Indian J. Plant Physiol., Vol. XXX, No. I, pp. 44-51 (March. ]987)
EFFECT OF WATERLOGGING IN BARLEY DURING THE SEEDLING STAGE
K. WIGNARAJAH
Centre for Arid Zone Studies, University College of North Wales, Bangor, Gwynedd LL57 2UW, Wales, United Kinadom
SUMMARY
This paper discusses the regulation of ionic levels by seedlings of waterlogging sensitive and tolerant cultivars of barley during periods of waterlogging and recovery from the waterlogging stress. The tolerant cv Atlas-57 showed higher levels ofNa+ in the shoot than the sensitive cv Hispont and it appears that barley could maintain normal growth even when fairly high concentrations of sodium are found in the leaves. There was no significant differences in the uptake and transport to the shoot of K+ ions by the two cultivars. However, significantly higher rates of uptake and transport to the shoot of CI- ions were found in the sensitive cv Hispont. It is suggested that the sensitivity of barley to waterlogging may be associated with the inability to regulate the uptake and transport of CI- ions during the early stages of waterlogging stress, and that this physiological response could be used to screen barley cultivars for their ability to withsland Waterlogging stress.
INTRODUCTON
Waterlogging has sometimes been shown to result in higher levels of ADH activity in the roots of flooding sensitive species when compared with flooding tolerant species (Crawford. 1976) and this was always the case with Trifolium
subterraneum sub-species (Francis et aI., 1974). Since one of the major conse
quences of flooding was the development of low O2 tensions at the root level, attempts have been made to screen agriculturally important crops for sensitivity to waterlogging by estimating levels of ADH in the roots of plants subjected to waterlogged conditions. This approach proved to be successful with sub-species of Trifolium subterraneum (Francis et al., 1974) but not amongst the cereal crops; ADH levels in the roots were highest in rice, intermediate in the flood tolerant cv Atlas-57 and lowest in the flood sensitive cv Hispont and the flood tolerance capability of rice was significantly higher than the flood tolerant barley (Wignarajah et al., 1976). Hence, the relationship, if any, between ADH activity and tolerance to flooding was the
J":'''
WATERL()(j(JING ON BARLEY SEeDLING 45
raverse with the most Hooding tolerant plant having the highest ADH activity. Further experiments in which maize roots were subjected to varying concentra
tions of Os showed that the ADH activity could not be used to diagnose the degree of O2 deficiency to which roots had been exposed. Hence, it was conclu
ded that changes in ADH activity were neither the primary cause of reduced growth under anaerobiosis nor particularly useful in screening for tolerance to waterlogging (Smith and Ap Rees. 1979; Wingarajah and Greenway, 1976a; Wingarajah et al., I 976b).
In a previous paper it was shown that the barley cv Atlas-57 was more tolerant to waterlogging than the cv Hispont (Wignarajah et al., 1976).
