Tomasz Zaborowski
1, Kamila Wojas-Krawczyk
1, Paweł Krawczyk
1,
Olga Jankowska
1, Jan Siwiec
1, Tomasz Kucharczyk
1, 2, Maciej Grzybek
3,
Janusz Milanowski
1, 4The Effect of
CD14
and
TLR4
Gene Polymorphisms
on the Occurrence of Atopic and Non-Atopic Asthma
Wpływ polimorfizmu genu
CD14
i
TLR4
na występowanie atopowej
i nieatopowej astmy oskrzelowej
1 Department of Pneumonology, Oncology and Allergology, Medical University, Lublin, Poland 2 Postgraduate School of Molecular Medicine, Warsaw Medical University, Warsaw, Poland 3 Faculty of Biology and Animal Breeding, University of Life Science, Lublin, Poland 4 Institute of Agriculture Medicine, Lublin, Poland
Abstract
Background. A significant increase in the incidence of asthma which has been observed in recent years leads many researchers to the opinion that it is not caused only by hereditary factors but also by exposure to external factors. The fact that environmental changes cause asthma should not exclude other causes though, especially in atopic, genetically-predisposed individuals. Evidence of this phenomenon is the protective effect of lipopolysaccharide on the incidence of atopic asthma in carriers of certain polymorphic variations of LPS receptor genes: TLR4 and CD14.
Objectives. The aim of this study was to determine the genetic predisposition to the occurrence of asthma, depend-ing on the variability of the endotoxin receptor gene.
Material and Methods. The study included a group of 106 individuals diagnosed with asthma. The control group numbered 159 healthy individuals. Using the RFLP-PCR technique, polymorphism –159C < T of the CD14 gene and 1187A < G of the TLR4 gene were identified. In addition, the number of eosinophiles in peripheral blood and the total concentration of IgE in serum were determined in the group of individuals with asthma.
Results. The TT genotype in position 159 of CD14 and the AA genotype in position 1187 of TLR4 are present sig-nificantly less frequently in atopic asthma and are associated with a lower concentration of IgE in blood serum in individuals with asthma. CC homozygotes in the CD14 gene and AG heterozygotes in the TLR4 gene correspond to a higher risk of occurrence of atopic asthma, while the TT genotype of the CD14 gene and AA of the TLR4 gene are connected with a higher risk of non-atopic asthma.
Conclusion. Atopic and non-atopic asthma differ in etiology at the molecular level. Exposure to endotoxin in patients with the characteristic polymorphisms of genes for LPS receptors may have a protective effect on the development of atopic asthma (Adv Clin Exp Med 2011, 20, 4, 413–421).
Key words: asthma, polymorphism, TLR4 gene, CD14 gene, lipopolysaccharide.
Streszczenie
Wprowadzenie. Znaczące zwiększenie zachorowań na astmę oskrzelową obserwowane w ostatnich dekadach spra-wia, że większość badaczy przyczyny tego zjawiska dopatruje się bardziej po stronie zmieniającego się narażenia na czynniki zewnętrzne, niż w predyspozycjach dziedzicznych. Nie można jednak wykluczyć, że zmiany środowiska wywołują astmę oskrzelową, zwłaszcza atopową u osób predysponowanych genetycznie. Dowodem na to ma być protekcyjny wpływ lipopolisacharyduna częstość występowania astmy atopowej u nosicieli pewnych polimorficz-nych odmian genów receptorów LPS: TLR4 i CD14.
Cel pracy. Określenie genetycznej predyspozycji do wystąpienia astmy oskrzelowej w zależności od zmienności genów receptorów dla endotoksyny.
Materiał i metody. Badaniem objęto grupę 106 chorych z rozpoznaną astmą oskrzelową. Grupę kontrolną stano-wiło 159 osób zdrowych. Z użyciem metody PCR RFLP oznaczono polimorfizm –159C < T genu CD14 oraz
poli-Adv Clin Exp Med 2011, 20, 4, 413–421 ISSN 1230-025X
ORIGINAL PAPERS
The number of people with asthma in Poland and in the world is growing steadily. It is estimated that by the year 2015 the number of patients world-wide will grow to 400 million. A rapid increase in the growth rate observed in recent decades leads many researchers to the opinion that it is not only caused by hereditary factors but also by exposure to external factors [32].
One of the most widely discussed hypotheses explaining the increase of incidence of asthma and other allergic diseases is the ‘hygienic hypothesis’, by David P. Strachan, who in 1989 observed a sig-nificantly less frequent occurrence of hay fever and contact dermatitis in children from large fami-lies [31]. The author explained these findings by greater exposure to infectious agents in children from large families, which significantly influences the development of the immune system [31]. The ‘hygienic hypothesis’ was developed by further re-search which showed that increased exposure to infection causes stimulation of the immune system towards development of the Th1 lymphocyte and a lack of infectious agents in the environment re-sults in the survival of the fetal Th2 lymphocyte phenotype conducive to the development of asth-ma [20, 27].
