http://dx.doi.org/10.4236/ojas.2015.53030
How to cite this paper: Akbary, P., Mirvaghefi, A.R., Akhlaghi, M. and Fereidouni, M.S. (2015) Influence of Maternal and Larval Immunisation against Lactococcus garviae Infection in Rainbow Trout Oncorhynchus mykiss (Walaum)Lysozyme Ac-tivity and IgM Level. Open Journal of Animal Sciences, 5, 258-269. http://dx.doi.org/10.4236/ojas.2015.53030
Influence of Maternal and Larval
Immunisation against
Lactococcus garviae
Infection in Rainbow Trout
Oncorhynchus
mykiss
(Walaum)
Lysozyme Activity
and IgM Level
Paria Akbary
1,2, Ali Reza Mirvaghefi
2*, Mostafa Akhlaghi
3, Mohammad Saeid Fereidouni
3 1Department of Marine Sciences, Fisheries Group, Chabahar Maritime University, Chabahar, Iran2Department of Natural Resources, Faculty of Fisheries, Tehran University, Tehran, Iran 3Aquatic Animal Health Unit, School of Veterinary Medicine, Shirazu University, Shiraz, Iran
Email: *[email protected]
Received 25 February 2015; accepted 1 June 2015; published 4 June 2015
Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY). http://creativecommons.org/licenses/by/4.0/
Abstract
This study evaluated efficacy of maternal and larval immunisation against Lactococcus garviae in-fection and on the lysozyme and immunoglobulin (IgM) levels in rainbow trout Oncorhynchus my-kiss (Walaum). Forty-eight-day-old larvae (mean weight 96 mg) originating from injected weekly with letrozole and immunised, only immunised and non-immunised parents were experimentally infected with the L. garvieae, and the mortality rate was recorded daily. Larvae were vaccinated by immersion at 58 days post hatch with live L. garvieae (109 cells/mL) for 15 min. Every third day post larvae vaccination, two larvae from each group were collected for analysis lysozyme (by a method based on the ability of lysozyme to lyse the bacterium Micrococcus lysodeikticus) and IgM (by enzyme-linked immunosorbent assay (ELISA)) parameters. Vaccinated and control larvae were tested for protection against L. garvieae 30 days post larvae immunization when the larvae were 88 days old. Larvae were challenged by bath exposure with live L. garvieae (109 cells/mL) for 2 min and monitored for mortality for at least 10 days following challenge. The challenge experi-ment with L. garvieae showed a significant reduction in larvae from immunised (54.44% ± 0.64%) and injected weekly with letrozole and immunised fish (52.96% ± 0.97%) compared to larvae from control fish (62.96% ± 2.22%). Vaccinated larvae originated from injected weekly with le-trozole and immunised parents showed significantly higher lysozyme activity compared to other fish groups. Vaccinated larvae showed significantly less mortality compared to controls. The
tive percent survival (RPS) values of larvae from only immunised, injected weekly with letrozole and immunised and non-immunised parents vaccinated with L. garvieae were 67.36% ± 0.9%, 68.05% ± 0.66%and 48.27% ± 2.79% respectively. The results indicate that the effect of maternal immunization rainbow trout against L. garvieae infection by eliciting the immune responses as in-dicated by an increase in the IgM level and lysozyme activity.
Keywords
Immunisation, Relative Percent of Survival (RPS), Rainbow TroutLarvae, Lactococcus garviae, Immunoglobulin
1. Introduction
Lactococcus garvieae is considered the most important risk factor for the rainbow trout and marine fish industry [1]. Also the rapidly expanding aquaculture industry incurs a substantial economic loss due to lactococcosis caused by L. garvieae resulting in more than 50% mortality over a period of 3 - 7 days [2]. In a study in 2002 in Iran from 235 cases of streptococcosis among rainbow trout collected from farms of Fars province [3], strepto-cocci were isolated from 99 cases. These isolates were biochemically allocated to Streptococcus iniae and L. garvieae. Also L. garvieae outbreaks in farmed rainbow trout are described from different parts of Iran [4] [5]. Therapeutic measures are generally ineffective and therefore, development of vaccines is essential to control the disease [4] [6].
There have been a variety of studies analyzing the effectiveness of immunisation in preventing Lactococcosis in rainbow trout [7]. To prevent rainbow trout lactococcosis caused by L. garvieae, different vaccine formula-tions have been administered by intraperitoneal injection, showing good initial levels of protection but very short duration of immunity and the RPS values obtained with the aqueous vaccine was 40% [7]. Petrie-Hanson and Ainsworth [8] carried out field trials with this type of vaccines in Spanish fish farms and observed mortali-ties of 1.7% in vaccinated fish compared to 6.5% in non-vaccinated fish. The optimal time for vaccination is when fish weight approximately 50 g.
Larvae vaccinated with Photobacterium damsel subsp. piscicida (phdp) and lipopolysaccharides (LPS) in sea bream (Sparus auratus) showed significantly higher lysozyme activity and total immunoglobulin compared to the controls. Vaccinated larvae from both groups showed significantly less mortality compared to the respective controls and RPS values of larvae from immunised parents vaccinated with phdp and LPS was 95.83% and 72.22% respectively [9]. Significant increase of the survival in Atlantic halibut (Hippoglossus hippoglossus L.) larvae was reported after exposure to 50 and 100 µg/mL of LPS from day 20 to 96 and throughout the yolk sac period as compared to untreated larvae [10].
