ith 6 figures in Great Britain
THE POSSIBLE INVOLVEMENT OF BRAIN MICROTUBULES
IN MEMORY FIXATION
BY JOHN CRONLY-DILLON, DAVID CARDEN AND CAROLE BIRKS
Visual Sciences Laboratories, Department of Ophthalmic Optics, University of Manchester Institute of Science and Technology,
Manchester, England
{Received 4 April 1974)
SUMMARY
Goldfish were trained to discriminate between two coloured lights. When colchicine was administered intracranially immediately after training it produced complete retrograde amnesia 2 days later, but the fish dis-played considerable retention if instead the injection was given 1-25 h after training. If colchicine was administered 1-25 h before training the fish were able to learn as readily as control fish but then showed retrograde amnesia when tested 2 days later. It is suggested that colchicine may affect memory fixation by interfering with the fast transport of material needed to bring about a structural modification at certain synapses.
INTRODUCTION
Restriction of sensory experience during early postnatal life has been found to change cell morphology (Cragg 1969; Globus & Schiebel, 1967; Mollgard et al. 1971; Rozenzweig 1971) and alter the pattern of functional connectivity within the brain (Blakemore & Cooper, 1970; Gaze et al. 1970, Hubel & Wiesel 1970).
According to Valverde (1967) rearing mice in darkness produces a loss of spines on pyramidal cells of the visual cortex, and Volkmar & Greenough (1972) report that rearing complexity affects branching of cortical dendrites.
Despite the many interesting reports of environmentally induced changes in brain function and morphology, the mechanism whereby either early experience or learning induces such enduring effects upon the functional organization of neuronal networks is still unclear. Yet such factors within the cell, that regulate the growth and trophic maintenance of cytoplasmic processes and synaptic connexions must clearly play a central role in bringing about some <<f the long-term changes that arise from learning or early experience, but until recently (Rose, 1970) these have attracted com-paratively little attention.
F N D OG B OG ND F \
Projector
Projector
Train Rest Train
L_
Shock
80 120
Time sec.
Fig. i. Apparatus used for training the fish. F, gelatin colour filter, ND, neutral density filter, OG, opal glass plate, B, barrier. Electric shocks were delivered through grid electrodes (not shown) situated across the width of the tank on each side of the barrier.
and perhaps also, for the formation or maintenance of its synapses. Roisen et al. (1972) have shown that axonal elongation depends on microtubule formation where the latter was stimulated by cyclic AMP which initiates tubule assembly from a pre-existing pool. Also, drug-inhibition of fast axonal transport mimics the effect of axotomy (Pilar & Landmesser, 1972) and leads to functional depression of synapses (Perisic & Cuenod, 1972) and morphological signs of synaptic degeneration after a few days (Cuenod, Sandri & Akert, 1972).
Furthermore, experiments using radioactively labelled precursors have shown that the molecular constituents of cell membranes are continuously turning over (Smith, 1968; Warren & Glick 1969; Schmitt 1973). Since these constituents are responsible for imparting to the cell and its membrane many of their specialized functions, any alteration in supply of materials to the membrane could affect the number or nature of molecules available for incorporation; perhaps modifying the membrane's proper-ties, and thereby possibly the cell's functional relationship with other cells. If stable memory is recorded by the brain in the form of some altered growth pattern, then memory storage must seemingly involve some alteration of membrane structure where molecular constituents are either resorbed or incorporated locally into the membrane to produce a functionally altered pattern of synaptic connectivity (Deutsch,
I972)-As a working hypothesis, we therefore considered the possibility that microtubules may be involved in conveying the necessary materials required to bring about the local changes in membrane structure or cell morphology that result from learning or early experience.
MATERIALS AND METHODS
Goldfish were obtained from a local pet dealer. Before training they were kept with other fish in communal stock tanks where they were fed on Tetrafin Goldfish Flakes. After training, each was transferred to an individual tank. The water temperature in all the experiments was approximately 20 °C and before training or testing, individual fish were always left for at least 20 min to familiarize themselves with the training tank.
