INTRODUCTION
A limited number of intercellular signaling pathways are used reiteratively in different contexts to control the development of metazoans. An unanswered question in the study of organism development is how do these signals cooperate and how are they are modulated to give rise to different tissue- and time-dependent responses?
The Notch (N) signaling pathway, which is involved in a vast number of processes in Drosophilaand vertebrates, is a notable example (reviewed by Artavanis-Tsakonas et al., 1999; Doroquez and Rebay, 2006; Lai, 2004). Strikingly, a relatively simple conserved core pathway controls the entire range of N responses. The N protein is a transmembrane receptor that is modified through glycosylation and cleavage (S1) in the Golgi (Logeat et al., 1998; Moloney et al., 2000; Okajima and Irvine, 2002). Two transmembrane ligands are known in Drosophila: Delta (Dl) and Serrate (Ser) (reviewed by Fleming et al., 1997). Upon binding, ligands induce two additional proteolytic events of N, the S2 and S3 cleavages, leading to the release of the intracellular domain (Nintra), the active form of N (Pan and Rubin, 1997) (reviewed by Selkoe and Kopan, 2003). Nintratranslocates to the nucleus and interacts with a protein complex that includes the transcription factors Suppressor of Hairless [Su(H)] and Mastermind (Mam) to regulate target gene expression (Bray, 2006; Kopan, 2002).
The Drosophila retina is one tissue in which reiterative N activity is evident. Ommatidial cell determination and differentiation initiates in third instar larvae in a very ordered manner. An indentation, the morphogenetic furrow (MF), sweeps across eye discs from posterior to anterior, leaving behind it the ommatidial founder cells, photoreceptor 8 precursors (R8), at
evenly spaced intervals (Wolff and Ready, 1993). N function in the MF is required for the expression of the proneural factor atonal (ato, ‘proneural enhancement’) (Jarman et al., 1994; Li and Baker, 2001). Proneural enhancement is followed by lateral inhibition defining the spacing of single ato-expressing R8 precursors (Li and Baker, 2001; Sun et al., 1998). R8 secretes the EGFR ligand Spitz (Spi), which signals to surrounding progenitors to be recruited. Newly specified photoreceptors also secrete Spi to further recruit the cells that will form the mature ommatidium. This process is counteracted by N-mediated lateral inhibition. Overactivation of EGFR signaling leads to an increased number of ommatidial cells, whereas overactivation of N signaling has the opposite effect (Cagan and Ready, 1989; Freeman, 1996; Lai and Rubin, 2001). The process of lateral inhibition by N involves transcriptional activation of various target genes, including members of the Enhancer of split complex [E(spl)-C] that act as transcriptional repressors of proneural genes (Jimenez and Ish-Horowicz, 1997; Lecourtois and Schweisguth, 1995).
In addition to its role during ommatidial assembly, N has other roles in cell fate specification posterior to the MF: it is involved in the specification of photoreceptor R4 versus R3 and R7 versus R1/R6 fates (Cooper and Bray, 1999; Cooper and Bray, 2000; del Alamo and Mlodzik, 2006; Fanto and Mlodzik, 1999; Tomlinson and Struhl, 2000), and the determination of cone cell fate (Fu and Noll, 1997). In addition, N signaling is required to control cell cycle progression during the second mitotic wave (SMW) (Baonza and Freeman, 2005), the regulation of apoptotic cell death and the specification of pigment cells during pupal development (Miller and Cagan, 1998; Rusconi et al., 2000).
Although the regulation of the core N pathway is largely understood, much remains to be known about tissue- and time-specific modulation of N responses. We address the role of the rotund(rn; m– FlyBase) gene in the context of N signaling. rn produces two different transcripts, rnand roughened eye (roe), which encode two isoforms that are members of the Krüppel family of Zn-finger transcription factors. Rn and Roe have identical C-terminal regions but differing N-C-terminal sequences caused by the use of alternative promoters. They display non-overlapping
Self-modulation of Notch signaling during ommatidial
development via the Roughened eye transcriptional
repressor
David del Alamo* and Marek Mlodzik†
The Notch (N) signaling pathway is involved in a vast number of patterning processes in all metazoans. The regulation of the core N pathway is largely understood, but little is known about fine-tuning modulatory effects. Here, we address the role of Drosophila
Krüppel-family Zn-finger transcription factor roughened eye(roe) in the context of N signaling. We demonstrate that during eye patterning, N signaling regulates the expression of roe. In turn, Roe negatively modulates the expression of target genes of N-signaling activation. In the absence of roefunction, expression of N target genes is elevated and the resulting phenotypes during patterning of the retina are similar to those of N gain-of-function scenarios. Importantly, our data show that Roe binds regulatory DNA sequences of N target genes of the E(spl)-complex both in vitro and in vivo, independently of Su(H)-DNA interaction. Thus, our data suggest that Roe acts as a transcriptional repressor in a negative-feedback loop of the N pathway.
KEY WORDS: Drosophila, Cell signaling, Feedback modulation, Transcriptional repression Development 135, 2895-2904 (2008) doi:10.1242/dev.022194
Department of Developmental and Regenerative Biology, Mount Sinai School of Medicine, 1 Gustave L. Levy Place, New York, NY 10029, USA.
*Present address: Département Biologie du Développement, Institute Pasteur, 25 rue du Docteur Roux, 75724 Paris cedex 15, France
†Author for correspondence (e-mail: marek.mlodzik@mssm.edu)
Accepted 15 June 2008
D
E
V
E
LO
P
M
E
N
expression patterns and are not functionally interchangeable (St Pierre et al., 2002). Although rnis expressed and required during wing, haltere, antenna, leg and proboscis development (del Alamo et al., 2002; St Pierre et al., 2002), roeis specifically expressed in the eye imaginal disc. No detailed functional studies have been performed but previous reports have shown that eyes mutant for roe show disrupted ommatidial patterning caused by reduced photoreceptor numbers (St Pierre et al., 2002).
Here, we analyze the function of roeduring eye development and demonstrate that N signaling modulates its own target gene activation during retinal patterning through regulation of roe expression. In the absence of roefunction, expression of N targets is elevated, causing a phenotype similar to a N gain-of-function scenario. Our results indicate that roe is required for the specification of single R8 cells and subsequent recruitment of ommatidial cells. We demonstrate that roeis expressed, under the control of N signaling, at high levels in the MF and at lower levels in progenitor cells posterior to it. Importantly, our data show that Roe can bind E(spl)-C regulatory DNA sequences both in vitro and in vivo, and, thus, Roe acts as a transcriptional repressor of N target genes.
MATERIALS AND METHODS
Drosophilastocks
Flies were grown on standard fly medium at 25°C unless otherwise stated. Strains used are described in FlyBase (www.flybase.net). rn16and rn20are
both null alleles caused by small deficiencies (St Pierre et al., 2002). Loss-of-function and ectopic expression clones were generated as previously described (Basler and Struhl, 1994; Xu and Rubin, 1993). Roe ectopic expression clones in Fig. 3D were induced onsepGAL4FRT40 UAS-roe/arm-Z FRT40 heterozygous larvae.