Letrozole (CGS 20264), with commonly used brand name Femara, is a non-steroidal trizole derivation and one of the most potent aromatase inhibitors yet developed [11]. It has been approved for the first line therapy for hormone receptor positive, metastatic breast cancer in postmenopausal women [11] [12]. Letrozole has potential for use to prevent the conversion of androgenic steroids to estrogens [12]. Also it is capable of inhibiting aro-matase 98% - 99% and reducing serum concentrations of estrone and E2 beyond the limit of detection in patients [11] [12].
During the last decade, a number of studies have shown that in addition to their classically described repro-ductive functions, estrogens and androgens also regulate the immune system in teleost [13].
The reproductive-immune interactions studied in fishes indicate that sexual maturation (i.e., gonadal maturity and reproductive activities) can potentially affect both the innate and adaptive immune responses [13]. In fish, the reproductive cycle is marked by seasonal variation in plasma sex-steroids and by changes in immune para-meters which render them more sensitive to the presence of pathogens [14]. Furthermore, immune biomarkers may prove useful for the identification of contamination in the wild [13].
the suppressive effects of E2 on IgM production are not observed in carp and zebra fish (Danio rerio) [15]. Elevation of sex steroids can make fish more vulnerable to several microbial infections during the spawning season and subsequent mortality following the spawning phase. Therefore the overall health and immune status of brood fish is very important not only for breeding performances but also for the health condition of offspring [10] [14] [16]. In this paper, we investigated the effects of endocrine disrupter component (EDC) letrozole on lysozyme and IgM levels after female immunisation of rainbow trout
This study was aimed to determine the effect of rainbow trout maternal and larval iimmunisation and injected of non-steroidal compound, letrozole, two months before spawning season on the susceptibility by L. garvieae
and on lysozyme activity and IgM levels.
2. Materials and Methods
2.1. Bacteria
L. garvieae (EU727199) used in this study was isolated from an outbreak of Lactococcosis in Fars province, Iran. This strain was selected on the basis of its antigenic characteristics that were determined in previous work [17].
2.2. Media and Culture Conditions
Inoculate of L. garvieae were prepared by growing cells overnight on blood agar (BioMerieus, France) at 25˚C for 24 - 48 h. Bacterial cells were collected by centrifugation at 6500 r/30 min at 4˚C and washed three times with phosphate buffered saline (PBS:pH 7.4) and then were re-suspended in PBS at a final concentration of 1.2 × 108 cells/mL. Cultures were checked for purity and the number of colony forming units (CFU) estimated by plate counting [18].
2.3. Preparation of the Bacteria
A formaline-killed vaccine was prepared as previously described [18]. The selected strain of L. garvieae was grown in Trypticase Soy Broth (TSB, Difco) for 48 h, obtaining a final concentration of 1 × 109 cells/mL. Then cells were killed by addition of formalin to achieve a final concentration of 0.3% and incubated for 3 h at 25˚C and then 4˚C overnight.
2.4. Fish and Rearing Conditions
Adult rainbow trout (mean weight approximately 850 g) were maintained in 800 L concrete pond, under natural day length, at the Dalkhan Fisheries Research center of Shiraz, Iran. The water temperature was maintained at 12˚C - 15˚C, with oxygen 5 mg/L and pH 7.5. The total capacity of the system was 9 m3 and the exchange of 25% water occurred every day. The levels of ammonia, nitrites and nitrates were monitored daily. Fish were fed 2 times per day with pellets a commercial feed at 1% of body weight.
2.5. Vaccination and Injection of Letrozole of Brood Fish
2.6. Larvae Rearing Conditions
The completion of yolk sac absorption was achieved very early at day 14. Larvae were fed 8 times per day with pellets from a commercial feed at 5% of body weight. After weaning they were transferred to Department of Aquatic Animal Health Unit, School of Veterinary Medicine, and Shiraz University, Iran at day 42 of age. Lar-vae were kept in a tank system. The water temperature was maintained 12˚C - 15˚C, oxygen 5 mg/L and pH 7.8. The water was aerated with the aid of an air pump with capacity of 1 m3/h. Each tank had 500 L holding capacity, the total capacity of system was 6 m3 and the water exchange occurred every 1 h. The levels of ammonia, ni-trites and nitrates were monitored daily. Larvae were fed two times per day with pellets from a commercial feed at 1% of body weight. Experimentally infected larvae were kept in a static water aquaria system and fed two times a day. All other conditions were as described previously.
2.7. Susceptibility Test of Rainbow Trout Larvae from Injected Weekly with Letrozole and
Immunised, Only Immunised and Non-Immunised Parents
A virulent bath challenge was used to evaluate differences in protective immunity in larvae from vaccinated and non-vaccinated parents when larvae were 48 days old. The challenge baths were prepared by cultivating the bacteria in Trypticase Soy Broth (TSB; Difco) for 48 h with continuous shaking. The cultures were washed twice in phosphate-buffered saline (PBS, pH 7.2). After final centrifugation, the suspension was diluted to a density giving 50% transmission at 520 nm in a spectrophotometer, corresponding to approximately 1.2 × 108 cells/mL. For the challenge experiments, triplicate groups of forty-eight days old fry originating from non-im- munised (NIM), immunised (IM) only and injected with letrozole and immunised (Let.IM) broodstock were immersion for 2 min with dilutions of 1.2 × 108 bacterial cells/mL. Control non infected larvae (CNI) of each group received exactly the same treatment with sterile sea water. Thirty larvae were used in each sub group. Mortalities were monitored for period of 10 days and fish were tested with sampling of kidney and viewing it by microscope to confirm the cause of morbidity.
