With 2 figures Jmted in Great Britain
STEADY STATE POTENTIAL IN THE DEVELOPING
OOCYTES OF LOCUSTA MIGRATORIA:
PASSIVE AND ACTIVE COMPONENTS
BY Z. WOLLBERG AND R. COCOS
Department of Zoology, The George S. Wise Faculty of Life Sciences, Tel Aviv University, Tel Aviv, Israel
{Received 19 January 1981)
Ovarioles of the African migratory locust (Locusta migratoria) are of the panoistic type, where each oocyte is surrounded by a layer of follicle cells, and a single ovariole forms a longitudinal chain of 'follicles' held together by an ovariole sheath. In an earlier publication (Wollberg, Cohen & Kalina, 1976) we demonstrated that in this species, a potential difference exists across the 'follicle' wall (defined hereafter as 'oocyte potential'), ranging between 30 and 55 mV (inside oocyte negative in respect to external grounded medium), with the more mature oocytes possessing larger po-tential differences than the less-mature oocytes. We have also shown that all the follicles along a single ovariole were electrotonically and histologically coupled by oocyte follicle cell gap junctions. The ionic basis of oocyte and egg potentials have been investigated in several animal species, invertebrates and vertebrates (e.g. Powers & Tupper, 1975; Miyazaki & Hagiwara, 1976; Shen & Steinhardt, 1976; Ziegler & Morrill, 1977). In most of these cases, oocyte membrane potentials were determined, primarily, by the passive distribution of potassium ions and to a lesser extent by sodium ions. For some species the involvement of an undefined metabolic electro-genie pump, in maintaining the steady-state oocyte potential, was proposed (Powers & Tupper, 1975; Miyazaki & Hagiwara, 1976; Ziegler & Morrill, 1977). In the present report we show that the steady-state oocyte potential of Locusta migratoria is mainly determined by the passive distribution of potassium ions and by an ouabain-sensitive electrogenic Na/K pump.
The insect material, ovariole preparation, general experimental conditions and electrophysiological techniques used in the present study were essentially the same as those described in a previous paper (Wollberg et al. 1976). Here we describe only some modifications. The composition of the standard physiological solution was as follows (concentrations expressed in mM): NaCl 170, KC16-4, MgClj 3-5, CaClt 2-1, NaHCOB
2-1, NaHgPOj 4-3, sucrose 10. In experiments where sodium concentrations were varied, choline ions replaced the sodium. Variations in potassium concentration were achieved by replacing potassium for or by sodium ions. Osmotic pressure, ionic strength, and pH (6-3) were kept constant in all experimental solutions.
a peristaltic pump. The routine rate of flow of perfusion solution was about i / Potential measurements were conducted while oocytes were continuously impaled? and monitored on a Tektronix dual-beam oscilloscope (Model 502 A) with a Beckman paper recorder (model RP) connected to its vertical signal output. Potential differ-ences are expressed hereafter in mean values ± 1 S.D.
The potential difference across the wall of 65 developing oocytes (45 females) ranging in size between o-6 and 2-9 mm (oocytes no. 1-3, numbered according to their location along the ovariole, starting with the terminal oocyte) was — 537 +
IO-I mV. This value of oocyte potential is in agreement with our previous findings (Wollberg et al. 1976).
A total of 25 oocytes (20 females) were tested for the effect of reduced external sodium concentration (from 176-4 to 10 mM) on the potential difference. Most oocytes (16) disclosed some depolarization (5-0 ±3-4 mV) as a result of this treatment. Three oocytes did not show any response, and three oocytes were slightly hyperpolarized ( — 2-1 ± 0-9 mV). In some of the depolarized oocytes, depolarization was preceded by a small and short transient hyperpolarization. The possibility that these three alterna-tive effects of reduced external sodium concentration, on oocyte potentials, are randomly scattered was statistically rejected (x2 test; P < o-oi, D.F. 2). Routine potential measurements across the micropipette and the agar bridge, both immersed in the experimental solutions, ruled out the possibility that we measured junction potentials.