Anaerobiosis increased the permeability of the roots to Na+ and
cr
ions in a waterlogging sensitive cultivar of barley (John et al.. 1977) and this increased passive transport may occur either through the cell membranes or via an extracellular pathway (Greenway, 1962; John et al., 1977). Varietal diffe rences in tolerance to salinity in soybean was associated with the ability to exclude CI-ions from the tops of the salt tolerant varieties and this exclusion mechanism was shown to be under the control of a dominant gene NCI; sensitive cultivars lacked this dominant gene resulting in the inclusion of CI- ions in their tops (Abel, 1969). Genetic control of transport of other mineral nutrients to the tops of plants has been demonstrated (Epstein, 1972) and the control mechanisms in particular for Na+ and/or Cl- ions has been suggested as being a major physiological mechanism of salt tolerance in many important agricultural crops (Wyn-]ones and Gorham, 1986). It was therefore felt that an investigation into the ionic relations of the flooding tolerant and sensitive cultivars of barley during waterlogging may give some light on the role of ionic regulation as a possible mechanism of the tolerance ability of barley to waterlogging. The results of the investigation are reported and discussed in this paper.MATERIALS AND METHODS
Barley seeds were germinated on filter papers moistened with 0.5 mM CaSO" in petri dishes. Four-day old seedlings were transferred to 15 cm diameter pots containing acid-washed sand. .On the 5th day after sowing. the plants were separated into three groups, two of which were subjected to water logging treatment, while the third was kept as a control. Each group had eight replicate plants. All plants were provided with nntrient solutions. For the waterlogging treatment, deionised water was added until the level of the water in the pot was 5.0 cm above the surface of the sand, while in the control (non waterlogged treatment) the nntrient solution was added and allowed to drain freely through the base of the sand fiU:d pot. On the 11th day after sowing,
46 K. WIGNARAJAH
one group of waterlogged plants was allowed to recover by stopping the water logging treatment and allowing the nutrient solution to drain freely as in the controls. On every third day of the experimentation all the plants were pro vided with 800 cm3 of nutrient solution containing 3mM K+, 8mM Na+, 3mM Ca2+, 2mM MgH, IImM NO,-, 5mM Cl-, 2mM SO"I-, 0.16 mM P04 2- and micronutrients according to Amon and Hoagland (1940) solution. Five plants from each treatment were harvested on the 7th. 9th. 13th, 15th and 19th day after sowing, separated into roots and stems and dried at 70°C for 3 days.
Roots and stems were digested in 10% nitric acid and the K+ and Na+ analysed by atomic absorption spectrophotometry while the Cl- ions were analys ed using an electrometric chloride analyser. Rates of ion uptake by the plant and transport of ions to the shoot were calculated using the' method of Pitman (1972).
(iP2-ipl) (1nWRI-InWR1)
(T2-T1 (WRlI-WID)
(lnWRI-lnWRl)
(WR.-WK1)
where Ui=Uptake orion, i. by the plant,
Ri=Transport of ion, i, from the root to the shoot,
is! and iSI are the ionic contents (in po moles) in the shoot at barvest times, T1 and T. respectively,
ip1 and ip:z are the ionic contents in the whole plant at Tt and TI , Wrt and Wrl are the dry weights of the roots at Tl and T2
RESULTS
WATERLOOGINO ON BARLEY SEEDLING
47
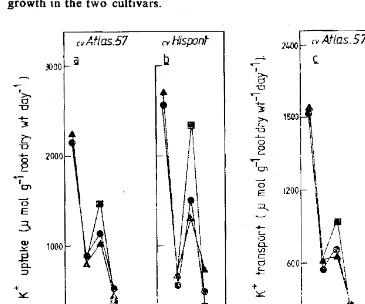
growth suggests that increased K+ translocation was the result of increased shoot growth in the two cultivars.
(
~ ':>. o. U•
...
~ ~ u...
e
~ c:n 0 E (3
Q.I ~ .g C. ::::I + !lCr
( , varscvAt/as.57 cvHis;:x;nf
g
b
..-:lOon I
Ei'
i:l...
....
I ~ :>. t._ U....
0 0 L. 2000 "Tc:n 0 E ::::1.....
L. 0 01000 III
C 0 l:: + !lC
I
L
0 I6
1410
Seedling age (days).
Fig. 1: Rate of uptake by Plant (0 & b) and shoot (c & d) of K+ ions in the two
evAtlas.57 (V Nispont
21,00
1200 .
g
c.
t
, T
, t
\1 I
~
I I
14 II 6
Seedling age (days).
rate of transport barley cultivars.
to the during waterlogging and recovery from waterlogging.
(e) Non-waterlogged (CONTROL) treatment;
<Y) Waterlogged treatment;
CD) Recovery from wateriogging treatment.
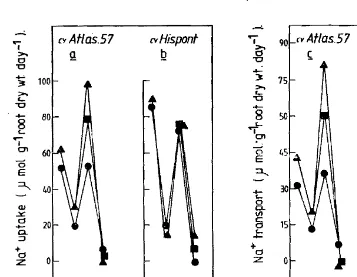
Rates of uptake and transport to the shoot of Na+ ions by the two culd are shown in Figures 3 (a-b). In cv Hispont no differences were observed in the rates of uptake or transport to the shoot of Na+ ions either due to water ,
,
r logging or the recovery treatment. However Atlas-57 showed higher levels ofuptake by the plant and transport to the shoot ofNa+ ions during the period of waterlogging. During periods of recovery these rates were lowered and values intermediate between the control and waterlogged plants were obtained •
•
•
48
"
K. WlGNARAJAH
Sodium retained by the roots was unaffected. so that any increased uptake by the plant would presumably be the result of increased transport to the shoot.
