of 8 months on strainT. rubrumNCPF 295. Multiple DNA extractions, restriction digests, and Southern blotting were also performed for several other isolates for comparative purposes. In each case, the rDNA types resulting from repeat extractions were unchanged (data not shown).
The results obtained from a comparison ofMvaI ITS diges-tion and convendiges-tional species identificadiges-tion were fully concor-dant for multiple isolates of certain species, including 55T.
ru-brum isolates 18 T. mentagrophytes isolates, 8T. tonsurans isolates, 6T. erinaceiisolates, and 5M. canisisolates (data not shown). The species-characteristic patterns were reproducible irrespective of the strain phenotype, geographic origin, fre-quency of subculture, or storage conditions. Additionally, the method demonstrated the stability ofT. rubrum, in thatT. ru-brumNCPF 295, isolated several decades ago, showed a restric-tion pattern identical to that of strains from the modern era.
DISCUSSION
[image:3.612.148.462.103.505.2]We have developed a molecular method for strain differen-tiation inT. rubrumto determine whether recurrent posttreat-ment onychomycosis is due to treatposttreat-ment failure or to reinfec-tion with a new fungal strain. Strain typing of dermatophytes
FIG. 2. Southern blot ofEcoRI-digested genomic DNA from 14 strains ofT. rubrum, probed as described in Materials and Methods. Two distinct types of restriction
fragment length polymorphism are present in the rDNA repeat of these strains. The six strains (types A to F) illustrated in panel A have a single variable fragment in the size range 4.7 to 5.8 kb, while the eight pattern types (G to N) shown in panel B have multiple variable fragments (either two or four) in the same region. An additional high-molecular-weight variable fragment(s) is present in the 9.0-kb region of all strains. The single, invariant band at 3.0 kb representing fragment Ec 1 to Ec 2 (Fig.
1) is not shown. Molecular weights of standards (in thousands) are given to the right of each panel.l 5molecular weight marker (HindIII-digested lambda DNA).
on May 15, 2020 by guest
http://jcm.asm.org/
has a number of other potential clinical and epidemiological applications. Nail infections due toT. rubrumare increasingly being treated with a new generation of systemic triazole and allylamine antifungal agents. Treatment courses of either con-tinuous or pulsed therapy (22) typically last for several months, increasing the potential for acquisition of resistance to some of these compounds (5). If resistant isolates do emerge, typing methods will allow these strains to be characterized and their occurrence and distribution to be monitored.
The ability to type dermatophytes could also provide new insights into the epidemiology, population biology and patho-genicity of these fungi. For example, our data suggest that only a limited number of strains are prevalent inT. rubrum
infec-tions, with four pattern types (DNA types A to D) predominant among our clinical isolates and one type (DNA type A) ac-counting for over a third of all strains. One reason for this may be a low discrimination index for the typing system we de-scribe. However, this distribution pattern may also have result-ed from a recent and widespread dissemination of these par-ticular strain types, which may possess enhanced infectivity, invasiveness, or other virulence characteristics. The etiology of dermatophyte infection in the United Kingdom is undoubted-ly linked to factors such as recent changes in foot hygiene, combined with an increased usage of communal recreational facilities such as swimming pools and leisure centers. These developments may have provided ideal conditions for the pro-liferation of a single or small number of particularly anthropo-philic or pathogenicT. rubrumclones. This type of strain dis-tribution has been termed epidemic or explosive spread in prokaryotes (14).