One of the most important environmental factors affecting Th1/Th2 balance is lipopolysac-charide (LPS, endotoxin), a component of the cell wall of gram-negative bacteria. Some of the recent studies have shown a protective influence of lipopolysaccharide (LPS) on the incidence of allergic diseases in children and adults [4, 5, 9, 11, 16, 25, 26], whereas other studies have not confirmed this relationship [6, 10]. The reason for the discrepancies of the results could be the different time and duration of exposure to endo-toxin, its concentration in various environmental subjects and the existence of different polymor-phic forms of genes encoding receptors for endo-toxin in patients.
CD14 and TLR4 are the most important recep-tors responsible for transmitting signals by LPS. After entering the body, endotoxin binds to an LPB protein [34, 37]. The LBP transports
lipopoly-saccharide and forwards it to the mCD14 (mem-brane CD14), a receptor present on the surface of monocytes, the majority of macrophages, and granulocytes or sCD14 circulating in peripheral blood. sCD14 enables the binding of LPS to cells that do not possess an mCD14 receptor – endothe-lial, epithelial and dendritic cells [8, 35]. mCD14 and sCD14 in the final stage allow LPS to interact with the target receptor, which is TLR4.
Gene encoding of the CD14 receptor is located on the long arm of chromosome 5 in the close vi-cinity of the 5q31-q33 loci, in which genes are re-sponsible for the production of IgE and increased risk of asthma and overactive bronchitis [19, 22, 37, 38]. CD14 receptor activity is connected with the polymorphism of promoter region of the
CD14 gene (–159T < C). The presence of the C al-lele at position –159 of the gene correlates with increased production of IgE and the prevalence of positive skin tests. However, TT homozygotes are recorded with a higher concentration of sCD14 in serum [2].
The TLR4 gene is located on chromosome 8. The two most common polymorphic forms are 1187A < G substitution (D299G, rs4986790) and 1487C < T substitution (T399I, rs4986791). In-creased response to LPS in terms of FEV 1 decline in the provocative test of endotoxin and a higher incidence of overactive bronchitis after inhaling LPS occurs in carriers of A and C alleles in the
TLR4 gene. The occurrence of A allele in position 1187 is also associated with a higher affinity TLR4 receptor to LPS [1, 23]. On the basis of the results of these tests it can be supposed that the preva-lence of these polymorphisms may be related to the occurrence of asthma.
The aim of this study was to determine the risk of asthma development, depending on the prevalence of polymorphisms –159C < T of the
CD14 gene and 1187A < G of the TLR4 gene. It was also to demonstrate the relationship between polymorphisms of given genes and the percentage and number of peripheral blood eosinophiles and concentration of IgE in the serum of patients with bronchial asthma.
morfizm 1187A < G genu TLR4. W grupie osób chorych na astmę oskrzelową oznaczono ponadto liczbę eozynofili we krwi obwodowej oraz stężenie IgE całkowitego w surowicy krwi.
Wyniki. Genotyp TT w pozycji 159 genu CD14 oraz genotyp AA w pozycji 1187 genu TLR4 wiąże się z istotnie rzadszym występowaniem atopowej astmy oskrzelowej i mniejszym stężeniem IgE całkowitego w surowicy krwi osób chorych na astmę oskrzelową. Homozygoty CC w genie CD14 oraz heterozygoty AG w genie TLR4 charak-teryzują się większym ryzykiem wystąpienia astmy atopowej. Genotyp TT genu CD14 i AA genu TLR4 sprzyja natomiast powstawaniu astmy nieatopowej.
Wnioski. Atopowa i nieatopowa astma oskrzelowa różnią się etiologią na poziomie molekularnym. Ekspozycja na endotoksynę u chorych z odpowiednimi polimorfizmami genów receptorów dla LPS może wywierać efekt protek-cyjny na rozwój atopowej astmy oskrzelowej (Adv Clin Exp Med 2011, 20, 4, 413–421).
Material and Methods
Study Group
The study included a group of 106 patients (including 32 men and 74 women, median age = 49 years) with diagnosed bronchial asthma treated in the Department of Pneumonology, Oncology and Allergology, Medical University of Lublin in the years 2007–2008. Based on the results of skin tests and clinical symptoms, the patients were divided into groups: with atopic asthma (55 per-sons, median age = 45 years) and with non-atopic asthma (51 patients, median age = 52 years). The results of skin tests were as follows: 33 individuals were mites positive; 45 individuals were tree pol-len, grasses or weeds positive; 23 individuals were pet fur positive, and 7 individuals were mould positive. The control group included 159 healthy unrelated individuals with the excluded presence of any allergic diseases in the past (96 men and 63 women, median age = 41 years).
In all of the patients, peripheral blood samples were collected into EDTA tubes to prepare genetic tests. In addition, the number of eosinophiles in peripheral blood and the level of total IgE in se-rum was determined in the group of patients with asthma. The studies have obtained the acceptance of the local Bioethics Committee operating at the Medical University in Lublin.