2.8. Bath Vaccination Experiments of Larvae from Immunised and Non-Immunised Fish
For the bath vaccination experiments triplicate groups of fifty-eight days old larvae originating from non immu-nized (NIM), immuimmu-nized (IM) only and injected with letrozole and immuimmu-nized (Let.IM) females were vacci-nated by immersion with 1.0 mL of formalin (3%) inactivated L. garvieae (109 cells/mL). Also some fifty-eight days old larvae originating from NIM, IM and Let.IM females were vaccinated only with phosphate buffered sa-line (PBS) as control non immunized (CNI) larvae of each group.
2.9. Collection of Larvae after Vaccination, Weight and Homogenates
Every third day post-larvae vaccination, two larvae from each group were collected for analysis of lysozyme ac-tivity and total immunoglobulin. Larvae were washed two times with sterile PBS, pH 7.2, homogenized with 1 volume of phosphate-buffered saline and centrifuged at 3000 r/10 min at 4˚C. The supernatants were collected, centrifuged once at 3000 g for 5 min and stored at −20˚C. Samples from the different larval rearing were col-lected, rinsed with distilled water, and their wet body weight measured and recorded. Weight was recorded in triplicate groups.
2.10. Susceptibility Test of Vaccinated Larvae from Injected Weekly with Letrozole and
Immunised, Only Immunised and Non-Immunised Parents
Vaccinated and control larvae were tested for protection against Lactococcus garvieae 30 days post immunisa-tion when the larvae were 88 days old. Larvae were challenged by bath exposure as described above with the previously dose (1.2 × 108 cells/mL). Thirty larvae were used in each group and the experiment was run in trip-licate. Mortality was recorded for 10 days and the RPS was calculated:
RPS= (1 − %mortality of immunised larvae/% mortality of control non-immunised larvae) × 100 [20].
2.11. Lysozyme Activity
lysozyme to lyse the bacterium Micrococcus lysodeikticus [21]. In a 96-well microplate, 250 μL of larval ho-mogenates in four twofold serial dilutions in PBS (0.05 M, PH 7.2) were mixed with 250 μL of 0.7 mg/mL sus-pension of Micrococcus lysodeikticus (Sigma) in phosphate buffer (175 mL). The micro plate was incubated at 24˚С and O.D. was read at 450 nm at 0, 15, 30 and 60 min. For a positive and a control, serum was replaced by hen egg white lysozyme (serial dilutions starting at 1.6 μg/mL) and buffer respectively. Data were expressed ei-ther as μg/mL.
2.12. Enzyme Linked Immunosorbent Assay (ELISA) for Determination of Total
Immunoglobulin
The IgM levels of Larval extracts of fish were assessed with ELISA by the method of Bakopoulos, Peric [22] with some modification. 50 μL of serial dilutions of the sera (1:50) were added to the appropriate micro titer plate wells with a HRP-conjugated IgM and antibody preparation specific for IgM and incubated. Then substrate solutions were added to each well. The enzyme-substrate reaction was terminated by the addition of a sulphuric acid solution and the color change was measured spectrophotometrically at a wavelength of 450 nm ± 2 nm.The concentration of steroid in the samples was then determined by comparing the O.D of the samples to the stan-dard curve. All samples were placed in triplicate on the plates and the mean and stanstan-dard deviation (S.D) were calculated for each sample IgM concentration. Data were expressed either as μg/g of wet body weight larvae.
2.13. Statistical Analysis
Data were evaluated using one-way analysis of variance (ANOVA). Groups were considered to be significantly different if P < 0.05. When a significant F value was obtained for ANOVA the differences between all groups were tested by using Duncan ̓s multiple range test . All statistics were performed using SPSS for windows ver-sion 16.
3. Results
3.1. Susceptibility Test of Larvae from Injected Weekly with Letrozoleand Immunized
(Let.IM), Only Immunized (IM) and Non-Immunized (NIM) Parents
The larvae originating from Let.IM and IM parents showed delayed onset and reduced mortality rates in com-parison to the larvae originating from NIM parents. The challenge programmed using with 1.2 × 108 cells/mL of
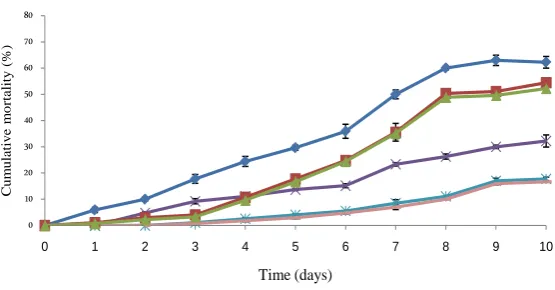
L. garviae in the larvae from each group indicated a cumulative mortality rate was significantly lower in the lar-vae from Let.IM parents (52.96% ± 0.97%) than the larlar-vae from IM (54.44% ± 0.64%) and from NIM (62.96% ± 2.22%) parents. Cumulative mortality rate, in the larvae originated from IM and Let.IM parents had not signifi-cantly different (Table 1). The mortality rate was again different between larval groups, with the mortalities of larvae originating from Let.IM and IM broodstock beginning between day3 while for the offspring of NIM par-ents mortalities started from day 1 (Figure 1).
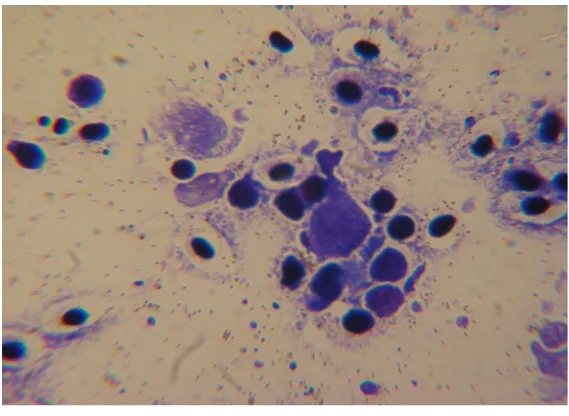
Microscopic evaluation (×100) of the kidney tissue confirmed the cause of morbidity (L. garvieae) (Figure 2).