The success in typing dermatophytes according to pheno-typic criteria such as colony morphology or biochemical reac-tivity has been limited. Molecular approaches such as RAPD have similarly failed to identify substantive intraspecific poly-morphisms withinT. rubrumor other dermatophyte species (7, 11, 16, 27). What little variations were seen for RAPD between isolates ofT. rubrumwere minor, and reduced-specificity PCR amplifications are notorious for problems of reproducibility (23). These results may reflect an innate lack of chromosomal variation in these fungi, perhaps as a consequence of the clonal population structure suggested above. However, our results demonstrate that substantial molecular diversity is present in the rDNA repeat ofT. rubrum, and interstrain variations else-where in the genome may subsequently be identified by mo-lecular methods other than RAPD, such as microsatellite typ-ing. Interestingly, an early report also demonstrated rDNA variation in the dimorphic pathogenic fungusHistoplasma cap-sulatum(24). This species is a member of the order Onygen-ales, a monophyletic lineage from whose ancestor the family of dermatophyte fungi (Arthrodermataceae) also evolved (9).
The molecular polymorphisms we have identified may be due to variations in the copy number of a repetitive element present in the nontranscribed intergenic regions of the rDNA cistrons. Similar repetitive units have been identified in the rDNA intergenic regions of higher organisms such asXenopus laevis(25) andDrosophila melanogaster(12), as well as in fungi such as Schizophyllum commune (20), Pythium ultimum (8), and C. krusei (3). The size difference separating each of the fragments in types A to D appears to be about 100 to 150 bp, which may indicate the presence of a repeating sequence of this length in the T. rubrum NTS region. We are currently attempting to confirm the existence and genetic identity of this presumptive repetitive element in our laboratory. The size difference separating types D, E, and F is somewhat larger than 150 bp, and fragments of intermediate sizes may be iden-tified from a wider sample of isolates. The multiple banding patterns (types G to M) may be the result of heterogeneities in the number of repeat units within different copies of the rDNA cistrons of individual strains (8). If T. rubrumis diploid, then these isolates may additionally demonstrate heterozygosity in the rDNA interrepeats derived from each chromosomal ho-mologue (13). A third possibility is that these strains represent heterokaryons, with different rDNA polymorphisms originat-ing from each haploid mate (20).
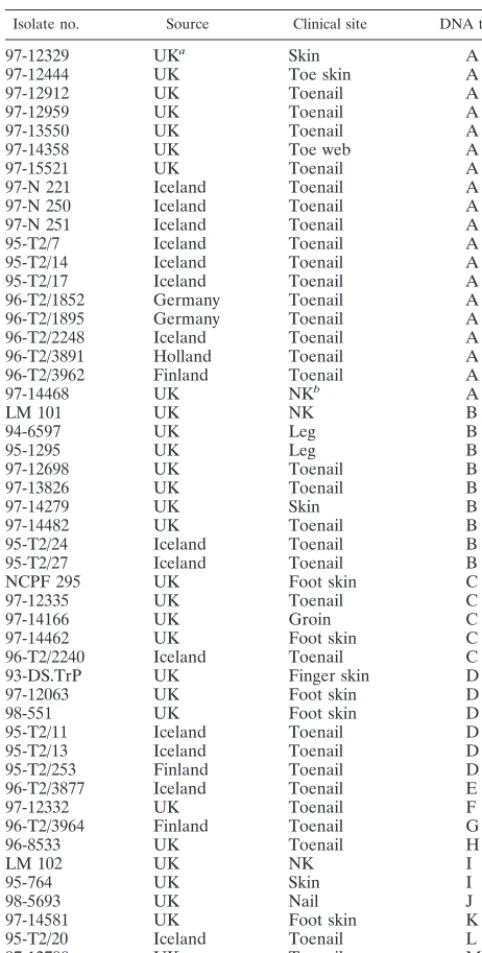
[image:4.612.53.294.92.569.2]While strain typing is useful for studying the clinical and epidemiological aspects of onchomycoses, species identifica-tion has a wider role in monitoring the demographic distribu-tion and changes in frequency of specific dermatophyte infec-tions (6). Accurate identification of dermatophytes can be TABLE 1. Strain details and DNA types for 50 clinical
isolates ofT. rubrum
Isolate no. Source Clinical site DNA type
97-12329 UKa Skin A
97-12444 UK Toe skin A
97-12912 UK Toenail A
97-12959 UK Toenail A
97-13550 UK Toenail A
97-14358 UK Toe web A
97-15521 UK Toenail A
97-N 221 Iceland Toenail A
97-N 250 Iceland Toenail A