( v Atlas-57 cvHispont
0"7
06
05
+ 0 0
.c <II 04
I-~
03
02
0-1
0
I I
8 14 20
Seedling age
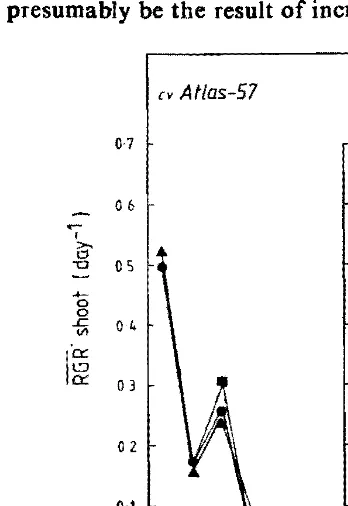
Fig. 2: Changes in relative growth rate of shoot (RGR shoot) of the two barley cultivars. duriog waterlogging and recovery from waterlogging. (key as in Fig. 1)
Figures 4 (a-b) shows the uptake and transport to the shoot of
a-
ions in the two cultivars. Atlas-57 showed low levels of CI- ion upatke and waterlogging resulted in slightly elevated levels of uptake between the 8th and 10th day of "waterlogging but returned to the levels of the control plants by the last harvestin both the waterlogged and recovering plant. On the othe hand, cv Hispont showed a high level of
a-
ion transport to the shoot in the early harvest. In fact, nearly 95% of the Cl- ions taken up by the waterlogged plants were trans ported to the shoot. With continuation of the waterlogging treatment, the rates ofcr
ion uptake and transport to the shoots declined and reached values similar to those of the non-waterlogged plants.DISCUSSION
K+ ion uptake by the plant and transport to the shoot showed no diffe rences between the sensitive and tolerant cultivars. However. when the water
8 14
pe;::
_x
I
;
W A TERLOGGlNG' ON BARLEY SEEDLING 49
-
(Y Atlcis.57....
I>. g
eI "0 +-' 100 31 >. L. "0
....
8
' . I CI -0 E :::J... <11 oX eI....
c:::L::J
+
0 z 0
-20
I
I I
8 \4 20
cvHispont
Q
8 14 2
SeedLing age (days),
...
cvAtlas.57 v Hispont
"I 90
~ ~ Q
"C
....
~ 7S
>. L. "0
....
8
' "T CI -0 E :::l.......
' 8. III C eI '....
+
0 z -15I I I
8 14 20 8 14 20
Seedling age (days)
Fig. 3: Na+ uptake by plant (0 & b) and transport to the shoot (c &. d). Key as in Fig. 1.
logging stress was removed, rates of uptake and transport to the shoot of K+ ions was substantially increased in both cultivars. Translocation of K+ ions to the sboot appears to be dependent on increases in shoot growth.
The bigher levels of Na+ ion uptake and transport to the shoot by the tolerant cv Atlas-S7 during waterlogging is significant in that there was no in crease in the accumulation and retention of these ions in the roots. The paralle lism between the transport to the shoot and the uptake by the plant suggests that the increased uptake was the result of an increased transport to the shoot. Not surprisingly, the higher rates of Na+ ions transported to the shoot did not appear to be detrimental to plant growth. Greenway (1962) has shown that high levels of sodium ions occur in salt-stressed barley without detrimental effects on growth and it is significant that the levels are higher in the water logging tolerant dutivar, Atlas-57; It is possible that in the tolerant variety, the Na+ ions could be sequestered in the vacuoles of leaf cells.