DNA Isolation
DNA was isolated using the Blood Mini Kit (A & A Biotechnology, Poland) according to the manufacturer’s instructions. In brief, after remov-al of erythrocytes, leukocytes were lysed in a buffer and proteinase K. The buffer contained chaotropic salts and nonionic detergents (LT buffer). In addi-tion, proteinase K was also used in the lysis pro-cess. After rinsing the column with proteins, puri-fied DNA was eluted with a Tris buffer (10 mM TRIS.HCl pH 8.5). DNA concentration was calcu-lated using BioPhotometer Plus through the use of microliter cells Hellma TrayCell (Eppendorf AG, Germany). DNA was stored in the Tris buffer at –20°C until the examination of CD14 and TLR4
polymorphisms.
PCR-RFLP of –159C < T
Polymorphism of the
CD14
Gene
The CD14 gene fragment containing the poly-morphic site was amplified in 50 µl of a reaction mixture containing the following: 20 pmol of each of the primers: forward
5-’GTGCCAACA-GATGAGGTTCAC-3’ and reverse 5’-GCCTCT-GACAGTTTATGTAATC-3’, Taq DNA poly-merase buffer (1.5mM MgCl2), 0.25 U Taq DNA polymerase and a 0.2mM dNTP nucleotide mix-ture. The mixture was supplemented with nucle-ase-free water. The amplification was carried out in a T-Personal thermocycler (Biometer, USA) un-der the following conditions: initial denaturation at 94°C for 1 min followed by 35 cycles of 30 sec at
94°C, 30 sec at 60°C and 45 sec at 72°C with a final elongation step of 10 min at 72°C.
The PCR products were digested with AvaII Fast Digest (Fermentas, Germany) according to instruction. The restricted products were analyzed by electrophoresis in 2% agarose gel (constant am-perage 8 mA/cm2) containing ethidium bromide. The place of cutting for the restriction enzyme appeared in the amplified DNA fragment if the T allele was found in –159 position of the CD14
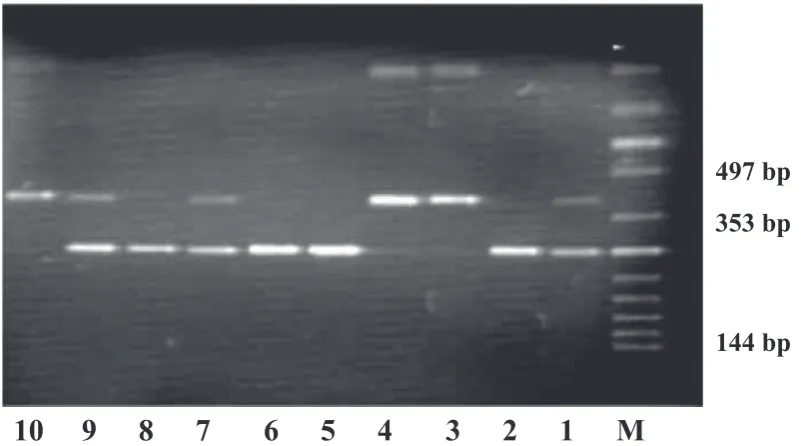
gene. In the case of CC homozygotes, an undi-gested fragment of 497bp length occurred. In the case of CT heterozygotes, fragments with a length of 497bp, 353bp and 144bp were seen. For TT ho-mozygotes, fragments with a length of 353bp and 144bp were seen (Fig. 1) [3].
PCR-RFLP of 1187A < G
Polymorphism of the
TLR4
Gene
A TLR4 gene fragment containing the poly-morphic site was amplified in 50 µl of a reaction mixture containing all the reagents described above and 20 pmol of each specified primer: for-ward 5-’GATTAGCATACTTAGACTACTAC-CTCCATG-3’ and reverse 5’-GATCAACTTCT-GAAAAAGCATTCCCAC-3’. PCR reaction was done under the following conditions: initial dena-turation (94°C for 4 min), 35 proper cycles, that included denaturation (94°C for 30 sec), hybrid-ization with primers (55°C for 10 min), elongation
(72°C for 30 sec) and finalelongation (72°C for 10 min) [18].
The PCR products were incubated with the
NcoI restriction enzyme (Fermentas, Germany) according to manufacturer’s instructions. The re-stricted products were detected by electrophore-sis on 2% agarose gel as described previously. The cutting place for the restriction enzyme appeared in amplified DNA fragments if the allele G of the
Statistical Analysis
Statistical analysis was carried using the U Mann-Whitney test to examine the differences in analyzed parameters between the two indepen-dent groups. The Kruskal-Wallis test was used to compare performance in three or more indepen-dent groups. Statistical analysis was performed us-ing Statistica v. 8.0 (StatSoft, Poland). The accura-cy Fisher test was used to establish the distribution of genotypes according to the Hardy-Weinberg equilibrium. Differences in the frequency of differ-ent alleles in patidiffer-ents with asthma and the control group were assessed by a Pearson χ2 test. Hazard ratio (HR) and 95% confidence interval calculated
for the likelihood of asthma, depending on the fre-quency of different genotypes, was calculated by using logistic regression (http://ihg.gsf.de). An as-sumption of 5% error of inference and the related level of significance p < 0.05 indicate the existence of statistically significant differences or dependen-cies.