The RPS values of larvae from only immunised, injected weekly with letrozole and immunised and non-im- munised parents were66.41% ± 1.20%, 66.42% ± 0.83%, and 53.88% ± 0% respectively. Significant difference (P < 0.05) was found between IM and NIM, also Let.IM and NIM. No significant difference (P > 0.05) was found among IM and Let.IM.
3.2. Weight of Larvae
The wet body weight of vaccinated larvae was recorded in three groups from day 0 to day 30 (Figure 3). A ference in body weight increase was observed in larvae originating from Let.IM, IM and parents, but this dif-ference was non-significant (P > 0.05). In vaccinated larvae from immunised (IM), injected weekly with letro-zole and immunised (Let.IM) and from non-immunised (NIM) parents vaccinated with L.garvieae, the body weight was 252.67 ± 0.66, 250 ± 1.15 and 251.33 ± 1.33 mg respectively.
3.3. Lysozyme Activity
Figure 1. Cumulative percent mortality due of larvae from injected weekly with letrozole and immunised (Let.IM) and only immunised (IM) non-im- munised (NIM) broodstock Control Non-infected (CNI). Each data point represents the mean (±S.E.) of triplicates.
Figure 2. Microscopic evaluation (×100) of the kidney tissue to confirm the
cause of morbidity.
Table 1. Difference tested by one-way ANOVA for Cumulative mortality (%) in larvae* of experimental groups.
Groups/days 1 2 3 4 5 6 7 8 9 10
NIM (CNI) A BCD AB A B A A A B A
IM (CNI) A AB A A AB A A A A A
Let.IM (CNI) A A A A A A A A A A
NIM (L) B D C C D C C C D C
IM (L) A C B B C B B B C B
Let.IM (L) A BC AB B C B B B C B
*Larvae from immunised (IM), injected weekly with letrozole and immunised (Let.IM) and non-immunised (NIM) parents Control Non-infected
(NIC). Each data point represents the mean (±S.E.) of triplicates..different letters indicate a significant difference (P < 0.05).
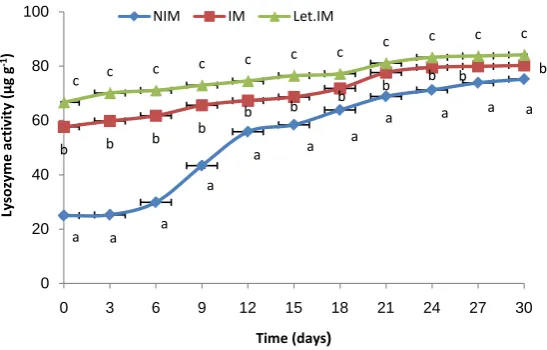
lysozyme activities was observed. Significant difference (P < 0.05) was found among activities in IM, Let.IM and NIM groups (Figure 4).
0 20 40 60 80 100
0 2 4 6 8 10
%
Cu
mu
la
tiv
e mo
rt
al
ity
Time (days) NIM (CNI
[image:6.595.89.539.538.660.2]Figure 3. The wet body weight of vaccinated larvae from injected weekly with letrozole and immunized (Let.IM), only immunised (IM) and non-im- munised (NIM) broodstock Each data point represents the mean (± S.E) of triplicates. Similar values among the groups are identified by the same super-script letter (P > 0.05).
Figure 4. Total lysozyme level changes (μg/g wet body weight) in larvae
from only immunised (IM), injected weekly with letrozole and immunised and non-immunised (NIM) parents Each data point represents the mean (±SE) of triplicates. Different letters indicate a significant difference (P < 0.05).
3.4. IgM Levels
Data on IgM in the larval extracts were shown in Figure 5. Significant difference (P < 0.05) was found among immunoglobulin levels between Let.IM, IM and NIM. No significant difference was found between IM and Let.IM (P > 0.05).
3.5. Susceptibility Test of Vaccinated Larvae from Injected Weekly with Letrozole and
Immunised, Only Immunised and Non-Immunised Parents
Data on cumulative mortality rate in immunised (IM (L) and control non immunized (CNI) larvae of each fe-male (IM, Let.IM and NIM fefe-males) after challenging with live Lactococcus garvieae (L)(1.2 × 108 cells/mL) were shown in Figure 6. The cumulative mortality rate was significantly lower in immunised larvae originating from Let.IM and IM females compared to immunised larvae from NIM females. Significant (P < 0.05) was
b b a 0 50 100 150 200 250 300
0 3 6 9 12 15 18 21 24 27 30
W ei gh t ( m g) Time (days) NIM (L
( (IM(L (Let.IM (L
00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00
0 3 6
00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00 0 3 00
0 3 6 9 12 15 18 21 24 27 30
a a a
a
a a a
a a a a
b b b
b b
b b
b b b b
c c c c
c c c c
c c c
0 20 40 60 80 100
0 3 6 9 12 15 18 21 24 27 30
Lys oz ym e ac tiv ity ( μ g g -1) Time (days)
[image:7.595.178.452.339.514.2]Figure 5. Total IgM level changes (μg/g wet body weight) in larvae from immunised (IM) , injected weekly with letrozole and immunised (Let.IM) and non-immunised (NIM) parents after immunised Each data point represents the mean (±SE) of triplicates. Different letters indicate a significant difference (P < 0.05).