97-N 251 Iceland Toenail A
95-T2/7 Iceland Toenail A
95-T2/14 Iceland Toenail A
95-T2/17 Iceland Toenail A
96-T2/1852 Germany Toenail A
96-T2/1895 Germany Toenail A
96-T2/2248 Iceland Toenail A
96-T2/3891 Holland Toenail A
96-T2/3962 Finland Toenail A
97-14468 UK NKb A
LM 101 UK NK B
94-6597 UK Leg B
95-1295 UK Leg B
97-12698 UK Toenail B
97-13826 UK Toenail B
97-14279 UK Skin B
97-14482 UK Toenail B
95-T2/24 Iceland Toenail B
95-T2/27 Iceland Toenail B
NCPF 295 UK Foot skin C
97-12335 UK Toenail C
97-14166 UK Groin C
97-14462 UK Foot skin C
96-T2/2240 Iceland Toenail C
93-DS.TrP UK Finger skin D
97-12063 UK Foot skin D
98-551 UK Foot skin D
95-T2/11 Iceland Toenail D
95-T2/13 Iceland Toenail D
95-T2/253 Finland Toenail D
96-T2/3877 Iceland Toenail E
97-12332 UK Toenail F
96-T2/3964 Finland Toenail G
96-8533 UK Toenail H
LM 102 UK NK I
95-764 UK Skin I
98-5693 UK Nail J
97-14581 UK Foot skin K
95-T2/20 Iceland Toenail L
97-12790 UK Toenail M
97-12496 UK Toenail N
aUK, United Kingdom
bNK, not known.
934 JACKSON ET AL. J. CLIN. MICROBIOL.
on May 15, 2020 by guest
http://jcm.asm.org/
time-consuming and requires extensive familiarity with the mi-croscopical and cultural characteristics of these taxa (21). A molecular solution to problems of species identification in the commercially important black truffle fungi (Tuber melanospo-rum) involved simple restriction analysis of the amplified ITS region (18). The same approach has proved successful for the dermatophytes, although some closely related taxa cannot be distinguished. These includeT. quinkeanumandT. schoenlenii, which have been shown to be closely related by mtDNA re-striction analysis (17). Similarly, the 18S rDNA sequence data forT. soudaneseandT. rubrum, species which have the same MvaI profile, are practically identical (9). No pattern differ-ences were observed amongT. mentagrophytesvarieties inter-digitale, granulosum, mentagrophytes, or sulphureum. Overall, the discrimination achieved byMvaI digestion of the ITS re-gions correlates well with species identification obtained by AP-PCR (7, 11). Further refinements of our system may enable the lack of discrimination evident from the above examples to be overcome. However, we cannot exclude the possibility that some of these apparently anomalous results will be resolved when dermatophyte taxonomy is fully defined on the basis of comparative sequence data.
The MvaI restriction patterns are highly reproducible and
consistent for several species, indicating that the ITS regions in dermatophytes are conserved. This contrasts with pathogenic fungi such asP. carinii, for which sequence variability of the ITS regions is sufficient for strain characterization (10). Strain-specific variations inT. rubrumare located in the NTS rather than the ITS intergenic regions, and the demonstration of these rDNA polymorphisms has provided the first molecular technique for strain differentiation in this species.
ACKNOWLEDGMENT
We are grateful to the Janssen Research Foundation, Beerse, Bel-gium, who provided the financial support for this project.
REFERENCES
1.Berbee, M. L., A. Yoshimura, J. Sugiyama, and J. W. Taylor.1995. Is Peni-cilliummonophyletic—an evaluation of phylogeny in the family Trichoco-maceae from 18S, 5.8S and ITS ribosomal DNA sequence data. Mycologia
87:210–222.