Immunofluorescence and histology
Larvae and pupae (~48 hours APF) were dissected and stained as described (Fanto et al., 2000). Primary antibodies used were: mouse α-Elav, rat α-Elav, mouse α-Wg, mouse α-Dl, mouse α-Nintra, mouse α-Pros (DSHB,
http://www.uiowa.edu/~dshbwww), rabbit α-βGal (Promega), mouse α
-βGal (Promega), rabbit α-Bib (Doherty et al., 1997), rabbit α-Ato (Jarman et al., 1994) and mouse α-Boss (Van Vactor et al., 1991). Secondary antibodies were from Jackson Laboratories. Adult eyes were embedded in Durcupan resin and tangential sections taken at the equatorial region. At least three independent eyes were analyzed per genotype.
Antibody production
The Roe ORF was subcloned into pQE (Qiagen) and pGEX (Amersham) for His-tagged and GST-tagged protein production, respectively. Immunization of rabbits and preparation of sera was performed by Covance. Antibody quality and specificity was evaluated by staining imaginal disc containing
rn16clones (not shown). Details are available upon request.
In situ hybridization
Two different digoxigenin probes were used (DIG RNA labeling kit, Roche). One was from the roefull-length cDNA while the other was synthesized from a ~500 bp PCR fragment of this cDNA (details available upon request). In brief, imaginal discs were fixed in 4% paraformaldehyde at 4°C, and hybridized in standard 50% formamide hybridization buffer (55°C). Probe was detected with 1:2000 Alkaline Phosphatase-coupled sheep α-DIG antibody followed by development with NBT/BCIP (Roche).
Electrophoretic mobility shift assay (EMSA)
EMSA was performed as described (Ausubel et al., 1995). Oligonucleotides were 32P-radiolabeled using Polynucleotide Kinase (NEB). The 0.3 kb probe
was amplified from the 0.5 kb mδenhancer using radiolabeled primers. The non-competitive oligonucletide is a 40 bp fragment from the
E(spl)m8regulatory region that contains one Su(H) Paired Site (Nellesen et al., 1999) with the following sequence: ATTGTGTGAGA AACTT -ACTTTCAGCTCGGTTCCCACGCCAC [Su(H) binding sites in bold].
GST::Su(H)and GST::roewere produced using pGEX vectors (see above). All binding reactions contained 10-12% glycerol, 100 ng poly dIdC and 4.5 μg BSA and were incubated for 15 minutes at 30°C before loading on a prerun gel. Gels were run at 4°C.
Chromatin immunoprecipitation (ChIP) in vivo
In vivo ChIP was performed basically as described (Delaney et al., 2006). Each experiment, including Input, IP and Mock-IP fractions, was performed using eye imaginal discs (brain included) from 200 third instar larvae of the appropriate genotypes. The input fraction was not immunoprecipitated, the IP fraction was immunoprecipitated using α-Roe antibody (1:1875) and ‘mock-IP’ was immunoprecipitated with preimmune Normal Goat Serum (1:1875). Final DNA eluate was resuspended in 20 μl for all fractions. The input fraction (1 μl) and 5 μl each of the IP and mock-IP fractions were used for PCR in 50 μl. The input PCR (5 μl) and 20 μl each of the IP and mock-IP were loaded on gel.
RESULTS
roeis required for proper cell recruitment during
ommatidial development
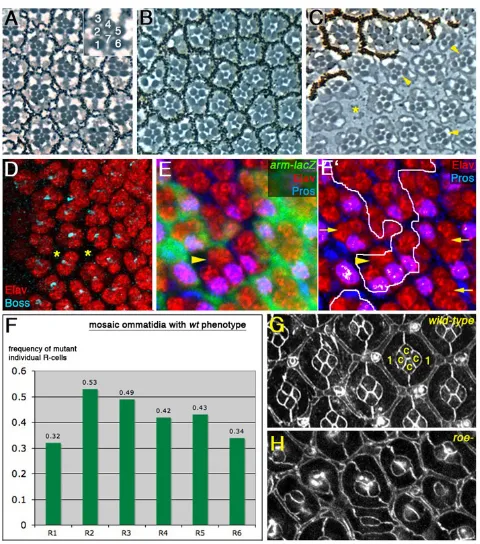
Flies mutant for the roenull condition rn16/rn20(see Materials and methods) die mostly during pupal stages, with escapers displaying rough eyes that are reduced in size (not shown) (St Pierre et al., 2002). Tangential sections of these eyes revealed defects in retinal structure (Fig. 1B), including abnormal ommatidial spacing, a reduction in number of outer photoreceptors (R1-R6) and an increase in the number of R7 cells (Fig. 1B,C; see Fig. S1 in the supplementary material). Additional defects included ommatidial fusion and loss of pigment cells. The roephenotype was evident in third instar imaginal discs, reflecting its requirement in early ommatidial patterning. Antibody staining revealed fewer Elav-positive cells per ommatidium, consistent with a reduction in photoreceptor recruitment, an occasional loss of R8, which leads to the absence of the cluster and spacing defects (asterisks in Fig. 1D, also Fig. 2A), and supernumerary R7 cells (Fig. 1E,E⬘, see below).
We next analyzed eye clones mutant for the roenull allele, rn16 (see Materials and methods). These displayed phenotypes identical to those observed in adult escaper eyes (Fig. 1C). In order to identify the requirement of roein specific photoreceptors, we analyzed phenotypically wild-type mosaic ommatidia. In white (w) clones (only affecting pigment production but not retinal development), the ratio between ommatidia that are w for one specific outer photoreceptor and total mosaic ommatidia is 0.5 for each photoreceptor type, because mosaic ommatidia include either a w(mutant) or a wild-type photoreceptor with the same probability (not shown). By contrast, if roeis required for the recruitment or specification of a photoreceptor type, the number of phenotypically normal ommatidia mutant for that photoreceptor will be significantly lower than the number of ommatidia wild type for that photoreceptor, leading to a decreased ratio. We did observe a reduced ratio for R1 (r=0.32) and R6 (r=0.34; Fig. 1F), which in both cases was statistically significant (χ2test, P<0.001). We also analyzed the number of R7s when compared with the number of outer photoreceptors per ommatidia (see Fig. S1 in the supplementary material). We detected only ommatidia with multiple R7 cells when the number of outer R1-6 photoreceptors was reduced. R1, RR1-6 and R7 are initially equivalent following their recruitment into the precluster, and Dl signaling from the R1/R6 precursors to N in R7 specifies the latter as the sole R7. Ectopic activation of N in R1 and R6 causes these cells to adopt the R7 fate (Cooper and Bray, 2000). The reduction in the number of R1-R6 cells in mutant ommatidia, and the
D
E
V
E
LO
P
M
E
N
presence of multiple R7s only in the context of decreased outer R cells, suggest that the multiple R7 phenotype is due to a transformation of R1 and R6 into R7. This is further supported by the fact that Roe expression is enriched in R1 and R6 precursors prior to recruitment (see below).
These results were confirmed by the analysis of rn16clones in imaginal discs using markers for different cell types. Three phenotypic defects were observed in mutant tissue: (1) reduction of the number of R8 cells and thus ommatidial preclusters (Fig. 2A,A⬘; α-Ato staining); (2) reduced number of recruited photoreceptors per ommatidium (Fig. 1D); and (3) multiple R7 cells, as detected in adult sections and by the molecular R7 marker Prospero (Fig. 1B,C,E,E⬘; and not shown).
In addition, we analyzed the cell number and organization of cone and pigment cells in pupal eyes. α-Arm staining (see Materials and methods), which delineates cell membranes, revealed under-recruitment of cone cells (Fig. 1G,H). This was confirmed by imaginal disc staining with the cone cell marker Cut (not shown). This analysis also revealed a reduction of secondary and tertiary pigment cells and defects in interommatidial bristle specification (Fig. 1G,H).