Figure 6. Cumulative mortality with LD50 infection of Lactococcus garviea
(L) in larvae from immunised (IM), injected weekly with letrozole and im-munised (Let.IM) and non-imim-munised (NIM) parents. Control non-immu- nized (CNI). Each data point represents the mean (±S.E.) of triplicates.
found among the cumulative mortality rate between IM (L), IM (CNI) and NIM (CNI), NIM (L) groups and Let.IM (L), Let.IM (CNI) and NIM (CNI), NIM (L) (Table 2).
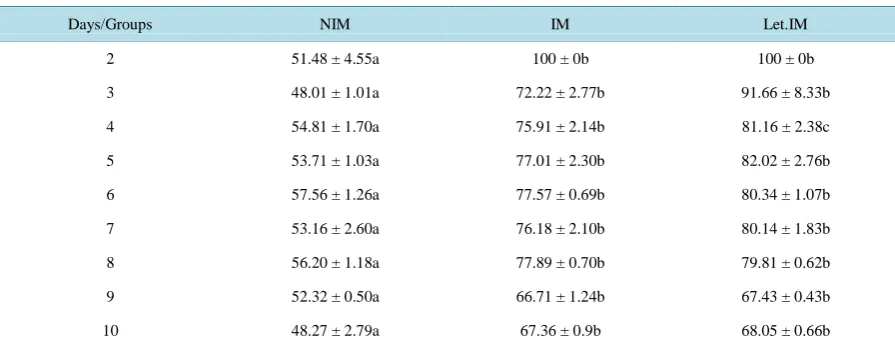
The RPS values of larvae from immunised, injected weekly with letrozole and immunised and non-immu- nised parents vaccinated with L. garvieae was 67.36% ± 0.9%,68.05% ± 0.66%and 48.27% ± 2.79% respec- tively (Table 3). Significant difference (P < 0.05) was found between IM and NIM, also Let.IM and NIM. No significant difference was found between IM and Let.IM (P > 0.05).
4. Discussion
The aquaculture industry is recently facing a serious setback due to infectious [23] diseases leading to severe economic loss [24]. Although mortalities have been reported in all stages of life, maximum mortalities in all the cultivated fish species have been reported at larval stages. Due to lack of immune competence at the early stages [25], an alternate method was evaluated for enhancing the resistance for more availability of healthy fish seed by immunisation.
Delamare-Deboutteville, Wood [26] used immunisation (i.e. by injection and immersion in formalin killed
a a a
a a
a a a
a a
a
b b
b b
b b
b b
b b
b
0 2 4 6 8 10 12 14
0 3 6 9 12 15 18 21 24 27 30
IgM
(
μ
g
g
-1)
Time (days)
NIM IM Let.IM
0 10 20 30 40 50 60 70 80
0 1 2 3 4 5 6 7 8 9 10
C
u
m
u
la
tiv
e
m
o
rta
lity
(
%
)
Time (days) NIM (CNI
[image:8.595.174.452.340.485.2]Table 2. Difference tested by one-way ANOVA for Cumulative mortality (%) larvae* of experimental groups after immu-nized by bath exposure for 15 min with formalin (3%) treated L.garvieae 109 cells.
Groups/days 1 2 3 4 5 6 7 8 9 10
NIM (CNI) B D D C D D D D D D
IM (CNI) A B B B C C C C C C
Let.IM (CNI) A B AB B C C C C C C
NIM (L) A C C B B B B B B B
IM (L) A A A A A A A A A A
Let.IM (L) A A A A A A A A A A
*
Larva from immunized (IM), injected weekly with letrozole and immunized (Let.IM) and non-immunised (NIM) parents Control non-immunised (CNI). Different letters indicate a significant difference (P < 0.05).
Table 3. Difference tested by one-way ANOVA for RPS values (%) in larvae* of experimental groups after immunised by
bath exposure for 15 min with formalin (3%) treated L. garvieae 109 cells/mL.
Days/Groups NIM IM Let.IM
2 51.48 ± 4.55a 100 ± 0b 100 ± 0b
3 48.01 ± 1.01a 72.22 ± 2.77b 91.66 ± 8.33b
4 54.81 ± 1.70a 75.91 ± 2.14b 81.16 ± 2.38c
5 53.71 ± 1.03a 77.01 ± 2.30b 82.02 ± 2.76b
6 57.56 ± 1.26a 77.57 ± 0.69b 80.34 ± 1.07b
7 53.16 ± 2.60a 76.18 ± 2.10b 80.14 ± 1.83b
8 56.20 ± 1.18a 77.89 ± 0.70b 79.81 ± 0.62b
9 52.32 ± 0.50a 66.71 ± 1.24b 67.43 ± 0.43b
10 48.27 ± 2.79a 67.36 ± 0.9b 68.05 ± 0.66b
*
Larvae from immunised (IM), injected weekly with letrozole and immunised (Let.IM) and non-immunised (NIM) parents. In a column different let-ters indicate a significant difference (P < 0.05).