Results
The genotype distribution of the tested genes remained in compliance with the Hardy-Weinberg equilibrium. There was no connection between the presence of polymorphic forms of the investigated
1
2
3
4
5
6
7
8
9
10
M
497 bp
353 bp
144 bp
1
2
3
4
5
6
7
8
9
10
11
12
13
14
M
223 bp
249 bp
M – DNA marker ladder; lines 2, 5, 6 and 8 – genotype CC; lines 1, 7 and 9 – genotype CT;lines 3, 4 and 10 – genotype TT.
M – drabinka DNA; linie 2, 5, 6 i 8 – genotyp CC; linie 1, 7 i 9 – genotyp CT; linie 3, 4 i 10 – genotyp TT.
Fig. 1. Representative example of polymorphism –159C < T CD14 gene
Ryc. 1. Przykładowa analiza polimorfizmu –159C < T genu CD14
M – DNA marker ladder; lines 2 and 12 – genotype AG; the other lines – genotype AA. M – drabinka DNA; linie 2 i 12 – genotyp AG; pozostałe linie – genotyp AA.
Fig. 2. Representative example of polymorphism 1187A < G TLR4 gene
genes and the risk of bronchial asthma. The inci-dence of different genotypes in a group of individ-uals with asthma and healthy subjects was similar (Table 1).
AG genotype of the TLR4 gene (p < 0.005) and CC genotype of the CD14 gene (p < 0.00001) oc-curred significantly more often in patients with atopic asthma than in subjects with non-atopic asthma, while the TT genotype of CD14 gene was observed significantly less frequently (p < 0.01). There were no statistically significant differences in the incidence of CT genotype in these groups (Table 2).
The risk of atopic asthma was significantly higher in carriers of AG genotype of the TLR4
gene than in those with AA genotype (HR = 2.33; 95% CI = 1.033–5.261; p < 0.05). By contrast, in-dividuals with AG genotype in the TLR4 gene had a significantly lower risk of non-atopic asthma
than those with AA genotype (HR = 0.081; 95% CI = 0.005–1.365; p < 0.05).
AG genotype in the TLR4 gene and CC gen-otype in the CD14 gene were observed signifi-cantly more frequently in the group of patients with atopic asthma than in healthy individuals. TT genotype of the TLR4 gene was observed with a significantly lower frequency (p < 0.05, Table 3) in atopic asthma patients. By contrast, in patients with non-atopic asthma AA genotype of the TLR4
gene was significantly more frequent than in healthy individuals (p < 0.05). AG heterozygotes were not found in the examined group (Table 4).
In patients with asthma, total IgE concentra-tion was significantly lower in individuals with TT and CT genotype of the CD14 gene than in the group with CC genotype (median levels of IgE, re-spectively: 22.95 U/ml, 65.4 U/ml, 208 U/ml; p <
Table 1. The frequency of CD14 and TLR4 gene polymorphisms in patients with asthma and in control group
Tabela 1. Częstość występowania polimorfizmów w genach CD14 i TLR4 w grupie pacjentów chorych na astmę i osób zdrowych
Gene (Gen) Genotype (Genotyp) Asthma (Astma)
n (%) Control group (Grupa kontrolna)n (%) Total (Razem)n (%)
TLR4 AA 94 (88.7) 142 (89.3) 236 (89.0) AG 12 (11.3) 17 (10.7) 29 (11.0)
CD14 CT 53 (50.0) 73 (45.9) 126 (47.5)
TT 21 (19.8) 33 (20.8) 54 (20.4) CC 32 (30.2) 53 (33.3) 85 (32.1)
Table 2. The frequency of CD14 and TLR4 gene polymorphisms in patients with atopic and non-atopic asthma
Tabela 2. Częstość występowania polimorfizmów w genach CD14 i TLR4 w grupie pacjentów chorych na astmę atopową i nieatopową
Gene (Gen) Genotype (Genotyp) Asthma (Astma) χ2 value p value
non-atopic (nieatopowe)
n (%) atopic (atopowe)n (%)
TLR4 AA 51 (100) 43 (78.2) χ2 = 10.47
p < 0.001
AG 0 (0.0) 12 (21.8)
CD14 CT 30 (58.8) 23 (41.8) χ2 = 11.08
p < 0.005
TT 16 (31.4) 5 (9.1)
CC 5 (9.8) 27 (49.1)
CT + TT 46 (90.2) 28 (50.9) χ2 = 17.56
p < 0.00005
CC 5 (9.8) 27 (49.1)
TT 16 (31.4) 5 (9.1) χ2 = 6.93
p < 0.01 CC+CT 35 (68.6) 50 (90.9)
CT 30 (58.8) 23 (41.8) χ2 = 2.42
0.01). There was no relationship between CD14
gene polymorphism and the percentage and num-ber of peripheral blood eosinophiles. In patients with AG genotype in the TLR4 gene, the level of total IgE in serum was significantly higher than among those with the AA genotype (respectively median IgE concentrations: 320 U/ml and 65.4 U/ ml, p < 0.05). There was no correlation between the presence of polymorphic forms of the TLR4
gene and the percentage and number of eosino-philes in blood.