Taken together, our results indicate that roeis required for proper ommatidial cell recruitment at several stages of eye development. Strikingly, the roephenotype resembles that of a mild overactivation of N signaling during eye patterning.
N target gene expression is elevated inroeclones
Based on this phenotype, we explored the possibility that roeis acting as a negative regulator of N activity. We thus analyzed the expression of N target genes in roe16clones.
Ato expression (α-Ato) is first detected as a continuous stripe in the MF and subsequently resolves into single cells (R8) (Jarman et al., 1994; Sun et al., 1998). In roemutant clones, we observed occasional loss of Ato staining in single cells, indicative of the failure to specify R8 photoreceptors (arrows in Fig. 2A,A⬘; compare with arrowheads in same panels). There was no significant change in Ato expression during proneural enhancement within the MF (Fig. 2A,A⬘). To confirm a role of roein the regulation of ato expression during lateral inhibition, we analyzed the expression of the lacZ-reporter ato5⬘F9.3-Z (ato-lacZin Fig. 2B,B⬘). This reporter is repressed by N signaling through lateral inhibition at this stage (Sun et al., 1998). In roe clones, ato5⬘F9.3-Z expression was completely lost, indicating that N activity on this reporter was increased in the roemutant background (Fig. 2B,B⬘).
We next analyzed the expression of lacZreporter lines that are positively regulated by N signaling during eye patterning (Cooper et al., 2000). E(spl)mβ1.5-Z (mβ-lacZ in Fig. 2C,C⬘) shows a general expression posterior to the furrow and its expression was increased inside roe- clones (Fig. 2C,C⬘). E(spl)mγ1.1-Z(mγ-lacZin Fig. 2D,D⬘) is expressed in a more restricted manner posterior to the MF. In roe–clones, this expression was also upregulated (Fig. 2D,D⬘).
In contrast to these N targets, we did not observe changes in expression of the core N signaling component Dl or N itself (Fig. 2E-F), suggesting that roe might act downstream of N. Taken together, these data suggest that Roe attenuates N signaling.
Roe represses N target gene expression
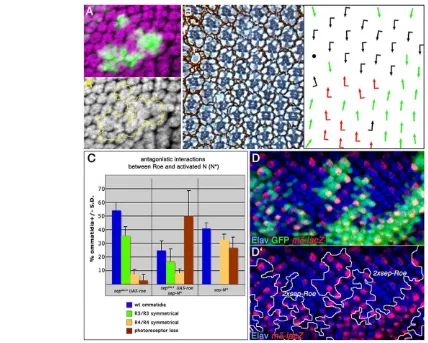
[image:3.612.56.296.58.330.2]The loss-of-function data suggested that roeantagonizes N signaling activity (see above). We thus wanted to confirm this hypothesis with gain-of-function studies. First, clonal overexpression of roe in developing ommatidia produced a phenotype similar to that of N loss-of-function clones: the recruitment of supernumerary photoreceptors to the cluster (Fig. 3A,A⬘). This is in agreement with our loss-of-function data (reduced number of R-cells) and consistent with Roe being a negative regulator of N signaling. Expression of N signaling target genes was difficult to analyze owing to the recruitment of extra photoreceptors, which show reduced levels of Fig. 1. roeloss-of-function causes defective ommatidial cell
recruitment. (A-C) Tangential eye sections with anterior leftwards and dorsal upwards. (A) Wild-type adult eye. Outer photoreceptors (R1-6) and the inner R7 are found in every ommatidium (R8 is in a different plane) surrounded by pigment cells (rhabdomeres are numbered in inset). (B) rn16/rn20mutant eye with most ommatidia containing fewer
R-cells and irregular spacing. (C) Mosaic eye bearing rn16clones
(absence of pigment). roe–ommatidia show a reduced number of R1-R6 cells and often multiple R7s (examples indicated with yellow arrowheads; see Fig. S1 in the supplementary material). There are also defects in spacing (example marked by asterisk). (D-E⬘) Areas posterior to the MF in 3rd instar eye discs (anterior is leftwards) stained for α-Elav (red, marking all R-cells), α-Boss (turquoise in D, marking R8) and α -Pros (blue in E,E⬘; marking R7). (D) rn16mutant disc; (E,E⬘) mosaic discs
with wild-type tissue marked by arm-lacZ(green in E, outlined in E⬘). Fewer R cells are recruited per ommatidium. Spacing defects result from the presence of fewer R8 founder cells (examples highlighted by asterisks, see also Fig. 2A,A⬘). α-Pros labels R7 cells (and cone cells in a different plane, which are Elav negative). The multiple R7 phenotype is observed inside mutant tissue (example indicated by yellow arrowhead; wild-type clusters with a single R7 are indicated by arrows). Only the nuclei of R7, R3 and R4 are visible in this focal plane in most clusters. (F) Graph showing the frequency of individual mutant R cells in mosaic ommatidia with a wild-type wtphenotype (random frequency reflecting no requirement is 0.5; n=152 in nine eyes analyzed). R1 and R6 show a significantly reduced frequency, indicating a functional requirement of roein these cells. (G,H) Pupal retinae stained with α-Arm, which delineates cell membranes: (G) wild type (‘c’ marking cone cells and ’1’ primary pigment cells in example) and (H) rn16are shown. The
quasi-crystalline lattice of wild-type pupal eyes is severely disturbed in roe mutants. Cone cell, secondary and tertiary pigment cell, and
interommatidial bristle numbers are reduced compared with wild type.
D
E
V
E
LO
P
M
E
N
N activity per se. To circumvent this problem, we used the well-defined N function in R3/R4 specification. Following the recruitment of the R3/R4 precursor pair, elevated Fz/PCP signaling in R3 leads to higher expression levels of Dland neuralizedin R3. This causes a directional activation of N in R4, which then specifies R4 fate. N activity can be monitored by the expression of the reporter line E(spl)mδ0.5-Z(Cooper and Bray, 1999; del Alamo and Mlodzik, 2006; Fanto and Mlodzik, 1999). We overexpressed Roe using a sevenless(sev)-derived driver line, sepGAL4, which is active at high levels in the R3/R4 precursor pair (Fanto et al., 2000), and observed a large number of R3/R3 symmetrical ommatidia (35.4±6.7%, Fig. 3B,C), characteristic of a failure of N activation in R4 precursors. Consequently, imaginal disc staining of the N target reporter E(spl)mδ0.5-Zshowed reduced or absent expression in R4 (not shown). To further analyze this effect, we induced clones of cells expressing two copies of sepGAL4, UAS-Roeand confirmed the reduction in expression of E(spl)mδ0.5-Z(Fig. 3D,D⬘).
In contrast to Roe overexpression, constitutive activation of N in both cells of the R3/R4 pair (sep-NΔECD, see Materials and methods), causes the opposite phenotype, leading to R4/R4 symmetrical clusters (32.6±4.3%, Fig. 3C and not shown). Notably, this phenotype was suppressed by simultaneous co-expression of Roe and NΔECD (Fig. 3C and not shown), resulting in 16.9±9.1% R3/R3 symmetrical clusters and 8±3.5% R4/R4-type clusters (Fig. 3C). The loss-of-R-cell phenotype of sep-NΔECDwas increased, although this phenotype is not likely to be produced by defective cell recruitment as sepGAL4 is expressed at high levels only in photoreceptors after recruitment.