bacteria) to look at the possibility of vaccination of Lates calcarifer against streptococcus (entrococcus). After one month, a positive effect vaccination was observed in immunised fish. Also the percent of mortality in con-trol group was 75%. In our experiment, rainbow trout were vaccinated with L. garviea bacterial cells. Larvae originating from IM, Let.IM and NIM parents were subjected to experimental infectivity trials with L. garviea at day 48 post-hatch. The larvae originating from IM and Let.IM parents showed delayed onset and reduced mor-tality rates in comparison to the larvae originating from NIM parental group. The present results are comparable with other studies on transfer of maternal immunity in some fish species. In the sea bream, plaice, tilapia and red sea bream, transfer of specific protection against various pathogens has been reported [10] [14] [16]. However, experimental immunisation of Atlantic salmon brood females against yearsiniosis did not protect the offspring against the disease although specific antibodies were demonstrated in the eggs and fry [10]. Similarly, transfer of maternal antibody from red sea bream mother fish, immunised against Vibrio sp., and did not have protective value for the larvae. This is in accordance with the fact that the concentration of maternal IgM in the larval stag-es is generally greatly reduced from that observed in the ovarian stagstag-es [14]. The ability of larvae originating from Let.IM and IM parents to respond better to infectious was clearly seen in this study (RPS), Cumulative mortalities in larvae originating from Let.IM and IM parents in comparison to larvae originating from NIM group revealed a difference in favour about 10% (Figure 1). The mechanism underlying the transfer of maternal immunity and protection in offspring by injection of letrozole and immunisation of fish prior to spawning season needs further studies.
[image:9.595.89.537.288.461.2]vac-cinated via immersion with L. garviea bacterial cells at day 58 (128 mg). The following 30 days larval extracts were evaluated for the presence of non-specific (lysozyme) and specific (IgM) humoral immune parameters and at day 88 they were subjected to experimental infectivity trials with L. garviea virulent bacterial cells in order to assess the effect of the above vaccinations on the protection of larvae.
The effectiveness of vaccination of cultured fish heavily depends on the degree of maturity of their immune system [14]. The first appearance of surface IgM in lymphocytes varies considerably in different fish species, this appearance being generally more dependent on size than age [27] [28], although this is not always the case [28]. Fresh water species like rainbow trout and channel cat fish show the first appearance of surface IgM at about 1 week after hatching [28]. Fish larval are very small in size and fragile at the early stages of their life cycle and vaccination by injection is unfeasible. Immersion and oral vaccination is an effective is an effective and practical method for mass vaccination of fish larvae.
The wet body weight of larvae was similar and no significant difference (P > 0.05) was found in all groups (Figure 3).
Higher lysozyme levels were found in immunised larvae originating from immunised IM and Let.IM females. Significant difference (P < 0.05) was found among lysozyme activity between IM and NIM (CNI) larvae and Let IM and NIM (Figure 4). The present result is comparable with investigation of Hanif, Bakopoulos [9] on seabream that larvae vaccinated with Photobacterium damsel subsp. piscicida and liopo lysaccharides showed significantly lysozyme activity compared to the controls [9]. Lysozyme is a bactericidal enzyme, involved in hydrolysis of the β-(1, 4) linked glycoside bonds of bacterial cell wall peptidoglycans. Lysozyme has also been detected in oocytes, fertilized eggs and larval stages of several species, for examples eggs of coho salmon and other salmonids [29]. Also there are several reports of lysozyme isolated from fish serum and ova, being bacte-ricidal even for important fish pathogens like Edwardsiella tarda [25], Escherichia coli [30], A. salmoncida and
A. hydrophila [16] and Photobacterium damsel [9].
Higher total immunoglobulin levels were found in immunised larvae originating from IM and Let.IM females. Significant difference was found among IgM levels between IM, and NIM (CNI) larvae and between Let IM and NIM) (Figure 5). The present result is comparable with investigation of Hanif, Bakopoulos [9] on seabream that larvae vaccinated with Photobacterium damsel subsp. piscicida and liopo lysaccharides showed significantly total IgM levels compared to the controls [9].
Sex hormones are usually elevated during spawning season of fishes and the rise of sex hormones can se-verely affect the immunity of fishes [13]. It is now well estabilished that estrogen receptors [31]. Mechanistical-ly estrogens can influence immune responses by acting directMechanistical-ly on Mechanistical-lymphoid tissue or indirectMechanistical-ly through mul-tiple non lymphoid targets [31] [32]. For example, in O. mykiss and gilthead sea bream, shown that the rise of sex hormones can suppress the plasma IgM and IgM secreting cells during spawning period leading to immu-nosuppressive condition [32] [33]. Due to, elevation of sex steroids can make fish more vulnerable to several microbial infections during the spawning season and subsequent mortality following the spawning phase. We used the endocrine disrupter component (EDC) letrozole for rising lysozyme and IgM levels in immunised fe-males. In the present study, effects of letrozole on sex steroid hormones have not been shown.
Eighty-eight days old immunised and non immunised larvae originating from IM, Let.IM and NIM females were subjected to bath challenges using the LD50 dose (1.2 × 108 cells/Ml). RPS values were calculated for lar-vae from three groups (Table 3). Significant difference was observed among the immunised groups (IM and Let.IM) and NIM parents. So it is evident that the immune system of rainbow trout larvae is efficient at this age (128mg) to respond against antigens. It is concluded that larvae can be vaccinated successfully as early as early as 58-days post hatch (128 mg) and that these larvae can be protected against L. garviea.
Rainbow trout is able for antibody production against Aeromonas hydrophila when exposed at 3 weeks post- hatch. The minimal size at which a response was detected in sockey (Oncorhynchus nerka), pink (O. gorbuscha), chum, coho, Chinook salmon (O. tshawytscha) and rainbow trout (O. mykiss) was 1.0 - 2.5 g. Size was also shown to be a better indicator of immunological maturation than age in Sebastiscus marmoratus [29]. This is not comparable with our present study because immune response was detected in rainbow trout larvae when their body weight was 128 mg.
three groups (Let.IM, IM and NIM) showed less mortality, so immersion vaccination can be used in larvae at the age of 58 days 128 mg) when the immune system can respond well against antigen. Hence, strategies should be put in place to immunise brood stocks of rainbow trout prior to breeding for better health management and to overcome the challenges in vaccinating large number of larvae at a time.