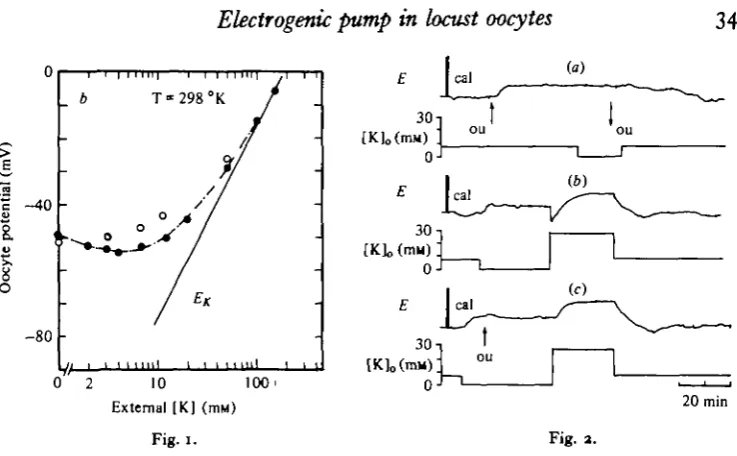
The potassium dependence of the oocyte potential was investigated on 15 oocytes (12 females). This was done by substituting K ions with or for sodium ions. The total concentration of these two ions was kept constant so that [K]0+[Na]0 = 182-8 mM. Chloride concentration was kept constant. After each test solution, and before apply-ing a new concentration, the oocyte was equilibrated in the standard physiological solution. Increasing external [K] beyond the standard concentration (6-4 mM) caused in all oocytes a depolarization the magnitude of which was related to the potassium concentration. Reducing external [K] below 6-4 mM revealed either no clear change in oocyte potential, or some depolarization. In several cases it produced a slight hyperpolarization. The depolarization which followed the increase of potassium con-centration reached a new steady state within 2-4 min. When membrane potentials were plotted against log [K]0(Fig. 1) it was found that a tenfold change of [K]o at concentrations beyond 20 mM caused a depolarization of 50-1 ± 7-4 mV. This is somewhat smaller than a slope predicted by the Nernst equation for a potassium electrode. For potassium concentrations lower than 20 mM the deviation from the predicted potassium equilibrium potential could not be explained solely by a passive contribution of other ions, less permeable than potassium, to the potential difference. This is especially true for potassium concentrations lower than 5-6*4 mM, where in most cases a depolarization was disclosed.
Electrogenic pump in locust oocytes
349
40
-a
o
E | c a l
30
(a)
[K]0(mM)
(b)
[image:3.451.37.406.52.280.2]10 100' External [K] (mM)
Fig. I.
20 min
Fig. a.
Fig. i. An illustration of the oocyte potential dependence on external potassium concentration. Re«ult8 plotted as oocyte potential v. log [KL, before (filled circles) and 15 min after exposure to ouabain (empty circles). Each change of [K]o was complete within 30 s. Standard [K]o =
6-4 mM. £*: a line predicted by the Nernst equation for a potassium electrode.
Fig. 2. Evidence for the contribution of a Na/K electrogenic pump to oocyte potential, (a) The effect of 5x10^1*1 ouabain on oocyte potential at standard potassium concentration (64 mM), and at [K]o = o. ou: application of ouabain; E, penwriter record of oocyte potential.
Upwards deflexion - depolarization, 'cal' designates 20 mV calibration pulses. [K]o, External
potassium concentration. (6) The effect of a fast increase in [K]o after its complete removal for
several min. (c) The same as (ft) except the addition of ouabain (ou, 5 x io~* min). Notice that the transient hyperpolarization was abolished.
adding the drug to: (a) a standard saline solution (8 oocytes, 8 females); (b) a free potassium solution (14 oocytes, 11 females); (c) when increasing [K]o after exposure of
the same 14 oocytes to a free K,, solution; and (d) when investigating the potassium dependence of the oocyte potential (4 oocytes, 4 females). Results were as follows: application of ouabain to the standard solution (a final concentration of 5 x io~* M) caused within about 12 min a depolarization of 4-2 ±3-5 mV (Fig. za). Complete removal of external potassium (Fig. 2b) produced, within 2-3 min, a similar depolarization (47-2-2 mV). A t test for equality of means did not show any significant difference between these two results (P > o-6, D.F. 20). In the presence of ouabain, removal of K,, does not cause any additional depolarization (Fig. 2a), and conversely, there is virtually no response to adding ouabain once external potassium is removed (Fig. 2c). This suggests that both the removal of K^and ouabain act upon the same process. Replacing the potassium-free solution by a high potassium solution caused an initial transient hyperpolarization followed by a depolarization the magni-tude of which was [K]0-dependent (Fig. 2b). This last phenomenon is generally
hyperpolarization, and the following depolarization was slightly deeper (Fig. 2 4 Investigation of the potassium dependence of oocyte potential, in the absence ariC presence of ouabain, showed that without ouabain, potentials were at all concentra-tions (except of [K]o = o) somewhat more negative (Fig. 1). These results suggest
the existence of a Na/K electrogenic pump which is abolished by both ouabain and the removal of external potassium. The validity of this pump is strongly supported by the fact that reducing external sodium concentration causes in most oocytes a slight depolarization. Considering these findings it is suggested that in Locusta migratoria, oocyte steady-state potential consists of two components: (1) the passive distribution of ions, mainly potassium, and (2) a metabolic Na/K electrotonic pump
which contributes about 8-5 % of the total negative oocyte potential.