Waterlogging of cv Atlas-57 resulted in only slightly higher rates of CI ion uptake by the plants but most of the ions were retained in the rool' and
50
,~~'
\
K. WIGNARAJAH
excluded from the shoots. However, in cv Hispont large amounts of CI- ions were taken by the plants and transported to the shoot particularly during the early stage of waterlogging resulting chloride toxic effects which could make the plant sensitive to waterlogging. Abel (1969) showed similar effects of chlori de accumulation in the leaves of soybean cultivars sensitive to salinity, during growth in saline media. He showed that the ability of the tolerant cultivars to withstand and grow successfully was associated with the mechanism of exclu ding the toxic CI- ions from the shoot. He further demonstrated that the mechanism of transport of
cr
ions to the shoot was under a monogenic control, where the presence of a dominant gene NCI enabled the salt-tolerant cultivars of soybean to exc1ude CI- ions from their shoots. Gentical control of transport of other mineral nutrients to the shoots of plant has also bren reported (Epstein, 1972) and in wheat cultivars Na/K selectivity and exclusion of Na+ ions from the tops of salt-tolerent cultivars has been shown to be under genetic control (Shah et 01., 1986). Our data suggests that the sensitivity of cv Hispont to waterlogging is associated with the attainment of high levels of CI- ions in the tops of the plant during the early stages of waterlogging. The present data however does not enable us to state whether these high levels resulted from the inability of the sensitive cultivar to restrict the transport of chloride ions to the tops by retaining it in the roots and/ or due to the lack of an efficient system or retranslocation of the chloride ions from the shoot to the root. However, the present investigation have provided an important diagnostic tool for screen ing cultivars of barley for tolerance to waterlogging by monitoring chloride levels in the shoots during the early stages of waterlogging. Further studies on the genetic mechanisms involved in the exclusion of chloride ions in waterlogg ing tolerant cultivars of barley and chloride inclusion in sensitive cultivars could enable breeders to pursue an active programme of breeding for water logging tolerance in barley.REFERENCES
Abel,O.H. (1969). Inheritance of the capacity for chloride inclusion and chloride exclusion by soybeans. Crop Sci.. 9 : 697-698.
Arnon, D.1. and Hoagland, D.H. (1940). Crop production in artificial solutions and in soils with reference to factors influencing yields and absorption of inorganic ions. Soil Sci., SO : 463-484.
Crawford. R.M.M. (1967). Alcohol dehydrogenase activity in relation to flooding tolerance in roots. J. Exp. Bot., 18: 451-464.
Epstein, E. (1912). Mineral nutrition of plants: Principles and Perspectives. John Wiley &: Sons, N.Y. (412 pp).
WATERLOGGING ON BARLEY SBEDLING
51
Greenway, H. (1962) Plant response to saline substrates. I. Growth and ion uptake of several varieties of Hordeum during and after sodium chloride treatment. Aust. J. Bioi. Sci., 15: 16-38.
Pitman, M.G. (1972). Uptake and transport of ions in barley seedlings III. Correlation between transport to the shoot and relative growth rate. Aust. J. BioI. Sd, 25 : 905-919. Shah, S.H. Gorham, J., Forster, B.P., and Wyn-Jones, R.G. (1986). Salt tolerance in tho
Triticeae: The contribution of the D genome to cation selectivity in hexaploid wheats. J. Exp. Bot., 37 : (Inpress).
Smith, A.M. and Ap Rees, T. (1919). Pathways of carbobydrate fermentation in the roots of marsh plants. Planta, 14(;: 321-334.
Wignarajah, K. and Greenway, H. (1916a). Effect of anaerobiosis on activities of alcohol dehydrogenase and pyruvate decarboxylase in roots of Zea mays. New Phytol .• 77:
575-584.
Wignarajah, K., Greenway, H. and John. C.D. (1976 b). Effect of waterlogging on growth and activity of alcohol dehydrogenase in barley and rice. New Phytol.. 77:
585-592.
Wyn-Jones, R.O., and Gorham, J. (1986). The potential for enhancing salt tolerance of wheat and other important crop plants. Outlook on Agriculture, 15: 33-39.