Acknowledgements
The authors thank the Ministry of Science, Research and Technology, Islamic Republic of Iran, for financial support. Grant No.: 89-345-189.
References
[1] Kim, J.H., Go, J., Cho, C.R., Kim, J.I., Lee, M.S. and Parka, S.C. (2013) First Report of Human Acute Acalculous Cholecystitis Caused by the Fish Pathogen Lactococcus garvieae. Journal of Clinical Microbiology, 51, 712-714.
http://dx.doi.org/10.1128/JCM.02369-12
[2] Harikrishnan, R., Balasundaram, C. and Heo, M.S. (2010) Lactobacillus sakei BK19 Enriched Diet Enhances the Im-munity Status and Disease Resistance to Streptococcosis Infection in Kelp Grouper, Epinephelus bruneus. Fish and Shellfish Immunology, 29, 1037-1043. http://dx.doi.org/10.1016/j.fsi.2010.08.017
[3] Hosseini, M.H., Akhlaghi, M. and Moazzeni Jula, G. (2011) Experimental Vaccine against Lactococcosis in Cultured Rainbow Trout (Oncorhynchus mykiss). Archives of Razi Institute, 66, 51-57.
http://www.archrazi.com/browse.php?a_code=A-10-1-1
[4] Haghighi Karsidani, S., Soltani, M., Nikbakhat-Brojeni, G., Ghasemi, M. and Skall, H.F. (2010) Molecular Epidemi-ology of Zoonotic Streptococcosis/Lactococcosis in Rainbow Trout (Oncorhynchus mykiss) Aquaculture in Iran. Iran Journal of Microbiology, 2, 198-209. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3279792/
[5] Erfanmanesh, M.S., Pirali, E., Mohammadian, S. and Taherimirghaed, A. (2012) Genetic Characterization of Strepto-coccus iniae in Diseased Farmed Rainbow Trout (Onchorhynchus mykiss) in Iran. Scientific World Journal, 594073- 594079.
[6] Ricci, G., Ferrario, C., Borgo, F., Rollando, A. and Fortina, M.G. (2012) Genome Sequences of Lactococcus garvieae TB25, Isolated from Italian Cheese, and Lactococcus garvieae LG9, Isolated from Italian Rainbow Trout. Journal of Bacteriology, 194, 1249-1250.http://dx.doi.org/10.1128/JB.06655-11
[7] Ravelo, C., Magariños, B., Herrero, M.C., Costa, L., Toranzo, A.E. and Romalde, J.L. (2006) Use of Adjuvanted Vac-cines to Lengthen the Protection against Lactococcosis in Rainbow Trout (Oncorhynchus mykiss). Aquaculture, 251, 153-158. http://dx.doi.org/10.1016/j.aquaculture.2005.05.027
[8] Petrie-Hanson, L. and Ainsworth, A.J. (2001) Ontogeny of Channel Catfish Lymphoid Organs. Veterinary Immunology and Immunopathology, 81, 113-127. http://dx.doi.org/10.1016/S0165-2427(01)00331-2
[9] Hanif, A., Bakopoulos, V., Leonardos, I. and Dimitriadis, G.J. (2005) The Effect of Sea Bream (Sparus aurata) Broodstock and Larval Vaccination on the Susceptibility by Photobacterium damsela subsp. Piscicida and on the Hu-moral Immune Parameters. Fish and Shellfish Immunology, 19, 345-361. http://dx.doi.org/10.1016/j.fsi.2004.12.009
[10] Zapata, A., Diez, B., Cejalvo, T., Frías, C.G. and Cortés, A. (2006) Onetogeny of the Immune System of Fish. Fish and Shellfish Immunology, 20, 126-136. http://dx.doi.org/10.1016/j.fsi.2004.09.005
[11] Pavone, M.E. and Bulun, S.E. (2012) Aromatase Inhibitors for the Treatment of Endometriosis: A Review. Fertility and Sterility, 98, 1370-1379. http://dx.doi.org/10.1016/j.fertnstert.2012.08.053
[12] Chumsri, S., Howes, T., Bao, T., Sabnis, G. and Brodie, A. (2011) Aromatase, Aromatase Inhibitors, and Breast Can-cer. The Journal of Steroid Biochemistry and Molecular Biology, 125, 13-22.
http://dx.doi.org/10.1016/j.jsbmb.2011.02.001
[13] Milla, S., Depiereux, S. and Kestemont, P. (2011) The Effects of Estrogenic and Androgenic Endocrine Disruptors on the Immune System of Fish: A Review. Ecotoxicology, 20, 305-319. http://dx.doi.org/10.1007/s10646-010-0588-7
[14] Zhang, S.C., Wang, Z.P. and Wang, H.M. (2013) Maternal Immunity in Fish. Developmental & Comparative Immu-nology, 39, 72-78. http://dx.doi.org/10.1016/j.dci.2012.02.009
[15] Li, L., Zhang, S.C., Tong, Z. and Liu, J. (2010) In Vivo Effect of 17-β-Estradiol on Plasma Immunoglobulin Levels and Leukocyte Density in Zebrafish Danio rerio.Chinese Journal of Oceanology and Limnology, 28, 527-532.
http://dx.doi.org/10.1007/s00343-009-9048-8
[16] Swain, P. and Nayak, S.K. (2009) Role of Maternally Derived Immunity in Fish. Fish & Shellfish Immunology, 27, 89- 99. http://dx.doi.org/10.1016/j.fsi.2009.04.008
Lactococcus garvieae from Diseased Rainbow Trout (Oncorhynchus mykiss, Walbaum) Cultured in Iran. Iranian Journal of Veterinary Research, 11, 342-350.