To confirm the ability of Roe to negatively modulate N signaling, we turned to a heterologous system, the leg imaginal disc, where roe is not expressed endogenously (not shown) (St Pierre et al., 2002).
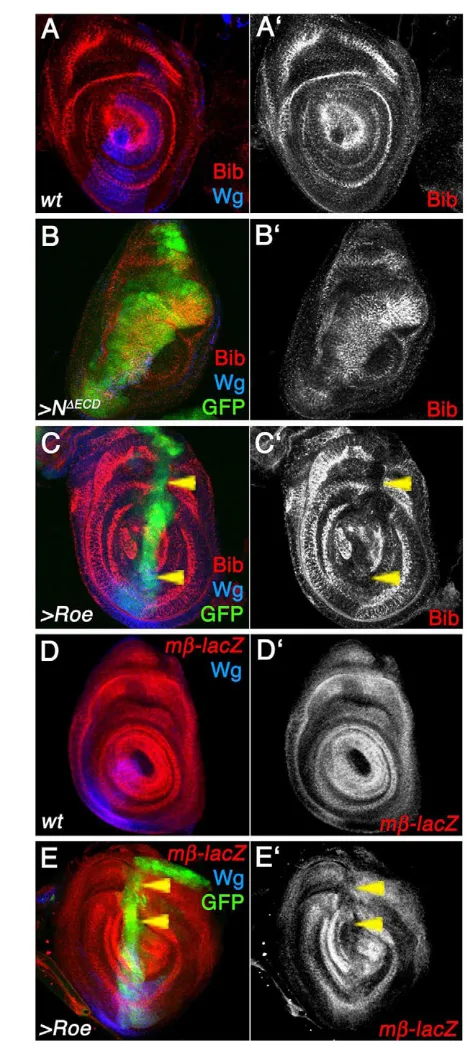
In leg imaginal discs, the big brain (bib) gene is expressed in concentric rings, which constitute the presumptive leg joint cells, in response to N signaling (Fig. 4E) (de Celis et al., 1998). Activation of N within the dpp expression stripe (dppGAL4, UAS-NΔECD) ectopically activates Bib expression (Fig. 4B,B⬘). By contrast, ectopic expression of Roe in the same domain, led to a repression of endogenous Bib expression (Fig. 4C,C⬘). In all cases, expression of wingless (wg) in a ventral-anterior wedge, independent of N signaling, was not affected (blue in Fig. 4C; not shown), indicating that the effect was specific. Similarly, the N-signaling reporter/target E(spl)mβ1.5-Z(mβ-lacZ in Fig. 4D-E) is expressed in concentric rings coincident with those of bib(Fig. 4D,D⬘). Expression of Roe within the dpp-domain (dppGAL4, green in Fig. 4E) represses E(spl)mβ1.5-Z(Fig. 4E,E⬘). By contrast, as with bib, expression of activated N under dppGAL4control causes ectopic upregulation of E(spl)mβ1.5-Z(not shown).
In summary, these data indicate that: (1) roe overexpression causes similar phenotypes to Nloss of function; and (2) that roeis a general inhibitor of Nsignaling target gene expression only limited by its expression pattern.
roeexpression is controlled by N signaling
[image:4.612.56.511.58.291.2]Previous studies have suggested that roeis expressed at high levels in the MF (St Pierre et al., 2002). Using two different RNA probes (see Materials and methods) we were able to detect roeexpression not only in the MF but also posterior to it (Fig. 5A; not shown). In order to define roeexpression in detail, we developed a polyclonal antibody (see Materials and methods). In agreement with our in situ hybridization experiments, antibody staining revealed that Roe is expressed at high levels in the MF, and at lower levels in Fig. 2. roeloss-of-function affects the expression of N-signaling targets. Panels show third instar mosaic imaginal discs bearing rn16clones,
marked by absence of arm-lacZ(red in A,E,F) or Ubi-GFP(red in B-D) and outlined in white in right panels. α-Elav is green in B-E. (A,A⬘) α-Ato (blue) is detected as a continuous band at the MF and resolves into individual cells (R8) posterior to it. α-Ato is reduced in single cells inside the mutant clone (arrows) when compared with surrounding wild-type tissue (arrowheads). (B,B⬘) The Ato reporter line ato5⬘F9.3-Z(blue), which is repressed by N-signaling during R8 lateral inhibition, is not detected inside mutant tissue. (C,C⬘) The E(spl)mβ1.5-Z reporter line (blue), which is regulated by N signaling, shows higher expression levels inside mutant clones when compared with surrounding wild-type cells. (D,D⬘) The E(spl)mγ1.1-Zreporter (blue), which is activated by N signaling, is expressed in isolated cells posterior to the MF. Inside mutant tissue, more positive cells are observed when compared with surrounding wild-type tissue. (E-F⬘) Neither N (E,E⬘, blue and monochrome, detected by α-Nintra) nor Dl expression levels (F,F⬘, turquoise and monochrome) are affected inside mutant tissue.
D
E
V
E
LO
P
M
E
N
interommatidial progenitor cells posterior to the furrow (Fig. 5B,B⬘, note largely non-overlapping staining between Elav- and Roe-positive cells). In accordance with its predicted Zn-finger transcription factor nature (St Pierre et al., 2002), Roe was detected only in the nuclei (Fig. 5B,B⬘). Furthermore, elevated Roe expression was observed in the R1/R6 precursor pair (identified by their position with respect to the whole cluster) at the onset of Elav expression (Fig. 5C). The identity of R1 and R6 was further confirmed by co-staining with a sevenless reporter line (sevGAL4, UAS GFP), which is expressed in R1, R6 and R7 prior to recruitment (Fig. 5D,D⬘). These observations support our previous observation of a specific requirement in the R1/R6 pair (Fig. 1F; see above).
The Roe expression pattern is strikingly similar to that of N activity posterior to the MF. Thus, we analyzed the effect of N signaling on Roe expression. First, we tested loss-of-function clones of N (N55e11) and observed a clear cell-autonomous
reduction of Roe expression inside mutant tissue, both at the MF and posterior to it (Fig. 5E,E⬘). A similar effect was seen with DlRevF10clones (not shown). Conversely, ectopic activation of N signaling (NΔECD; see Materials and methods) caused ectopic upregulation of Roe close to the MF and elevated expression posterior to it (Fig. 5F,F⬘; overexpression of Dl caused the same effects, not shown). Expression analysis at the MF is complex, because N can induce ectopic MFs and photoreceptor differentiation (with subsequent reduction of roe expression levels) through activation of dpp expression during the reincarnation process of MF movement (Fig. 5F,F⬘; data not shown) (Kumar and Moses, 2001).
[image:5.612.53.481.57.400.2]In contrast to N, loss-of-function of EGFR signaling generated through clonal expression of UAS-DERDNor UAS-aos(Freeman, 1994; Freeman, 1996), or loss-of-function of the Hh and Dpp signaling pathways did not affect roeexpression (not shown). Fig. 3. Overexpression of Roe phenocopies Nloss-of-function phenotypes and represses N target genes during eye development. (A,A⬘)roeoverexpression clones (marked by GFP in green, outlined in A⬘) in third instar eye disc cause recruitment of supernumerary photoreceptors (anti-Elav, magenta), similar to N pathway loss-of-function clones (compare with Fig. 5E). (B) Tangential section of adult eye of the genotype sepGAL4,
UAS-roewith schematic shown on the right (black and red arrows represent the two chiral forms; green arrows represent symmetrical clusters; black dot shows loss of R cells). Increased levels of Roe expression in R3/R4 precursors often cause the formation of symmetrical R3/R3 type ommatidia (some R4/R4 type are also observed; quantified in C). (C) Quantification of the phenotypes of sev-driven expression of roe, NΔECDand together.