Initial training
The fish were trained in a shock avoidance situation, to discriminate between two illuminated panels of different colour, situated at opposite ends of the training tank. Yellow (Amax = 575 nm) was chosen as the positive, and blue ( A ^ j = 463 nm),
the negative light stimulus. The choice of yellow as the positive stimulus was in part dictated by the fact that most fish appear to have an innate preference to swim to-wards blue (Bauer, 1910; Muntz & Cronly-Dillon, 1966; Muntz, 1962). Relative brightness was not controlled. Nevertheless, while it is conceivable that some fish may have learned to distinguish the stimuli on the basis of brightness, subsequent transfer tests on four control fish, where relative brightness between the two stimulus lights was randomized, suggests that the fish were actually discriminating on the basis of the difference in colour. In the early stages of this study we did carry out a few experiments with blue positive and these produced results in agreement with those obtained with yellow as the positive stimulus. However, because of the blue preference which goldfish display these were discontinued since we felt that a much more con-vincing case could be presented by training the fish to respond against an innate preference.
Each trial which lasted 40 sec allowed the fish to swim through a gap in the barrier dividing the tank, to whichever side was illuminated with the positive stimulus (Fig. 1). If a fish found itself on the wrong side of the tank, or failed to swim to the correct side within the first 24 sec, it received an electric shock (approximately 2 mA 59 c/s a.c.) from which it could escape by swimming to the correct end of the tank. The position of the positive stimulus was of course randomized from trial to trial. Interposed between successive trials were rest periods of 40 sec duration, where each end panel of the tank was illuminated uniformly with the same neutral grey light (10 ft Lamberts).
10
-9
8
S 7
Saline
10
-12-5 (ig colchicine
10 20 30 40 50 60
Trials (i)
10 20 30 40 50 60 Trials
[image:4.451.92.346.48.216.2](ii)
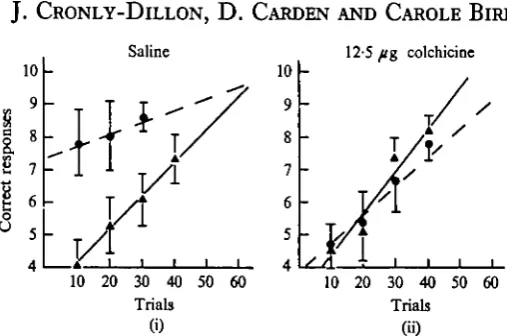
Fig. 2. Regression lines showing performance during initial training ( A — A ) and Transfer retraining ( • • ) for (i) saline control group (15 fish) and (ii) experimental Group 1 (15 fish) which received 12-5 fig colchicine intracranially immediately after initial training. Solid circles and triangles represent mean score for each of the groups, and standard deviation is represented by the vertical lines.
Transfer retraining
Each fish was retrained 36-48 h after initial training to determine if it displayed any significant improvement in performance to reach criterion as compared to initial training.
Statistical analysis
Regression lines were computed for the initial learning and transfer of each of the control and experimental groups using the data from the first 3 blocks of 10 trials. Likewise the analysis of variance on the data was always computed using the scores in the first two blocks of 10 trials achieved in the training or transfer sessions.
Intracramal injections
All injections were given intracranially into the subdural space overlying the optic tectum. The technique for administering the injections was the same as that described by Agranoff, Davis & Birch (1965).
Control fish received 50 (A saline intracranially. Our experimental groups received
I2'5/"g colchicine (Hopkin & Williams Ltd) in 50 fi\ saline. Two groups received
25 fig colchicine in 50 /A saline.
Experimental and control groups generally consisted of 15 fish, where each fish weighed approximately 13-14 g. One experimental group consisted of 7 fish (Fig. 4).