[18] Romalde, J.L., Alvarez, A.L., Ravelo, C., Toranzo, A.E. and Mendez, J.B. (2004) Oral Immunization Using Alginate Microparticles as a Useful Strategy for Booster Vaccination against Fish Lactococcosis. Aquaculture, 236, 119-129.
http://dx.doi.org/10.1016/j.aquaculture.2004.02.028
[19] Shilling, A.D., Carlson, D.B. and Williams, D.E. (1999) Rainbow Trout, Oncorhynchus mykiss, as a Model for Aro-matase Inhibition. The Journal of Steroid Biochemistry and Molecular Biology, 70, 89-95.
http://dx.doi.org/10.1016/S0960-0760(99)00090-4
[20] Bridle, A.R., Koop, B.F. and Nowak, B.F. (2012) Identification of Surrogates of Protection against Yersiniosis in Im-mersion Vaccinated Atlantic Salmon. PLoS ONE, 7, 40841-40850. http://dx.doi.org/10.1371/journal.pone.0040841
[21] Ellis, A.E. (1990) Lysozyme Assays. In: Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S. and Van Muis-winkel, W.B., Eds., Techniques in Fish Immunology,SOS Publications, Fair Haven, 101-103.
[22] Bakopoulos, V., Peric, Z., Rodger, H., Adamms, A. and Richards, R. (1997) First Report of Fish Pasteurellosis from Malta. Journal of Aquatic Animal Health, 9, 26-33.
http://dx.doi.org/10.1577/1548-8667(1997)009<0026:FROFPF>2.3.CO;2
[23] Buschmann, A.H., Tomova, A., López, A., Maldonado, M.A., Henríquez, L.A., Ivanova, L., et al. (2012) Salmon Aq-uaculture and Antimicrobial Resistance in the Marine Environment. PLoS ONE, 7, 42724-42735.
http://dx.doi.org/10.1371/journal.pone.0042724
[24] Peeler, E.J. and Taylor, N.G.H. (2011) The Application of Epidemiology in Aquatic Animal Health—Opportunities and Challenges. Veterinary Research, 42, 94-109. http://www.veterinaryresearch.org/content/42/1/94
http://dx.doi.org/10.1186/1297-9716-42-94
[25] Wang, Z., Zhang, S., Tong, Z., Li, L. and Wang, G. (2009) Maternal Transfer and Protective Role of the Alternative Complement Components in Zebrafish Danio rerio. PLoS ONE, 4, 4498-4505.
http://dx.doi.org/10.1371/journal.pone.0004498
[26] Delamare-Deboutteville, J., Wood, D. and Barnes, A.C. (2006) Response and Function of Cutaneous Mucosal and Se-rum Antibodies in Barramundi (Lates calcarifer) Acclimated in Seawater and Freshwater. Fish & Shellfish Immunolo-gy, 21, 92-101. http://dx.doi.org/10.1016/j.fsi.2005.10.005
[27] Rombouta, J.H.W.M., Abellic, L., Picchiettid, S., Scapigliatid, G. and Kiron, V. (2011) Teleost Intestinal Immunology. Fish & Shellfish Immunology, 31, 616-626. http://dx.doi.org/10.1016/j.fsi.2010.09.001
[28] Salinas, I., Zhang, Y.A. and Sunyer, J.O. (2011) Mucosal Immunoglobulins and B Cells of Teleost Fish. Developmen-tal & Comparative Immunology, 35, 1346-1365. http://dx.doi.org/10.1016/j.dci.2011.11.009
[29] Uribe, C., Folch, H., Enriquez, R. and Moran, G. (2011) Innate and Adaptive Immunity in Teleost Fish: A Review. Veterinarni Medicina, 56, 486-503.
[30] Van Herreweghe, J.M., Vanderkelen, L., Callewaert, L., Aertsen, A., Compernolle, G., Declerck, P.J., et al. (2010) Lysozyme Inhibitor Conferring Bacterial Tolerance to Invertebrate Type Lysozyme. Cellular and Molecular Life Sciences, 67, 1177-1188. http://dx.doi.org/10.1007/s00018-009-0241-x
[31] Thilagam, H., Gopalakrishnan, S., Bo, J. and Wang, K.J. (2009) Effect of 17-β-Estradiol on the Immunocompetence of Japanese Sea Bass (Lateolabrax japonicus). Environmental Toxicology and Chemistry, 28, 1722-1731.
http://dx.doi.org/10.1897/08-642.1
[32] Akbary, P., Mirvaghefi, A.R., Akhlaghi, M., Amiri, B.M. and Fereidouni, M.S. (2013) Effects of the Aromatase Inhi-bitore Letrozole on Serum Immunoglobulin and Lysozyme Levels in Immunized Rainbow Trout (Oncorhynchus my-kiss Walbaum) Females. International Journal of Aquatic Biology, 1, 273-280.
http://www.npajournals.com/ijab/index.php/ijab/article/view/115
[33] Sun, L., Shao, X., Wu, Y., Li, J., Zhou, Q., Lin, B., et al. (2011) Ontogenetic Expression and 17β-Estradiol Regulation of Immune-Related Genes in Early Life Stages of Japanese Medaka (Oryzias latipes). Fish & Shellfish Immunology, 30, 1131-1137. http://dx.doi.org/10.1016/j.fsi.2011.02.020
[34] Ángeles Esteban, M., Cuesta, A., Chaves-Pozo, E. and Meseguer, J. (2013) Influence of Melatonin on the Immune System of Fish: A Review. International Journal of Molecular Sciences, 14, 7979-7999.