In what follows we discuss, briefly, a possible role of a Na/K pump in the process of oocyte maturation.
Several investigators suggested that specific factors which regulate oocyte matura-tion might be transmitted from the follicle cells to the oocytes by means of g?p junctions (Anderson & Albertini, 1976; Gilula, Epstein & Beers, 1978; Browne, Wiley & Dumont, 1979). As mentioned earlier, we have shown that oocyte follicle cell gap junctions exist also in the follicles of the migratory locust. Browne et al. (1979) had demonstrated that in Xenopus laevis, uncoupling or assembly of such junctions are regulated by gonadotropic hormones.
In some tissues the maintenance of a trans-membranal sodium electrochemical gradient by an active sodium pump is essential for preservation of cell-to-cell junctions (De Mello, 1976; Cohen, 1977). In such a system the higher the Na/K coupling ratio is, the more favourable are the conditions for preserving these junctions. It is possible, thus, that the regulation of oocyte-follicle cell gap junctions by gonado-tropic hormones is mediated by an active sodium pump. That possibility is consistent with some recent findings revealing that in Rhodnius prolixus the appearance of large spaces between the follicle cells and oocyte (at the final stages of oocyte development), which is induced by the gonadotropic juvenile hormone (JH), is inhibited by ouabain, and in vitro activity of an ovarian Na/K ATPase is increased by JH (Davey & Huebner, 1974; Abu-Hakima & Davey, 1979).
REFERENCES
ABU-HAKIMA, R. & DAVEY, K. G. (1979). A possible relationship between ouabain sensitive (Na+-K+) dependent ATPase and the effect of juvenile hormone of the follicle cells of Rhodnhu prolixus.
J. Inject Biochem. 9, 195—199.
ANDERSON, E. & ALBBRTINI, D. F. (1976). Gap junctions between the oocyte and companion follicle cells in the mammalian ovary. J. Cell Biol. 71, 680-686.
BROWNE, C. L., WILEY, M. S. & DUMONT, J. N. (1979). Occyte-follicle cell gap junctions in Xenopus laevis and the effect of gonadotropin on their permeability. Science, N. Y. 303, 182-183.
COHEN, C. J. (1977). Characterization of the resting potential in Ckxronomus salivary gland cells. Expl Cell Res. 106, 15-30.
DAVEY, K. G. & HUEBNBR, E. (1974). The response of the follicle cells of Rhodnius prolixus to juvenile hormone and antigonadotropin. Can. J. Zool. 53, 14.07-14.1a.
DE MELLO, W. C. (1976). Influence of the sodium pump on intercellular communication in heart fibres: effect of intracellular injection of sodium ion on electrical coupling. J. Physiol., Lond. 263, 171—197. GILULA, N. B., EPSTEIN, M. L. & BEERS, W. H. (1978). Cell to cell communication and ovulation.
A study of the cumulus-oocyte complex. J. Cell Biol. 1, 58-75.
Electrogenic pump in locust oocytes 351
^ I , A. & HACIWARA, S. (1976). Electrical properties of the Drosophila egg membrane. Devi Biol.
S3, 91-100.
POWERS, R. D. & TUPPER, J. T. (1975). Ion transport and permeability in the mouse egg. Expl Cell Ret.
91, 412-421.
5HEN, S. & STEINHARDT, R. A. (1976). An electrophysiological study of the membrane properties of the immature and mature oocyte of the batstar, Patiria miniala. Devi Biol. 48, 148—162.