Discussion
The results obtained of polymorphism –159C < T CD14 gene frequency are comparable to those observed in other populations. CC genotype was found in 32.1% of respondents, CT in 47.5% and TT in 20.4%. Böttcher et al. studying two groups of 115 healthy Swedish children and children with bronchial asthma found the presence of CC genotype in 37%, CT in 42% and TT in 21% of respondents [3]. In turn, Baldini et al., among 481 randomly selected children discovered the CC genotype in 29.4%, CT in 49.4% and TT in 21.3%
[2]. For comparison, the study of polymorphism –159C < T CD14 gene in the Japanese population yielded similar results: genotype CC in 32.4%, CT in 44.1% and TT in 23.5% of healthy adults [9]. In the Turkish population, similar studies have shown the presence of CC genotype in 21.1%, CT in 52.8%, and TT in 26.1% among children with bronchial asthma [28].
The incidence of polymorphism 1187A < G in the TLR4 gene did not significantly differ from the values previously reported. Genotype AA oc-curred in 89%, and the AG genotype in 11% of respondents. One of the largest studies, conduct-ed in Austria and Germany among 609 children – healthy and with bronchial asthma, showed the prevalence of genotype AA in 92.1% while the AG genotype was diagnosed in only 7.9% of subjects [7]. In the case of the Swedish population of asthmatic children, AA genotype was found in 90% and the AG genotype in 10% of patients [3]. Among 334 adults with asthma surveyed in Ger-many, the incidence of AA genotype was 91.7%, and genotype AG, 8.3% [31 = 23].
Comparison of the distribution of polymor-phisms in the group of respondents with diagnosed asthma and in healthy individuals did not show
Table 3. The frequency of different genotypes of polymorphism CD14 and TLR4 genes in patients with atopic asthma and in control group
Tabela 3. Częstość występowania różnych form polimorfizmów w genach CD14 i TLR4 w grupie pacjentów z astmą atopową i osób zdrowych
Gene (Gen) Genotype (Genotyp) Atopic astma (Astma atopowa)
n (%) Control group (Grupa kontrolna) n (%)
χ2 value
p value
TLR4 AA 43 (78.2) 142 (89.3) χ2 = 4.32
p = 0.0377
AG 12 (21.8) 17 (10.7)
CD14 CT 23 (41.8) 73 (45.9) χ2 = 5.99
p = 0.049
TT 5 (9.1) 33 (20.8)
CC 27 (49.1) 53 (33.3)
Table 4. The frequency of different genotypes of polymorphism CD14 and TLR4 genes in patients with non-atopic asthma and in control group
Tabela 4. Częstość występowania różnych form polimorfizmów w genach CD14 i TLR4 w grupie pacjentów chorych na astmę nieatopową i u osób zdrowych
Gene (Gen) Genotype (Genotyp) Non-atopic astma (Astma nieatopowa)
n (%) Control group (Grupa kontrolna) n (%)
χ2 value
p value
TLR4 AA 50 (100) 142 (89.3) χ2 = 4.48
p < 0.05
AG 0 (0) 17 (10.7)
CD14 CT 30 (58.8) 73 (45.9) χ2 = 2.42
p = 0.12
TT 16 (31.4) 33 (20.8)
the existence of statistically significant differences in the incidence of various polymorphisms of the
CD14 and TLR4 genes. However, after isolating a group of people with the atopic form of bron-chial asthma, it turned out that the CC genotype in position –159 of the CD14 gene and AG in posi-tion 1187 of the TLR4 gene occurred significantly more frequently in those patients.
The study presented is another one that shows the relationship between gene polymorphisms of receptors for endotoxin and the incidence of atopic asthma.
Koppleman et al. studied 159 patients with atopic asthma, and 158 of their healthy spouses [14]. Among patients with asthma, 32.1% were CC homozygotes in the CD14 gene, 47.8% CT heterozygotes, and 20.1% TT homozygotes. In the control group, CC genotype was found in 19.6%, CT in 53.8%, TT in 26.6% of individuals [14]. In the group of individuals with CC genotype, atopic asthma occurred significantly more often. In the study of O’Donnell et al. conducted among 718 Australian children and young adults, it was found that in patients with genotype CC in the CD14
gene, the incidence of atopy is significantly greater than in children with CT and TT genotypes. [24]. The frequency of the given genotypes in 8-year-old atopic children were: CC in 37%, CT in 45% TT in 18% of patients. In the group of children without atopy, the prevalence of genotypes was as follows: CC in 23%, CT in 50% and TT in 27% of respondents. Böttcher et al. studied 115 Swedish children and found a four times higher incidence of atopic asthma in children with genotype AG in the TLR4 gene as compared to children with geno-type AA [3].