2.Bowman, B. H., J. W. Taylor, and T. J. White.1992. Molecular evolution of
the fungi: human pathogens. Mol. Biol. Evol.9:893–904.
3.Carlotti, A., T. Srikantha, K. Schro¨ppel, C. Kvaal, J. Villard, and D. R. Soll.
1997. A novel repeat sequence (CKRS-1) containing a tandemly repeated
sub-element (kre) accounts for differences betweenCandida kruseistrains
fingerprinted with the probe CkF1,2. Curr. Genet.31:255–263.
[image:5.612.138.474.101.440.2]4.Carlotti, A., F. Chaib, A. Couble, N. Bourgeois, V. Blanchard, and J. Villard.
FIG. 3. Agarose gel electrophoresis of PCR-amplified ITS regions (A) andMvaI restriction digests of amplified ITS products (B) from 17 dermatophyte species.
All products were electrophoresed in 2% agarose gels and stained with ethidium bromide. Lanes: 1,E. floccosumDS.EF; 2,M. audouiniiSJ.EM 4875; 3,M. canis
97/12400; 4,M. gypseumSJ.EM 3928; 5,M. persicolorNCPF 356; 6,T. concentricumNCPF 600; 7,T. erinaceiLM 47; 8,T. quinkeanumLM 54; 9,T. schoenleiniiSJ.EV
327; 10,T. verrucosum98/4545; 11,T. rubrumNCPF 295; 12,T. soudanense98/7676; 13,T. violaceumSJ.EM 7115; 14,T. mentagrophytesDS.TMvM; 15,T. equinum
SJ.A1; 16,T. tonsuransSJ.EM 6717; 17,T. terrestreLM 39.
on May 15, 2020 by guest
http://jcm.asm.org/
1997. Rapid identification and fingerprinting ofCandida kruseiby PCR-based amplification of the species-specific repetitive polymorphic sequence
CKRS-1. J. Clin. Microbiol.35:1337–1343.
5.Evans, E. G. V.1998. Causative pathogens in onychomycosis and the
possi-bility of treatment resistance: a review. J. Am. Acad. Dermatol.38:S32–S56.
6.Ginter, G., E. Reiger, K. Heigl, and E. Propst.1996. Increasing frequency of onychomycoses. Is there a change in the spectrum of infectious agents?
Mycoses39(Suppl. 1):118–122.
7.Gra¨ser, Y., M. el Fari, W. Presber, W. Sterry, and H.-J. Tietz.1998.
Iden-tification of common dermatophytes (Trichophyton,Microsporum,
Epidermo-phyton) using polymerase chain reactions. Br. J. Dermatol.138:576–582. 8.Klassen, G. R., and J. Buchko.1990. Subrepeat structure of the intergenic
region in the ribosomal DNA of the oomycetous fungusPythium ultimum.
Curr. Genet.17:125–127.
9.Leclerc, M. C., H. Philippe, and E. Gue´ho.1994. Phylogeny of dermato-phytes and dimorphic fungi based on large subunit ribosomal RNA sequence
comparisons. J. Med. Vet. Mycol.32:331–341.
10. Lee, C.-H., J. Helweg-Larsen, X. Tang, S. Jin, B. Li, M. S. Bartlett, J.-J. Lu, B. Lundgren, J. D. Lundgren, M. Olsson, S. B. Lucas, P. Roux, A. Cargnel, C. Atzori, O. Matos, and J. W. Smith.1998. Update onPneumocystis carinii
f. sp.hoministyping based on nucleotide sequence variations in internal
transcribed spacer regions of rRNA genes. J. Clin. Microbiol.36:734–741.
11. Liu, D., S. Coloe, J. Pedersen, and R. Baird.1996. Use of arbitrarily primed
polymerase chain reaction to differentiate Trichophyton dermatophytes.
FEMS Microbiol. Lett.136:147–150.
12. Long, E. O., and I. O. Dawid.1979. Restriction analysis of spacers in
ribo-somal DNA ofDrosophila melanogaster. Nucleic Acids Res.7:205–215.