RESULTS
Saline controls
The purpose of this control group was to determine if the trauma of an intracranial
injection given immediately after training affected memory fixation. Fifteen fish were
447
10 r 10 r
9 8 7 6 5
10 20 30 40 50 60 Trials
(i)
10 20 30 40 50 60 Trials
[image:5.451.86.347.42.225.2]GO
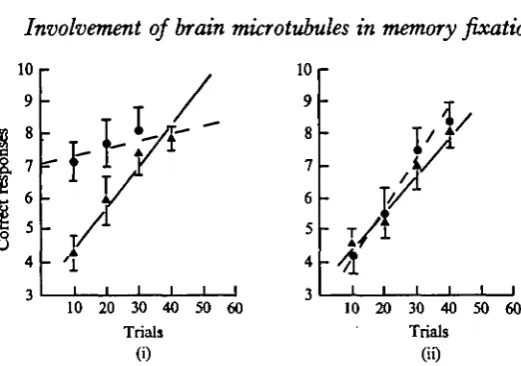
Fig. 3. Regression lines showing performance during initial training ( A — • ) and transfer retraining ( • • ) for (i) experimental Group a (15 fish) which received 12-5 fig colchi-cine 1 h and 15 min after initial training and (ii) experimental Group 3 (15 fish) which received
2
5 Ag colchicine 1 h and 15 min before initial training.
Regression lines have been plotted for learning (solid line) and transfer retraining (dashed line). It is dear from the figure that performance during the first 20 trials of transfer retraining was considerably better than during initial training. This was confirmed statistically by analysis of variance on the scores in the first 20 trials for initial training and transfer (F = 3s, d.f. = 28, P 0-005).
The group as a whole therefore displayed considerable retention of the originally learned task during transfer retraining.
Experimental Group 1
Does colchicine injected immediately after training affect memory fixation?
The same procedure was followed here as for the saline control group, except that the intracranial injection now contained 12-5 fig Colchicine. The results for this group are shown graphically in Fig. 2(ii). This time there was no statistically significant difference (F = 0-17, d.f. = 28, Pc-25) between scores during initial training and transfer retraining. This group therefore displayed complete retrograde amnesia for the originally learned task during the retraining session.
Experimental Group 2 Does colchicine affect established long-term memories?
10
-I/--10 20 30
Trials
40 50 60
Fig. 4. Regression lines showing performance during initial training (solid line) and transfer retraining (broken line) for a group (7 fish) that received 25 fig colchicine 1 h and 4s min after initial training.
Experimental Group 3
Does colchicine administered before training interfere with initial learning and short-term memory?
This group were given an intracranial injection of colchicine, in this case 25 /Jg in 50 /A saline, 1J h before initial training. After training they were returned to their individual tanks where they remained for 36-48 h before being subjected to the transfer retraining tests.
Fig. 3 (ii) shows the results for this group. The performance during initial learning was not significantly different from that of the saline control group (F = 0-638, d.f. = 28, P 0-25). Hence it would appear that colchicine administered through the intracranial route does not interfere with initial learning and short term memory, at least not for the first hour or two following the colchicine injection.
On the other hand, when these same fish were tested 2 days later the group dis-played considerable loss of memory for the task they had previously learned, and their performance was not significantly different from that they achieved during initial learning (F = 0-036, d.f. = 28, P 0-25).
ELECTROPHYSIOLOGICAL EXPERIMENTS
Colchicine blockade of axoplasmic flow is known to depress synaptic transmission (Perisic & Cuenod 1972). In doves it was found that the effect of a large (100 fig) intraocular injection of colchicine on synaptic transmission at the tectum was pro-gressive and developed slowly to a maximum approximately 7 days after the injection.