Werner et al. studied 334 adults suffering from asthma for the presence of polymorphism of 1187A < G in the TLR4 gene and endotoxin ex-posure [32]. It turned out that among individuals with AA genotype of the TLR4 gene, higher en-dotoxin levels in house dust samples were linked with a higher incidence of bronchial asthma. By contrast, in patients with genotype AG, a reverse relationship between the level of LPS in house dust and the occurrence of overactive bronchitis was discovered.
There are also studies that have not confirmed the link between polymorphisms of the tested genes and the occurrence of allergic diseases. Kedda et al., who studied 568 patients with asthma in Aus-tralia, did not confirm the relationship between the presence of CD14 gene polymorphism and the incidence and type of asthma and the sever-ity of its course [13]. Similar results were obtained by Shibasaki et al. in the population of Japanese children with atopic asthma [30]. Two major
stud-ies conducted in Germany showed no relationship between the polymorphisms –159C < T of the
CD14 gene and IgE level and the incidence of al-lergic diseases [7, 29]. In the case of 336 patients of Caucasian origin, Yang et al. did not confirm the link between the occurrence of polymorphic forms of the TLR4 gene and the severity of the course of bronchial asthma [36]. What is more, in the popu-lation of “Huts”, T allele in position 159 was posi-tively associated with positive skin tests [23].
Only a few cited studies evaluated the con-centration of endotoxin in the environment of the respondents. The divergence of results may therefore result from different levels of endotoxin whose protective impact on the development of asthma may depend on both receptor affinity and concentration in the environment. Another rea-son for the difference in results may be the differ-ent ages of patidiffer-ents in which they were exposed to LPS. Most authors believe that the endotoxin may have a protective effect only in childhood, when by promoting the Th1 response it impedes the emer-gence of diseases associated with Th2 response.
The mechanism explaining the association of different polymorphic forms of the CD14 gene and
TLR4 gene with a frequency of asthma occurrence has not been clearly described yet. It is assumed that the rare occurrence of atopic asthma in in-dividuals with TT genotype in position –159 of the CD14 gene and in people with genotype AA in position 896 of the TLR4 gene may be associated with the increased response to endotoxin. Arbor and colleagues found a significantly smaller fall of FEV1 after inhalation of endotoxin in 83 healthy patients with genotype AA of the TLR4 gene [1]. Michel et al. also showed that individuals with AG genotype with inhaled endotoxin were character-ized by a smaller systemic response to LPS [21]. Evidence of that were the lower levels of C-reactive protein and a smaller increase of the number of leukocytes in the blood of people from this group. In turn, Levan et al., examining the relationship of polymorphism –159C < T in the CD14 gene in response to LPS [17], found, among patients with TT genotype, a significantly lower increase of sCD14 in serum. Landmann et al. have shown that the primary stimulant of the secretion of a soluble form of CD14 receptor in endotoxin assumed that TT genotype is associated with a weaker response to inhaled endotoxin [15].
signifi-cantly higher levels of IgE [14]. This relationship was also confirmed by research carried out in a group of 481 randomly selected children in Italy [2]. The relationship between IgE in the serum and the polymorphism –159C < T of the CD14 gene was also investigated by Gao et al. (a group of 300 healthy British and 200 healthy Japanese people) [9]. British individuals showed that peo-ple with TT and CT genotype had significantly lower values of total IgE compared to those with genotype CC. The authors observe reasons for the higher levels of IgE in a group of individuals with genotype CC as a weaker impact of endotoxin on these people, and thus less stimulation of cells as-sociated with Th1 response. This hypothesis is confirmed by results of a study of the relationship between the polymorphisms of 1187A < G in the
TLR4 gene and the level of total IgE [9].
The original results of this study led to the conclusion that there is a greater risk of non-atopic asthma occurrence in patients with genotypes AA in the TLR4 gene and TT in the CD14 gene.
This proves the differences in the etiology of both diseases at a molecular level. Non-atopic asthma occurs at a later age than atopic asthma, and has many features in common with chronic obstructive pulmonary disease. Th1 lymphocytes
may play a role in its formation, and endotoxin may be an additional risk factor for the occurrence of this disease. Therefore, high concentrations of LPS, as well as changes in the affinity of receptors for endotoxin, can cause non-atopic asthma.
Summarizing, the effect of the protective ac-tions of endotoxin remains debatable, and it is assumed that it can occur only if the appropriate dose of stimulation is applied and it is done within a reasonable period of time before the onset of the disease. On the basis of the results of this work it can also be assumed that this effect is strongly de-pendent on CD14 and TLR4 genes polymorphism.
The investigated population does not differ substantially from other studied Caucasian popu-lations in terms of the different polymorphisms of the CD14 and TLR4 genes. TT genotype in the po-sition 159 of CD14 and AA genotype in the posi-tion 1187 of TLR4 are present significantly less fre-quently in atopic asthma and are associated with the lower concentration of IgE in blood serum in individuals with asthma. CC homozygotes in the
CD14 gene and AG heterozygotes in the TLR4
gene have a higher risk of atopic asthma, while the TT genotype of the CD14 gene and AA genotype of the TLR4 gene could promote the formation of non-atopic asthma.