13. Magee, B. B., T. M. d’Souza, and P. T. Magee.1987. Strain and species identification by restriction fragment length polymorphisms in the ribosomal
DNA repeat ofCandidaspecies. J. Bacteriol.169:1639–1643.
14. Maynard Smith, J., N. H. Smith, M. O’Rourke, and B. G. Spratt.1993. How
clonal are bacteria? Proc. Natl. Acad. Sci. USA90:4384–4388.
15. Mochizuki, T., S. Watanabe, and M. Uehara.1996. Genetic homogeneity of
Trichophyton mentagrophytesvar.interdigitale isolated from geographically
distant regions. J. Med. Vet. Mycol.34:139–143.
16. Mochizuki, T., N. Sugie, and M. Uehara.1997. Random amplification of polymorphic DNA is useful for the differentiation of several anthropophilic
dermatophytes. Mycoses40:405–409.
17. Nishio, K., M. Kawasaki, and H. Ishizaki.1991. Phylogeny of the genera
Trichophytonusing mitochondrial DNA analysis. Mycopathologia11:127– 132.
18. Paolocci, F., A. Rubini, B. Granetti, and S. Arcioni.1997. TypingTuber melanosporum and Chinese black truffle species by molecular markers.
FEMS Microbiol. Lett.153:255–260.
19. Radford, S. A., E. M. Johnson, J. P. Leeming, M. R. Millar, J. M. Cornish, A. B. M. Foot, and D. W. Warnock.1998. Molecular epidemiological study of
Aspergillus fumigatusin a bone marrow transplantation unit by PCR
ampli-fication of ribosomal intergenic spacer sequences. J. Clin. Microbiol.36:
1294–1299.
20. Specht, C. A., C. P. Novotny, and R. C. Ullrich.1984. Strain specific
differ-ences in ribosomal DNA from the fungusSchizophyllum commune. Curr.
Genet.8:219–222.
21. Summerbell, R., and J. Kane.1997. Physiological and other special tests for
identifying dermatophytes, p. 45–79.InJ. Kane and R. Summerbell (ed.),
Laboratory handbook of dermatophytes. Star Publishing Co., Belmont, Calif. 22. Tosti, A., B. M. Priaccini, C. Stinchi, N. Venturon, F. Bardazzi, and M. D. Colombo.1996. Treatment of dermatophyte nail infections: an open ran-domized study comparing intermittent terbinafine therapy with continuous terbinafine treatment and intermittent itraconazole therapy. J. Am. Acad.
Dermatol34:595–600.
23. Tyler, K. D., G. Wang, S. D. Tyler, and W. M. Johnson.1997. Factors affecting reliability and reproducibility of amplification-based DNA
finger-printing of representative bacterial pathogens. J. Clin. Microbiol.35:339–
346.
24. Vincent, R. D., R. Goewert, W. E. Goldman, G. S. Kobayashi, A. M. Lam-bowitz, and G. Medoff.1986. Classification ofHistoplasma capsulatum
iso-lates by restriction fragment polymorphisms. J. Bacteriol.165:813–818.
25. Wellauer, P., I. Dawid, D. Brown, and R. Reeder.1976. The molecular basis
for length heterogeneity in ribosomal DNA fromXenopus laevis. J. Mol. Biol.
105:461–486.
26. White, T. J., T. Bruns, and J. Taylor.1990. Amplification and direct
se-quencing of fungal ribosomal RNA genes for phylogenetics, p. 315–322.In
M. A. Innis (ed.), PCR protocols: a guide to methods and applications. Academic Press, London, United Kingdom.
27. Zhong, Z., R. Li, D. Li, and D. Wang.1997. Typing of common
dermato-phytes by random amplification of polymorphic DNA. Jpn. J. Med. Mycol38:
239–246.
936 JACKSON ET AL. J. CLIN. MICROBIOL.