Amplifier
449
Micro-electrode
Pre-amplifier
Indifferent / ^ V electrode <? /Display
Audio amplifier
Noise blanker
Speaker
Pulse rate intergrater
Monitor Joscilloscope
PDP-12 computer
ADC input Trigger output
XY
[image:7.451.63.386.48.179.2]recorder
Fig. 5. Arrangement of apparatus used for recording from single neurones in the optic tectum of the goldfish.
discharges or a reduction in neuronal activity, are known to produce amnesia, it was important, in the present series of experiments to determine if the doses of colchicine we employed had any effect on the brain's electrical activity during the first hour following the colchicine injection, when recently acquired memory is unstable.
Methods and results
Goldfish were anaesthetized by immersing them in a 0-4% solution of MS222 (Sandoz). The optic tectum was then exposed by making a window in the skull. The fish was then mounted in a special holder that allowed ordinary tap water to flow into the mouth and past the gills. Woods metal microelectrodes (Gesteland et al. 1959) were used to record responses from single units in the optic tectum. The receptive field characteristics of goldfish optic nerve fibres have been extensively studied by several investigators (Cronly-Dillon, 1964; Jacobson & Gaze, 1964). In the present study we selected units whose receptive field characteristics and spontaneous dis-charge patterns were different from those observed with optic nerve fibres. We are therefore confident that the majority of responses that we examined probably origi-nated from tectal neurons.
Fig. 5 shows the arrangement of apparatus for recording. Once the receptive field of a suitable unit was located, the screen of a CRO carrying the stimulus display, was brought into coincidence with it. The stimulus used to drive the unit was a vertical stripe (4-5° x 0-4°) with a brightness of 10 ft Lamberts, that was moved across the oscilloscope screen at a velocity of 2-60 deg/sec. The unblanked sweep returned at 4-5° in 0-05 sec. From the fish's viewpoint, the movement of the stimulus was in a temporo-nasal direction.
In the majority of cases this was not the most effective stimulus, however it was sufficient for the purpose of this experiment. (The background illumination in these experiments was o-i ft Lamberts.)
Movement of the bar across the screen was triggered by a pulse delivered by a PDP 12A computer. The response recorded with the microelectrode was also relayed to the computer which then displayed the time average integrated response to 8 con-secutive sweeps of the moving stimulus, where sweep time was 2 sec and the com-puter stored one thousand points per sweep.
Before injection Colcbicine
3
15 min
30min
45 min
60 min
Sweep time Isec
Fig. 6. Average responses recorded from a single neurone in the optic tectum to repetitive visual stimulus before and after topical application of a colchicine solution to the tectum. Each trace represents the average integrated response to 8 consecutive sweeps of a moving visual stimulus which in this instance was a vertical bar of light that was moved repetitively across an oscilloscope screen.
Although there were sometimes small changes in amplitude of recorded action potentials during recording sessions, due to slight electrode movement, the colchicine appeared to have little effect on the gross electrical activity of visually driven cells in the optic tectum, at least over the one hour period following colchicine administration, when memory storage for a newly learned response is unstable (Fig. 6).
PROTEIN SYNTHESIS
Is the effect on memory storage due to a colchicine induced inhibition of protein synthesis?
Our previous experiments in this series have shown that memory is stable i-J h after training when it is no longer susceptible to interference by colchicine. However, as other experiments have shown (Agranoff et al. 1965, 1973; Mark & Watts, 1970; Watts & Mark, 1970; Neale, 1970) inhibitors of protein synthesis can affect the transfer from short to long term memory if they are injected into the animal immediately after training It was therefore important to determine, in our present series of experiments, if gross protein synthesis in the brain of the fish was affected after 1 h exposure to colchicine (i2'5/ig colchicine in 50 /A saline).
Methods and results
Twenty-four fish were used for these experiments (weighing approximately 13-14 g) and they were separated into three groups of 8 fish.
Table 1. Incorporation of \^lC]L-leucine in brain protein 1 h 15 min after injecting the labelled amino-acid intraperitoneally
1 (dia/min brain protein)
n (dis/min supernatant)
3.D.