Ectopic expression of NΔECDcauses a high number of R4/R4-type symmetrical ommatidia and R-cell loss, while Roe causes mostly R3/R3-type and occasional R-cell loss. The NΔECDphenotype is antagonized by Roe co-expression and a reversion to a significant percentage of R3/R3-type ommatidia typical of Roe overexpression is observed (the ‘loss of R-cell’ phenotype is enhanced, see text for details). Total number of ommatidia scored was 391 (roe), 545 (NΔECD) and 329 (roe +NΔECD) with at least three eyes per genotype analyzed. (D,D⬘) Third instar eye disc bearing clones of cells expressing
two copies of sepGAL4, UAS-roe(marked by absence of Ubi-GFPin green, see Materials and methods). Expression of the R4-specific N reporter
mδ-lacZ(red) is largely suppressed inside the Roe-overexpressing tissue. Cells outside the clone can be either wild type or have one copy of sepGAL4,
UAS-roe, which can also suppress mδ-lacZto a lower extent (not shown). α-Elav is in blue.
D
E
V
E
LO
P
M
E
N
Taken together, our results indicate that Roe is localized to the nucleus of cells at the MF and in progenitor cells and that its expression is (at least partially) under N-signaling control. Our data suggest that Roe is part of a negative-feedback loop modulating N signaling outcome during ommatidial patterning.
Roe binds to E(spl)-C regulatory sequences
roeencodes a putative Zn-finger transcription factor (St Pierre et al., 2002) and our antibody staining showed nuclear subcellular localization of the protein, suggesting that Roe might act by binding to DNA to regulate the transcription of N targets. To test this hypothesis, we performed Electrophoretic mobility shift assays (EMSAs). As a probe, we used a 300 bp DNA fragment included in the ~500 bp mδ0.5 enhancer sequence (Cooper et al., 2000) that contains the Su(H) paired sites (SPS) (Nellesen et al., 1999) (see Fig. 6A for schematic; see also Materials and methods). Our previous experiments showed that the mδ0.5 sequence was responsive to Roe overexpression in vivo (e.g. Fig. 3D).
We used GST::Su(H) as a positive control and we detected Su(H) binding to the probe (Fig. 6B, lanes 1-2). We were also able to detect a shift with GST::Roe (Fig. 6B, lanes 5-6). Interestingly, the simultaneous presence of both proteins lead to the same shifts that were observed with each individual protein (Fig. 6B, lane 3), indicating that Su(H) and Roe bind DNA independently of each other. GST-pull down experiments failed to detect any interaction between Su(H) and Roe (not shown), further confirming this conclusion. The binding was successfully out-competed by an excess of cold probe in both cases (Fig. 6B, lanes 4 and 7), indicating a specific interaction. In an attempt to further define Roe binding sites in this probe, we performed EMSA with nine overlapping 60 bp oligonucleotides covering the whole 300 bp fragment (P1-P9 in Fig. 6A; see Fig. S2 in the supplementary material). We detected binding of GST::Roe to all oligonucleotides. The GST::Roe binding was specific, as it was out-competed by an excess of the respective cold oligonucleotides but not by an equal excess of an unrelated oligonucleotide (see Fig. S2 in the supplementary material; see also Materials and methods). These data suggest that multiple specific Roe-binding sites exist in E(spl)mδregulatory sequences.
In order to prove that Roe binds to regulatory DNA sequences in vivo during normal eye development, we performed chromatin immunoprecipitation assays (ChIPs, see Materials and methods) from in vivo samples. In this assay, we were able to recover the 300 bp fragment from DNA immunoprecipitated using the anti-Roe antibody on eye imaginal disc homogenates (Fig. 6C). By contrast, when preimmune serum was used (mock-ChIP), no specific bands were immunoprecipitated (Fig. 6C, right lane). As a specificity control, we performed ChIP with anti-Roe with sequences from the unrelated antimicrobial gene attacinA(Delaney et al., 2006) and did not detect binding, confirming that the Roe interaction with E(spl)mδregulatory DNA is specific (Fig. 6C, lower panel). To further define the specificity of the interaction, we performed ChIP with imaginal discs from roenull (rn16) larvae and did not detect an interaction, either with the E(spl)mδregulatory DNA sequences or with the AttA promoter (Fig. 6D, upper and lower panels, respectively), again indicating that the ChIP interaction is specific for Roe.
[image:6.612.50.285.55.578.2]In summary, our results indicate that Roe binds to E(spl)mδ regulatory DNA sequences both in vitro and in vivo independently of Su(H), providing a molecular mechanism for Roe action in repressing N-signaling target genes.
Fig. 4. Roe represses N signaling targets during leg disc patterning. Panels show leg discs, with anterior leftwards and dorsal upwards. Wg (blue, negative control) is expressed in a ventral wedge at the A/P boundary in a N-independent manner. (A,A⬘) Bib (red, monochrome in A⬘) is expressed in concentric rings in leg discs, corresponding to the presumptive leg segment joints defined by N signaling. (B,B⬘) dppGAL4
-driven NΔECDectopic expression (marked by GFP, green) causes a
cell-autonomous expansion of Bib expression inside the dppGAL4domain.
(C,C⬘) Ectopic expression of Roe under the control of dppGAL4(green)
causes cell-autonomous repression (arrowheads) of Bib (red,
monochrome in C’). (D,D⬘) The E(spl)mβ1.5-Z reporter line (mβ-lacZred and monochrome) is regulated by N signaling and expressed in concentric rings corresponding to presumptive cells of the leg joints. (E,E⬘) dppGAL4
-driven ectopic expression of Roe (green) causes cell-autonomous
repression (arrowheads) of mβ-lacZ(red, monochrome in E⬘).
D
E
V
E
LO
P
M
E
N
DISCUSSION
Lateral inhibition mediated by N signaling is used reiteratively during Drosophiladevelopment. Lateral inhibition normally occurs in groups of initially equivalent cells to limit the number of those that acquire a certain fate by repression of cell fate determinants via the E(spl) complex genes (reviewed by Bray, 2006; Jimenez and Ish-Horowicz, 1997; Lecourtois and Schweisguth, 1995). For example, N acts in proneural clusters to ensure that the proper number of sensory organ precursors (SOPs) is specified (reviewed by Simpson, 1997), or during eye development to restrict the number of progenitor cells that are recruited into the ommatidial cluster (Cagan and Ready, 1989).