References
Woolcock AJ, Peat JK:
[1] Evidence for the increase in asthma worldwide. Ciba Found Symp 1997, 206, 122–134.
Strachan DP:
[2] Hay fever, hygiene, and household size. BR Med J 1989, 299, 1259–1260.
Martinez FD, Holt PG:
[3] Role of microbial burden in aetiology of allergy and asthma. Lancet 1999, 354, SII12–SII13.
Romagnani S:
[4] Induction of TH1 and TH2 responses: a key role for the ‘natural’ immune response? Immunol Today 1992, 13, 379–381.
Braun-Fahrländer C, Riedler J, Herz U, Eder W, Waser M, Grize L, Maisch S, Carr D, Gerlach F, Buffe A, [5]
Lauener RP, Schierl R, Renz H, Nowak D, von Mutius A: Environmental exposure to endotoxin and its relation to asthma in school-age children. N Engl J Med 2002, 347, 869–877.
Braun-Fahrländer C:
[6] Environmental exposure to endotoxin and other microbial products and the decreased risk of childhood atopy: evaluating developments since April 2002. Curr Opin Allergy Clin Immunol 2003, 3, 325–329.
Gao PS, Mao XQ, Baldini M, Roberts MH, Adra CN, Shirakawa T, Holt PG, Martinez FD, Hopkin JM:
[7] Serum
total IgE levels and CD14 on chromosome 5q31. Clin Genet 1999, 56, 164–165.
Gehring U, Strikwold M, Schram-Bijkerk D, Weinmayr G, Genuneit J, Nagel G, Wickens K, Siebers R, Crane J, [8]
Doekes G, Di Domenicantonio R, Nilsson L, Priftanji A, Sandin A, El-Sharif N, Strachan D, van Hage M, von Mutius E, Brunekreef B: Asthma and allergic symptoms in relation to house dust endotoxin: Phase Two of the International Study on Asthma and Allergies in Childhood (ISAAC II). Clin Exp Allergy 2008, 38, 1911–1920.
Leynaert B, Neukirch C, Jarvis D, Chinn S, Burney P, Neukirch:
[9] Does living on a farm during childhood protect
against asthma, allergic rhinitis, and atopy in adulthood? Am J Respir Crit Care Med 2001, 164, 1829–1834.
Portengen L, Preller L, Tielen M, Doekes G, Heederik D:
[10] Endotoxin exposure and atopic sensitization in adult pig farmers. J Allergy Clin Immunol 2005, 115, 797–802.
Portengen L, Sigsgaard T, Omland O, Hjort C, Heederik D, Doekes G:
[11] Low prevalence of atopy in young Danish farmers and farming students born and raised on a farm. Clin Exp Allergy 2002, 32, 247–253.
Celedón JC, Milton DK, Ramsey CD, Litonjua AA, Ryan L, Platts-Mills TA, Gold DR:
[12] Exposure to dust mite
aller-gen and endotoxin in early life and asthma and atopy in childhood. J Allergy Clin Immunol 2007, 120, 144–149.
Gehring U, Bischof W, Fahlbusch B, Wichmann HE, Heinrich J:
[13] House dust endotoxin and allergic sensitization in children. Am J Respir Crit Care Med 2002, 166, 939–944.
Wright SD, Ramos RA, Tobias PS, Ulevitch RJ, Mathison JC:
[14] CD14, a receptor for complexes of lipopolysac-charide (LPS) and LPS binding protein. Science 1990, 249, 1431–1433.
Frey EA, Miller DS, Jahr TG, Sundan A, Bazil V, Espevik T, Finlay BB, Wright SD:
[15] Soluble CD14 participates
in the response of cells to lipopolysaccharide. J Exp Med 1992, 176, 1665–1671.
Verhasselt V, Buelens C, Willems F, De Groote D, Haeffner-Cavaillon N, Goldman M:
lipopolysaccha-ride stimulates the production of cytokines and the expression of costimulatory molecules by human peripheral blood dendritic cells: evidence for a soluble CD14-dependent pathway. J Immunol 1997, 158, 2919–2925.
Marsh DG, Neely JD, Breazeale DR, Ghosh B, Freidhoff LR, Ehrlich-Kautzky E, Schou C, Krishnaswamy G, [17]
Beaty TH: Linkage analysis of IL4 and other chromosome 5q31.1 markers and total serum immunoglobulin E concentrations. Science 1994, 264, 1152–1156.
Noguchi E, Shibasaki M, Arinami T, Takeda K, Maki T, Mlyamoto T, Kawashima T, Kobyashi K, Hamaguchi H: [18]
Evidence for linkage between asthma/atopy in childhood and chromosome 5q31–q33 in a Japanese population. Am J Respir Crit Care Med 1997, 156, 1390–1393.
Xu J, Levitt RC, Panhuysen CI, Postma DS, Taylor EW, Amelung PJ, Holroyd KJ, Bleecker ER, Meyers DA: [19]
Evidence for two unlinked loci regulating total serum IgE levels. Am J Hum Genet 1995, 57, 425–430.