Puromycin group (n = 8) 0-3661 0-1694 Colchicine group (n = 8) 0-9019 0-295 Control group (n = 8) 0-8125 0-2236
in saline. Two of these groups were then immediately given intracranial injections. One group received 12*5 /ig Colchicine in 50 /A saline, the other group received 170/ig Puromycin (Serva, Heidelberg) also in saline, the latter being the dose reported to produce complete retrograde amnesia in goldfish (Agranoff et al. 1965). The third group acted as controls and received no intracranial injections.
After i\ h the optic tectum of each fish was removed and weighed (after being first anaesthetized with 0-4% solution of MS 222). The excised tectum was then dropped into 1 ml of cold (5 °C) TCA (10%) containing 10 HIM unlabelled L-leucine, the tissue was immediately homogenized with a glass rod and left overnight in a cold room (5 °C). After centrifugation 0-5 ml supernatant was removed, and transferred to a counting vial with 5 ml scintillation fluid, 0-5 ml (5%) TCA+ 10 HIM L-leucine were added to the pellet which was heated (80 °C) for 20 min then washed twice in 5% TCA. Ethanol ether (4:1) was then added to the pellet and left at room tempera-ture for 20 min followed by two washes in ethanol ether. Finally after adding 0-5 ml Soluene (Packard) to the pellet and leaving for 15 min at 60 °C, the protein solution was transferred to a counting vial with 5 ml scintillation fluid (consisting of P.P.O., P.O.P.O.P., Toluene and Triton-X-100). Sample quenching was assessed by the channels ratio method.
Results are shown in Table 1. Also comparisons between control and puromycin groups, and control and colchicine groups, were carried out using the f-test. Protein synthesis in the puromycin group was clearly depressed as compared to controls
(t = 4-5007, d.f. = 14, P < o-oi). On the other hand the difference between the
colchicine and control groups was not statistically significant (t = o-68oi, d.f. = 14;
P > 0-25). Hence it was concluded that 1 \ h following the injection of 12-5 fig
colchi-cine there was no detectable interference with gross protein synthesis in the brain.
DISCUSSION
injection prior to training. Furthermore the fact that the same fish subsequentl^P displayed retrograde amnesia for the task suggests it is unlikely that the effect of colchicine on memory can be attributed to any discomfort induced by the drug. Also it will be recalled that when the colchicine injection was administered 1-25 h after training the memory was found to be no longer susceptible to interference by the drug. We interpret these findings as meaning that the cellular mechanism responsible for transferring learned information from the short to the long-term store, may, as others have suggested (Mark & Watts, 1970; Watts & Mark, 1970), comprise several components, and that while one of these is sensitive to inhibitors of protein synthesis, another appears to be susceptible to interference by the same agents which interfere with the fast transport of materials within cells.
A converse effect on growth of cellular process and on memory fixation has also been reported for agents which promote the formation or stabilization of microtubules. Heavy water, deuterium oxide, which stabilizes microtubules by substituting deu-terium for hydrogen, has been found to accelerate the growth of cell processes in isolated nerve tissues (Murray & Benitez, 1968), and Lehr, Wenzel & Werner (1970) also report that heavy water facilitates learning in goldfish.
Furthermore, a possible relationship between bioelectric activity and microtubule activation is suggested by the results of Kerkut, Shapira & Walker (1967) who showed that axonal transport in a motor nerve of the snail was enhanced in proportion to the increase in discharge rate of the nerve. In addition, other experiments point to cyclic AMP in some cells, as perhaps providing a possible link between synaptic activity and the assembly of microtubules (see also Rasmussen 1970). Indeed, Goodman et al. (1972) found that cyclic AMP will initiate the in vitro polymerization of microtubule subunits; and Kebabian & Greengard (1971) and Kebabian, Petzold & Greengard (1972) have described a dopamine sensitive adenyl cyclase that was activated by synaptic activity.