During Drosophilaeye development, following the specification of the ommatidial founder cell R8, the remaining cells of the ommatidium are sequentially recruited through a two-step mechanism: first, Spitz (Spi) signals to the DrosophilaEGF receptor in surrounding progenitor cells and induces recruitment (Freeman, 1996) and, second, Dl/N-signaling represses the recruitment to ensure that only the appropriate number of cells join the cluster (Cagan and Ready, 1989; Fortini et al., 1993). N signaling is not only involved in the regulation of ommatidial assembly during eye development, but several other processes as well (see Introduction). Presumably, different N targets or different levels of N activity, or both, together with integration of signals from other pathways account for the large variety of cell responses. How this is achieved is largely unknown (reviewed by Bray, 2006; Doroquez and Rebay, 2006). Here, we have identified and molecularly defined the role of the Roe transcription factor as a negative modulator of N target gene expression, functioning in a negative-feedback loop of N signaling itself.
roughened eye: a tissue-specific modulator of N signaling
Loss-of-function phenotypes of roephenocopy mild N gain-of-function defects during ommatidial patterning: failure to specify R8 cells, under-recruitment of R-cells and specification of multiple R7 cells at the expense of R1/R6. In accordance, we identify several N targets that are upregulated in roeloss-of-function clones, including E(spl)-Creporter lines. Pupal eye analyses reveal that in addition to photoreceptors other ommatidial cells are affected, including cone cells, secondary and tertiary pigment cells, and interommatidial bristles. Dl and Spi are expressed in R cells activating N and Egfr pathways, respectively, in surrounding progenitor cells, where they cooperate with the transcription factor Lozenge to specify the cone cell fate (Daga et al., 1996; Flores et al., 2000). roeloss-of-function can induce higher levels of N target genes that lead to a reduction in the number of R cells. As R cells produce the signals that specify cone cells, it is likely that fewer R cells produce, in turn, fewer cone cells.
Reciprocal to the loss-of-function defects, Roe overexpression (Fig. 3) leads to repression of N target genes resulting in the recruitment of extra photoreceptors. Similarly, we demonstrate that Roe is able to repress the N-signaling dependent R4 fate. Importantly, the repression effect of Roe can be antagonized by simultaneous co-expression of an activated form of N, suggesting that a fine balance between the activity of N-signaling and Roe is employed in many of these cell fate decisions. Taken together, these results indicate that roeis a negative modulator of N target gene expression. However, the effects of Roe are only partial when compared with N. For example, full activation of N leads to complete abolishment of photoreceptor recruitment (see Fig. 5F, for example), whereas roe-null (rn16) clones show a reduced number of photoreceptors per ommatidium (Fig. 1D,E). These data suggest that Roe functions as a negative-feedback modulator rather than as a core component of the N pathway.
[image:7.612.52.299.60.338.2]Importantly, loss- and gain-of-function experiments with N and Dlshow that roeexpression is under the control of N signaling, defining a new self-regulatory feedback loop. Sequence analysis demonstrates that four high-affinity Su(H)-binding sites exist in the introns of the rn/roegene and one of them lies within 1 kb of the roetranscriptional start site (Berman et al., 2004). We thus propose a model where N self-modulates its activity at the level of target gene expression by activating roeexpression, which, in turn, represses expression of N target genes during ommatidial patterning.
Fig. 5. roeexpression is under the control of N activity. Panels show third instar eye disc regions around the MF; anterior is upwards. (A) In situ hybridization staining: roeis expressed in the MF (yellow arrowhead) and posterior to it. (B,B⬘) α-Roe staining (turquoise, monochrome in B⬘). Roe is a nuclear protein detected at high levels in the MF and at lower levels posterior to it. Roe is higher in
interommatidial (progenitor) cells and shows only low levels in photoreceptors (Elav, red). (C) Third instar eye disc posterior to the MF showing α-Roe (turquoise) and α-Elav (red) staining. Note higher levels of Roe in R1/R6 precursors (examples marked with asterisks). (D,D⬘) High magnification of an ommatidial precluster showing α-Elav (red), α-Roe (blue) and sevGAL4UAS-GFP(green) staining (D) and schematic
cartoon (D⬘). Note that sevGAL4and Roe are co-expressed in R1/R6
precursors prior to Elav detection. (E,E⬘) N loss-of-function clones (marked by absence of Ubi-GFP, green) cause reduction in Roe expression levels (blue, monochrome in E⬘), both within the MF (compare to B’) and posterior to it. Photoreceptors are marked with α -Elav (red in E). (F-F⬙) Clones expressing NΔECD(marked by co-expression
of GFP, green, outlined in F⬘) cause ectopic MF initiation in cells anterior to endogenous MF and ectopic cell-autonomous upregulation of Roe (blue, monochrome in F⬘) posterior to MF. Clones are as expected associated with loss of photoreceptors (note reduction in α-Elav, red and monochrome in F⬙). Close to and within the MF, NΔECD-expressing
clones cause a precocious initiation of the MF state with associated non-autonomous Roe activation (see also Results).
D
E
V
E
LO
P
M
E
N
The transcriptional repressor Roe functions in parallel to Su(H) action
Our results demonstrate that Roe can bind directly to DNA both in vitro and in vivo. This binding is independent of Su(H), as no difference in DNA-binding ability of either of these transcription factors was detected in the presence of the other. However, the DNA-binding specificity of Roe remains unclear. EMSA analysis shows that multiple binding sites exist in E(spl)mδ regulatory sequences. Sequence comparison among the different fragments used (Fig. S2 in the supplementary material) did not identify a consensus binding site. This suggests that Roe displays low binding specificity to DNA in vitro.Thus, higher specificity as evident in vivo must be achieved through interactions of Roe with other DNA-binding transcription factors and/or transcriptional co-factors.
Current models of N activation propose that the Nintradomain translocates to the nucleus where it forms a complex with the DNA-binding transcription factor Su(H) and the co-activator Mam (Wilson and Kovall, 2006). This complex further recruits other co-activators, like the histone acetyl-transferase CBP/p300 or the spliceosome component SKIP (Wallberg et al., 2002; Zhou et al., 2000). In the absence of N signaling, Su(H) itself acts as a transcriptional repressor (Castro et al., 2005), interacting with the adaptor protein Hairless, which recruits co-repressors to the complex (reviewed by Bray, 2006; Kao et al., 1998; Nagel et al., 2005). Thus,
the translocation of Nintrato the nucleus and its interaction with Su(H) switches a transcriptional repressor complex to an activator complex.
How does Roe fit into this scenario? First, activation of N-signaling upregulates Roe expression. Roe in turn binds to the regulatory sequences of specific target genes in the vicinity of the Su(H) complex and antagonizes its activity. Through this mechanism, N signaling cannot only turn an off-state into an on-state [via Su(H)], but can also regulate the activity levels of this switch (via Roe). In this model, a target gene is activated less in the presence of Roe than it is in its absence, although it is generally on (Fig. 6E). Roe could also affect the turnover of Su(H) on the DNA. Recent studies have shown that the nuclear translocation of Nintranot only causes the switch from a repression to an activation complex, but also affects the Su(H) binding turnover at the DNA. In the presence of Nintra, Su(H) occupancy of target binding sites is higher than in its absence, suggesting that Su(H) binds dynamically to DNA (Krejci and Bray, 2007). Further studies will be required to precisely assess the role of Roe in N target repression.