Zostma DS, Bleecker ER, Amelung PJ, Holroyd KJ, Xu J, Panhuysen CI, Meyers DA, Levitt RC:
[20] Genetic
sus-ceptibility to asthma: bronchial hyperresponsiveness coinherited with a major gene for atopy. N Engl J Med 1995, 333, 894–900.
Baldini M, Lohman IC, Halonen M, Erickson RP, Holt PG, Martinez FD:
[21] A polymorphism in the 5’ flanking region of the CD14 gene is associated with circulating soluble CD14 levels and with total serum immunoglobulin E. Am J Respir Cell Mol Biol 1999, 20, 976–983.
Arbour NC, Lorenz E, Schutte BC, Zabner J, Kline JN, Jones M, Frees K, Watt JL, Schwartz DA:
[22] TLR4
muta-tions are associated with endotoxin hyporesponsiveness in humans. Nat Genet 2000, 25, 187–191.
Werner M, Topp R, Wimmer K, Richter K, Bischof W, Wist M, Heinrich J:
[23] TLR4 gene variants modify endo-toxin effect on asthma. J Allergy Clin Immunol 2003, 112, 323–330.
Lorenz E, Frees KI, Schwartz DA:
[24] Determination of TLR4 genotype using allele-specific PCR. Biotechniques 2001, 31, 22–24.
Böttcher MF, Hmani-Aifa M, Lindström A, Jenmalm MC, Mai X, Nilsson L, Zdolsek HA, Björksten B, [25]
Söderkvist P, Vaarala O: A TLR4 polymorphism is associated with astma and reduced lipopolysaccharide-induced interleukin-12(p70) responses in Swedish children. J Allergy Clin Immunol 2004, 114, 561–567.
Sackesen C, Karaaslan C, Keskin O, Tokol N, Tahan F, Civelek E, Soyer OU, Adalioglu G, Tuncer A, Birben E, [26]
Oner C, Kalayci O: The effect of polymorphisms AT the CD14 promoter and TLR4 gene on astma phenotypes In Turkish children with astma. Allergy 2005, 60, 1485–1492.
Eder W, Klimecki W, Yu L:
[27] Toll-like receptor 2 as a major gene for asthma in children of European farmers. J Allergy Clin Immunol 2004, 113, 482–488.
Koppelman GH, Reijmerink NE, Colin SO, Howard TD, Whittaker PA, Meyers DA, Postma DS, Bleecker ER: [28]
Association of a promoter polymorphism of the CD14 gene and atopy. Am J Respir Crit Care Med 2001, 163, 965–969.
O’Donnell AR, Toelle BG, Marks GB, Hayden CM, Laing IA, Peat JK, Goldlatt J, Le Souef PN:
[29] Age-specific
relationship between CD14 and atopy in a cohort assessed from age 8 to 25 years. Am J Respir Crit Care Med 2004, 169, 615–622.
Kedda MA, Lose F, Duffy D, Bell E, Thompson PJ, Upham J:
[30] The CD14 C-159 polymorphism is not associated with asthma or asthma severity in Australian adult population. Thorax 2005, 60, 211–214.
Shibasaki M, Ichikawa K, Arinami T, Noguchi E:
[31] Failure to find an association between CD14-159C/T polymor-phism and asthma. Allergol Intern 2006, 55, 55–58.
Sengler C, Haider A, Sommerfeld C, Lau S, Baldini M, Martinez F, Wahn U, Nickel R:
[32] Evaluation of the CD14
C-159 T polymorphism in the German Multicenter Allergy Study cohort. Clin Exp Allergy 2003, 33, 166–169.
Yang IA, Barton SJ, Rorke S, Cakebread JA, Keith TP, Clough JB, Holgate ST, Holloway JW:
[33] Toll-like receptor
4 polymorphism and severity of atopy in asthmatics. Genes Immun 2004, 5, 41–45.
Ober C, Tsalenko A, Parry R, Cox NJ:
[34] A second-generation genomewide screen for asthma-susceptibility alleles in a founder population. Am J Hum Genet 2000, 67, 1154–1162.
Michel O, LeVan TD, Stern D:
[35] Systemic responsiveness to lipopolisacharide and polymorphisms in the TLR4 gene in human being. J Allergy Clin Immunol 2003, 112, 923–929.
LeVan TD, Michel O, Dentener M, Thorn J, Vertongen F, Beijer L, Martinez FD:
[36] Association between CD14
polymorphisms and serum soluble CD14 levels: effect of atopy and endotoxin inhalation. J Allergy Clin Immunol 2008, 121, 434–440.
Landmann R, Knopf HP, Link S, Sansano S, Schumann R, Zimmerli W:
[37] Human monocyte CD14 is upregulated by lipopolisaccharide. Infect Immun 1996, 64, 1761–1769.
Address for correspondence:
Paweł KrawczykDepartment of Pneumonology, Oncology and Allergology, Medical University Chodźki 4a
20-950 Lublin Poland
Tel.: +48 81 756 48 17
E-mail: [email protected], [email protected]
Conflict of interest: None declared