We therefore suggest, as a possible component of the process of memory fixation, that a local increase in intracellular cyclic AMP, produced by synaptic activation of adenyl cyclase, may in some synapses, initiate tubule assembly, and hence increase the number of ' supply lines' from the cell soma. In modifiable neurones, we further suggest that these could then serve to direct the movements of various materials to the vicinity of synapses that have recently been activated.
In our present study, the effect of the drug colchicine on memory is attributed to its possible interference with an intermediate link between the short-term memory process and the stage requiring protein synthesis; and the role of microtubules in memory fixation is also viewed as perhaps mediating the fast transport of some intra-cellular material needed to bring about a structural change at certain synapses in the brain that are involved in the original learning of the task.
REFERENCES
AGRANOFF, B. W., DAVIS, R. E. & BRINK, J. J. (1965). Memory fixation in goldfish. Proc. natn. Acad.
Set. U.S.A. 54, 788-93.
AGRANOFF, B. W., DAVIS, R. E. CASOLA, L. & LIN, R. (1973). Actinomycin D blocks formation of
memory of shock avoidance in goldfish. Science, N. Y. 179, 1243.
BAUER, V. (1910). Uber das Farbenunterscheidungsvermogen der Fische. PflUgers Arch. ges. Pkytiol. 133.
7-*G-BLAKBMORE, C. & COOPER, G. F. (1970). Development of the brain depends on the visual environment.
Nature, Land. 338, 447-8.
CRAGG, B. G. (1969). The effects of vision and dark rearing on the size and density of synapses in the lateral geniculate nucleus measured by electron microscopy. Brain Res. 13, 53-67.
CRONLY-DILLON, J. (1964). Units sensitive to direction of movement in goldfish optic tectum. Nature,
Lond. 303, 214-15.
CUBNOD, M., SANDRI, C. & AKERT, K. (197a). Enlarged synaptic vesicles in optic nerve terminals induced by intraocular injection of colchicine. Brain Ret. 39, 285—96.
DEUTSCH, J. A. (1971). The cholinergic synapse and the site of memory. Science, N.Y. 174, 788-94.
GAZE, R. M., KEATING, M. J., SZEKBLY, G. & BEAZLEY, L. (1970). Binocular interaction in the formation
of specific intertectal neuronal connections. Proc. R. Soc. B 175, 107-47.
GESTELAND, R., HOWLAND, B., LETTVIN, Y. J., MCCULLOCH, W. & PITTS, W. (1959). Comments on
microelectrodes. Proc. Jnst. Radio Engrs 47, 1856-62.
GLOBUS, A. & SCHIEBEL, A. B. (1967). The effect of visual deprivation on cortical neurons: a Golgi study. Expl Neurol. 19, 331-45.
GOODMAN, D. B. P., RASMUSSEN, H., DI-BELLA, C. F. E. & GUTHROW, C. E. (1970). Cyclic adenosine
3'5'monophosphate stimulated phosphorylation of isolated neurotubule subunits. Proc. natn. Acad.
Set. U.SJl. 67, 652-9.
HUBEL, D. H. & WiBBEL, T. N. (1970). The period of susceptibility to the physiological effects of unilateral eye closure. J. Pkytiol. 306, 419-36.
JACOBSON, M. & GAZE, R. M. (1964). Types of visual responses from single units in the optic tectum and optic nerve of the goldfish. Q. Jl Exp. Phytiol. 49, 199-209.
KBBABIAN, J. W. & GREBNGARD, P. (1971). Dopamine-sensitive adenyl cyclate: possible role in Synaptic Transmission. Science, N.Y. 174, 1346-9.
KEBABIAN, J. W., PETZOLD, G. C. & GREBNGARD, P. (1972). Dopamine-sensitive Adenylate cyclase in
Caudate nucleus of rat brain and its similarity to the 'dopamine receptor'. Proc. Nat. Acad. Set.
U.S.A. 69, 2145-9.