[image:8.612.251.563.61.374.2]The fact that Roe can bind to regulatory sequences independently of the core N-transcriptional machinery raises the interesting possibility of differential target-specific modulation. This mode of regulation is particularly useful in the fine control of multiple cellular outcomes elicited by a single signaling pathway, as is the Fig. 6. Roe binds E(spl)-Cregulatory DNA
sequences both in vitro and in vivo. (A) Schematic presentation of the relative position of the 0.3 kb fragment used as a probe for EMSA or amplified in in vivo chromatin immunoprecipitation (ChIP) assays with respect to the E(spl)mδgene. Coordinates are based upon ‘Release 5’ of the Drosophilagenome sequence (BDGP, http://www.fruitfly.org/). 60 bp fragments denoted as P1-P9 correspond to the probes used in EMSAs described in Fig. S2 in the supplementary material. (B) Representative EMSA experiment using the 0.3 kb fragment (A) as a probe. Lanes are as follows: (1) Free probe (FP) +2.5 μg Su(H), (2) FP + 5 μg Su(H), (3) FP+2.5 μg Su(H) + 2.5 μg Roe, (4) FP + 2.5 μg Su(H) + 2.5 μg Roe + excess cold probe (ECP) as competitor, (5) FP + 2.5 μg Roe, (6) FP + 5 μg Roe, and (7) FP + 2.5 μg Roe + ECP. Shifts caused by Su(H) and Roe (lanes 1,2 and 5,6, respectively) are unaffected when both proteins are simultaneously present (lane 3). (C) In vivo ChIP from a wild-type eye imaginal discs. A band corresponding to the 0.3 kb fragment is amplified when the sample is
immunoprecipitated with anti-Roe antibody (Roe-ChIP lane, compare with INPUT lane) but not when preimmune NGS was used for IP (mock-ChIP lane). As a control for specificity, anti-Roe did not
co-immunoprecipitate DNA from the AttacinA(AttA) promoter. (D) In vivo ChIP from homozygous roenull (rn16) eye discs. No specific ChIP band was amplified
when the sample was immunoprecipitated either with anti-Roe antibody or preimmune NGS (mock-IP). Same results were obtained with DNA from the AttA promoter. In both C and D, weak bands can be
observed occasionally, owing to nonspecific binding of DNA to the resin used during the IP process. (E) Model for Roe action on N targets. In the absence of N-signaling activity, N target genes are repressed by DNA-bound Su(H) together with transcriptional co-repressors. Upon N activation, Nintratranslocates to the nucleus and, together with Mam and other transcriptional co-activators (not shown), binds to Su(H) to turn ON the expression of target genes. We propose a third scenario in which N self-modulates its transcriptional activity by upregulating Roe. Roe binds to regulatory sequences of N target genes independently of Su(H), leading to an attenuated transcriptional activity.
D
E
V
E
LO
P
M
E
N
case of N in the developing eye. In this regard, in a roe loss-of-function background, we observed upregulated expression of the N-target genes of the E(spl)-C, involved in ommatidial assembly; by contrast, we did not observe any effect on the N-target CyclinA, which is required in interommatidial cells to control proliferation (not shown) (Baonza and Freeman, 2005).
Rn and Roe: one gene with two isoforms and two functions
The Roe and Rn isoforms of the rngene share the last 450 C-terminal residues, including five of the six Zn fingers. However, rescue experiments performed in previous studies show they are not functionally interchangeable (St Pierre et al., 2002). Studies on the function of the Rn isoform during wing development have shown that it is required for hinge development, without any relationship to N signaling regulation (del Alamo et al., 2002). Our data and these previous studies indicate that Roe is a transcriptional repressor, whereas Rn is likely to function as an activator (Terriente Felix et al., 2007). The rngene is thus a notable example of how functional diversification can be achieved during evolution by domain swapping among proteins through alternative splicing and promoter usage.
We are grateful to Spyros Artavanis-Tsakonas, Sarah Bray, Fernando J. Diaz-Benjumea, Gary Struhl and the Bloomington Stock Center for fly stocks and reagents. We thank Sophy Okello for embryo injections, William Gault, Robert Krauss, Jaskirat Singh and Jun Wu for comments on the manuscript, and all members of the Mlodzik laboratory for input and support. This work was supported by NIH/NEI grant RO1 EY13256 to M.M. and a fellowship from the Spanish Ministry of Education and Science to D.D.A.
Supplementary material
Supplementary material for this article is available at http://dev.biologists.org/cgi/content/full/135/17/2895/DC1
References
Artavanis-Tsakonas, S., Rand, M. D. and Lake, R. J.(1999). Notch signaling: cell fate control and signal integration in development. Science284, 770-776. Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G.,
Smith, J. A. and Struhl, K.(1995). Current Protocols in Molecular Biology. New York, NY: John Wiley and Sons.
Baonza, A. and Freeman, M.(2005). Control of cell proliferation in the Drosophila eye by Notch signaling. Dev. Cell8, 529-539.
Basler, K. and Struhl, G.(1994). Compartment boundaries and the control of Drosophila limb pattern by Hedgehog protein. Nature368, 208-214. Berman, B. P., Pfeiffer, B. D., Laverty, T. R., Salzberg, S. L., Rubin, G. M.,
Eisen, M. B. and Celniker, S. E.(2004). Computational identification of developmental enhancers: conservation and function of transcription factor binding-site clusters in Drosophila melanogaster and Drosophila pseudoobscura.
Genome Biol. 5, R61.
Bray, S. J.(2006). Notch signalling: a simple pathway becomes complex.Nat. Rev Mol. Cell. Biol. 7, 678-689.
Cagan, R. L. and Ready, D. F.(1989). Notch is required for successive cell decisions in the developing Drosophila retina. Genes Dev.3, 1099-1112. Castro, B., Barolo, S., Bailey, A. M. and Posakony, J. W.(2005). Lateral
inhibition in proneural clusters: cis-regulatory logic and default repression by Suppressor of Hairless. Development132, 3333-3344.
Cooper, M. T. and Bray, S. J.(1999). Frizzled regulation of Notch signalling polarizes cell fate in the Drosophila eye. Nature397, 526-530.
Cooper, M. T. and Bray, S. J.(2000). R7 photoreceptor specification requires Notch activity.Curr. Biol. 10, 1507-1510.
Cooper, M. T., Tyler, D. M., Furriols, M., Chalkiadaki, A., Delidakis, C. and Bray, S.(2000). Spatially restricted factors cooperate with notch in the regulation of Enhancer of split genes. Dev. Biol. 221, 390-403.
Daga, A., Karlovich, C. A., Dumstrei, K. and Barnejee, U.(1996). Patterning of cells in the Drosophila eye by Lozenge, which shares homologous domains with AML1. Genes Dev.10, 1194-1205.
de Celis, J. F., Tyler, D. M., de Celis, J. and Bray, S. J.(1998). Notch signalling mediates segmentation of the Drosophila leg. Development125, 4617-4626. del Alamo, D. and Mlodzik, M.(2006). Frizzled/PCP-dependent asymmetric
neuralized expression determines R3/R4 fates in the Drosophila eye. Dev. Cell11, 887-894.
del Alamo, D., Terriente, J., Galindo, M. I., Couso, J. P. and Diaz-Benjumea, F. J.(2002). Different mechanisms initiate and maintain wingless expression in the Drosophila wing hinge. Development129, 3995-4004.
Delaney, J. R., Stoven, S., Uvell, H., Anderson, K. V., Engstrom, Y. and Mlodzik, M.(2006). Cooperative control of Drosophila immune responses by the JNK and NF-kappaB signaling pathways. EMBO J.25, 3068-3077.
Doherty, D., Jan, L. Y. and Jan, Y. N.(1997). The Drosophila neurogenic gene big brain, which encodes a membrane-associated protein, acts cell autonomously and can act synergistically with Notch and Delta. Development124, 3881-3893. Doroquez, D. B. and Rebay, I.(2006). Signal integration during development:
mechanisms of EGFR and Notch pathway function and cross-talk. Crit. Rev. Biochem. Mol. Biol. 41, 339-385.
Fanto, M. and Mlodzik, M.(1999). Asymmetric Notch activation specifies photoreceptors R3 and R4 and planar polarity in the Drosophila eye. Nature397, 523-526.
Fanto, M., Weber, U., Strutt, D. I. and Mlodzik, M.(2000). Nuclear signaling by Rac and Rho GTPases is required in the establishment of epithelial planar polarity in the Drosophila eye.Curr. Biol. 10, 979-988.
Fleming, R. J., Purcell, K. and Artavanis-Tsakonas, S.(1997). The NOTCH receptor and its ligands. Trends Cell. Biol. 7, 437-441.
Flores, G. V., Duan, H., Yan, H., Nagaraj, R., Fu, W., Zou, Y., Noll, M. and Barnejee, U.(2000). Combinatorial signaling in the specification of unique cell fates. Cell103, 75-85.
Fortini, M. E., Rebay, I., Caron, L. A. and Artavanis-Tsakonas, S.(1993). An activated Notch receptor blocks cell-fate commitment in the developing Drosophila eye. Nature365, 555-557.
Freeman, M.(1994). Misexpression of the Drosophila argos gene, a secreted regulator of cell determination. Development120, 2297-2304.
Freeman, M.(1996). Reiterative use of the EGF receptor triggers differentiation of all cell types in the Drosophila eye. Cell87, 651-660.
Fu, W. and Noll, M.(1997). The Pax2 homolog sparkling is required for development of cone and pigment cells in the Drosophila eye. Genes Dev.11, 2066-2078.
Jarman, A. P., Grell, E. H., Ackerman, L., Jan, L. Y. and Jan, Y. N.(1994). Atonal is the proneural gene for Drosophila photoreceptors. Nature369, 398-400.
Jimenez, G. and Ish-Horowicz, D.(1997). A chimeric enhancer-of-split transcriptional activator drives neural development and achaete-scute expression. Mol. Cell. Biol. 17, 4355-4362.
Kao, H. Y., Ordentlich, P., Koyano-Nakagawa, N., Tang, Z., Downes, M., Kintner, C. R., Evans, R. M. and Kadesch, T.(1998). A histone deacetylase corepressor complex regulates the Notch signal transduction pathway. Genes Dev.12, 2269-2277.
Kopan, R.(2002). Notch: a membrane-bound transcription factor. J. Cell Sci.115, 1095-1097.
Krejci, A. and Bray, S.(2007). Notch activation stimulates transient and selective binding of Su(H)/CSL to target enhancers. Genes Dev.21, 1322-1327. Kumar, J. P. and Moses, K.(2001). The EGF receptor and notch signaling
pathways control the initiation of the morphogenetic furrow during Drosophila eye development. Development128, 2689-2697.
Lai, E. C.(2004). Notch signaling: control of cell communication and cell fate.
Development131, 965-973.
Lai, E. C. and Rubin, G. M.(2001). Neuralized is essential for a subset of Notch pathway-dependent cell fate decisions during Drosophila eye development.
Proc. Natl. Acad. Sci. USA98, 5637-5642.
Lecourtois, M. and Schweisguth, F.(1995). The neurogenic suppressor of hairless DNA-binding protein mediates the transcriptional activation of the enhancer of split complex genes triggered by Notch signaling. Genes Dev.9, 2598-2608.
Li, Y. and Baker, N. E.(2001). Proneural enhancement by Notch overcomes Suppressor-of-Hairless repressor function in the developing Drosophila eye.Curr. Biol. 11, 330-338.
Logeat, F., Bessia, C., Brou, C., LeBail, O., Jarriault, S., Seidah, N. G. and Israel, A.(1998). The Notch1 receptor is cleaved constitutively by a furin-like convertase. Proc. Natl. Acad. Sci. USA95, 8108-8112.
Miller, D. T. and Cagan, R. L.(1998). Local induction of patterning and programmed cell death in the developing Drosophila retina. Development125, 2327-2335.
Moloney, D. J., Panin, V. M., Johnston, S. H., Chen, J., Shao, L., Wilson, R., Wang, Y., Stanley, P., Irvine, K. D., Haltiwanger, R. S. et al.(2000). Fringe is a glycosyltransferase that modifies Notch. Nature406, 369-375.
Nagel, A. C., Krejci, A., Tenin, G., Bravo-Patino, A., Bray, S., Maier, D. and Preiss, A.(2005). Hairless-mediated repression of notch target genes requires the combined activity of Groucho and CtBP corepressors. Mol. Cell. Biol. 25, 10433-10441.
Nellesen, D. T., Lai, E. C. and Posakony, J. W.(1999). Discrete enhancer elements mediate selective responsiveness of Enhancer of split Complex genes to common transcriptional activators. Dev. Biol. 213, 33-53.
Okajima, T. and Irvine, K. D.(2002). Regulation of notch signaling by o-linked fucose. Cell111, 893-904.
Pan, D. and Rubin, G. M.(1997). Kuzbanian controls proteolytic processing of Notch and mediates lateral inhibition during Drosophila and vertebrate
neurogenesis. Cell90, 271-280.
D
E
Rusconi, J. C., Hays, R. and Cagan, R. L.(2000). Programmed cell death and patterning in Drosophila. Cell Death Differ.7, 1063-1070.
Selkoe, D. and Kopan, R.(2003). Notch and Presenilin: regulated intramembrane proteolysis links development and degeneration. Annu. Rev. Neurosci.26, 565-597.
Simpson, P.(1997). Notch signaling in development: on equivalence groups and asymmetric developmental potential. Curr. Opin. Genet. Dev. 7, 537-542. St Pierre, S. E., Galindo, M. I., Couso, J. P. and Thor, S.(2002). Control of
Drosophila imaginal disc development by rotund and roughened eye: differentially expressed transcripts of the same gene encoding functionally distinct zinc finger proteins. Development129, 1273-1281.
Sun, Y., Jan, L. Y. and Jan, Y. N.(1998). Transcriptional regulation of atonal during development of the Drosophila peripheral nervous system. Development
125, 3731-3740.
Terriente Felix, J., Magarinos, M. and Diaz-Benjumea, F. J.(2007). Nab controls the activity of the zinc-finger transcription factors Squeeze and Rotund in Drosophila development. Development134, 1845-1852.
Van Vactor, D. L., Jr, Cagan, R. L., Kramer, H. and Zipursky, S. L.(1991). Induction in the developing compound eye of Drosophila: multiple mechanisms restrict R7 induction to a single retinal precursor cell. Cell67, 1145-1155. Wallberg, A. E., Pedersen, K., Lendahl, U. and Roeder, R. G.(2002). p300 and
PCAF act cooperatively to mediate transcriptional activation from chromatin templates by notch intracellular domains in vitro. Mol. Cell. Biol. 22, 7812-7819. Wilson, J. J. and Kovall, R. A.(2006). Crystal structure of the
CSL-Notch-Mastermind ternary complex bound to DNA. Cell124, 985-996.
Wolff, T. and Ready, D. F.(1993). Pattern formation in the Drosophilaretina. In
The Development of Drosophila melanogaster (ed. M. Bate and A. Martinez-Arias), pp. 1277-1326. Cold Spring Harbor, NY: Cold Spring Harbor Press. Xu, T. and Rubin, G. M.(1993). Analysis of genetic mosaics in developing and
adult Drosophila tissues. Development117, 1223-1237.
Zhou, S., Fujimuro, M., Hsieh, J. J., Chen, L., Miyamoto, A., Weinmaster, G. and Hayward, S. D.(2000). SKIP, a CBF1-associated protein, interacts with the ankyrin repeat domain of NotchIC To facilitate NotchIC function. Mol. Cell. Biol.
20, 2400-2410.