KERKUT, J., SHAPIRA, A. & WALKER, R. J. (1967). The transport of labelled material from CNS muscle along a nerve trunk. Camp. Biochem. Phytiol. 33, 729-48.
LEHR, E., WERNER, M. & WENZEL, G. (1970). Zur biochemie des gedSchtnisses. I. Einfluss von schwerem wasser auf das ged§chtnis von Fischen. Natunoissenschaften, 57, 521—4.
MURRAY, M. R. & BENITEZ, H. (1968). Action of heavy water (DtO) on growth and development of isolated nervous tissue. In Growth of the Nervous Syttem (ed. G. E. W. Wolstenholme and M. O'Connor). London: J. and A. Churchill Ltd.
MARK, R. F. & WATTS, M. E. (1970). Drug inhibition of memory formation in chickens. 1. Long-term memory. Proc. R. Soc. B 178, 439-54.
MOLLOAARD, K., DIAMOND, M., BENNET, E. L., ROSENZWEIG, M. & LINDWTN, B. (1971). Quantitative
synaptic changes with differential experience in rat brain. Int. J. Neurosd. 3, 113-28.
MUNTZ, W. R. A. (1962). Effectiveness of different colours of light in releasing the positive phototactic behaviour of frogs and a possible function of the retinal projection to the diencephalon. J.
Neuro-physiol. 3S, 712-20.
MUNTZ, W. R. A. & CRONLY-DILLON, J. R. (1966). Colour discrimination in goldfish. Anim. Behav. M.
351-55-NEALE, J. H., KLINGER, P. D., AGRANOFF, B. W. (1973). Campothecin blocks memory of conditioned avoidance in goldfish. Science, N.Y. 179, 1243.
OCHS, S. (1972). Fast transport of materials in mammalian nerve fibres. Science, N. Y. 176, 253—60. PERISIC, M., & CUENOD, M. (1972). Synaptic transmission depressed by colchicine blockade of
axoplasmic flow. Science, N.Y. 175, 1140-2.
PILLAR, G. & LANDMESSER, L. (1972). Axotomy mimicked by localized colchicine application. Science,
N.Y. IT7, 1116.
RASMUSSEN, J. (1970). Cell communication, calcium ion, and cyclic adenosine monophosphate. Science, N.Y. 170, 404-12.
ROISEN, F . J., MURPHY, R. A. PICHICHERA, M. E. & BARDEN, W. G. (1972). Cyclic adenosine
mono-phosphate stimulation of axonal elongation. Science, N. Y. 175, 73-4.
ROSE, S. P. R. (1970). Neurochemica lcorrelates of learning and environmental change. In Short
Memory Changes in Neural Activity and Behaviour (ed. G. Horn and R. A. Hinde). Cambridge
versity Press.
ROSENZWEIO, M. (1971). Effect of environment on development of brain and behaviour. In
Biopsy-chology of Development (ed. E. Toback). Academic Press.
SCHMITT, F. O. (1973). Molecular membranology. In The Dynamic Structure of Cell Membranes (ed. by D. F. Hotel). Wallack.
SJOSTRAND, J. & HANSON, H. (1971). Effect of colchicine on the transport of axonal protein in the retinal ganglion cells of the Rat. Expl Eye Res. 12, 261-9.
SMITH, M. E. (1968). The turnover of myelin in the adult rat. Biochim. Biophys. Acta, 164, 285-93. VALVERDE, F. (1967). Apical dendritic spines of the visual cortex and light deprivation in the mouse.
Expl Brain Res. 3,
337-52-VOLKMAR, F. R. & GREENOUOH, W. T. (1972). Rearing complexity affects branching of dendrites in the visual cortex of the rat. Science, N.Y. 176, 1445-7.
WARREN, L. & GLICK, M. C. (1969). Membranes of animal cells. 11. The metabolism and turnover of the surface membrane. J. Cell Biol. 37, 729-46.