A COM PARISON OF THE SO LVENT ST A BILITY OF THREE GRAM NEG ATIVE O R G ANISM S
Sunil Shrestha
A thesis submitted for the degree of Doctor of Philosophy
at the University of London
Departm ent of Chemical and Biochemical Engineering University College London
ProQuest Number: 10045643
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10045643
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
D e d ic a te d to my F a m ily
AC K N O W LED G EM EN TS
A B S T R A C T
A growing interest lies in the use of biocatalysts in industry to carry out stereo- and regio-specific conversions. M any of these reactions involve poorly w ater-soluble reactan ts and/or products. T his poses p ro b lem s in attain in g high su b stra te and p ro d u c t concentrations in the reactor, and tw o-liquid phase system s can be em p lo y ed in such cases. T he m ain d isa d v a n ta g e o f this ap p ro ach is d e sta b ilisa tio n of the b io c a ta ly st by the organic so lv e n t.
Protection against solvent is afforded by the outer m em brane in Gram negative cells. N aphthalene hydroxylation by Gram negative organisms was studied to compare solvent stability in an aqueous saturated buffer system, and a two-liquid phase system.
T h e n a p h th a le n e d io x y g e n a s e g en e, n a h A , was iso lated and inserted into plasmid pMM B66EH. The new plasmid construct was used to transform E . c o l i JM107 and P . a e r u g i n o s a strains P A C IR and PAC610. P.putida UV4 which expresses toluene dioxygenase activity from chrom osomally encoded genes, was used to complete the array of m icroorganism s studied.
E x p o su re to solvent, both in tw o-liquid phase system s and in solvent saturated buffer, show ed that d ifferen ces in cell outer m em brane composition affected the degree to which cells retained s t a b i l i t y . S o l v e n t h y d r o p h o b i c i t y and p r e s e n c e o f an a q u e o u s i o r g a n ic p h a s e i n te r f a c e , w e re a ls o i m p o r t a n t in determining the rate at which solvent damage occurred.
Loss of stability was shown to be a time dependent phenomenon, with sustained exposure leading to increased loss of stability, even with very hydrophobic solvents.
P A C lR (p S S 2 ) was unable to convert naphthalene at the same rate as P A C 6 1 0 (p S S 2 ). This d iffe re n c e is th o u g h t to be due to d ifferen ces in the extent of interactions betw een com ponents of th e ir o u te r m em b ran e s a ffe ctin g the rate o f u p tak e of the s u b s tra te . P a rtia l p e rm e a b ilis a tio n by so lv e n ts in c re a s e d the activity observed in PA C lR (pSS 2).
I A ctivity retention show n by UV4 w as low er than that of JM 107(pSS2) in hexane i saturated buffer for the first 4hours of biotransform ation, but greater over the
I final 21 hour period. UV4 also retained a greater lev el of activity than JM 107(pSS2)
lover the en tire biotransform ation in tetradecane saturated buffer. PAClR(pSS2) I activity appeared to be stim ulated by the presence of saturating lev els of hexane
and tetradecane over the first 4hours, but declined w ith continued exposure. This I stim ulated le v e l of activity w as still significantly lower than that of JM 107(pSS2).
L I S T O F F I G U R E S
1.1. S y n th e s is of p o l y p h e n y l e n e a n d p i n i t o l f r o t n
b e n z e n e . (1 2 )
1.2. T h e o r tic a l c o n c e n tr a ti o n p ro file s fo r t r a n s f e r of s u b s t r a t e d is s o lv e d in o rg a n ic p h a s e in a t w o -
liq u id p h a s e b io r e a c to r . (1 5 ) 1.3. C lassification of t w o - liq u id p h a s e s y s t e m s b y th e
n a t u r e of th e b io c a ta ly s t a n d t h e p h a s e ra tio . (1 8 ) 1.4. T h e o r e tic a l t w o - l iq u i d p h a s e p ro files: b i o c a ta l y s t
s o lu b ility a n d p h a s e ratio . (1 9 ) 1.5. Toxic e ffe c ts d u e to so lv e n t. (2 2 ) 1.6. T he N -a c e ty lg lu c o s a m in e (NAG) a n d t h e
N-a c e ty l m u r N-a m ic N-acid (NAMA) r e s i d u e s of cell w N-a ll
p e p tid o g ly c a n . (3 0 )
1.7. T h e s t r u c t u r e of teich o ic acid. (3 1 ) 1.8. T h e G ram n e g a tiv e cell w a ll s t r u c t u r e . (3 3 ) 1.9. S c h e m a tic d ia g r a m of th e S f o r m lip o p o ly
-s a c c h a rid e . (3 6 )
1.10.1. Core p o ly s a c c h a r id e s of r o u g h m u t a n t s of
S. t y p h i m u r i u m (Ra-Re). (3 8 )
1.10.2. P a r t ia l s t r u c t u r e of th e l ip o p o ly s a c c h a r id e of
P. ae r u gi no s a PACIR. (3 8 )
1.11. N a p h t h a l e n e d e g r a d a t i o n v i a t h e m e t a - c le a v a g e
p a th w a y . (4 1 )
1.12. D irection of e le c tr o n t r a n s f e r t h r o u g h n a p h t h a l e n e
d io x y g e n a s e . (4 3 )
1.13 LogP: a c tiv ity p ro file f r o m A . s i m p l e x a n d P . p u ti d a
UV4 (4 7 )
2.1. 7 5 m l s t i r r e d t a n k r e a c t o r (5 6 ) 2.2. N a p h t h a l e n e d ih y d r o d io l s t a n d a r d c u r v e (5 8 ) 2.3. C o n v e rsio n of t r y p t o p h a n a n d in d o le to in d ig o (6 3 ) 2.4. S c h e m a tic d ia g r a m illu s t r a ti n g b i o t r a n s f o r m a t i o n in
3.4. P . p u t i d a UV4 g r o w t h in v a r i o u s m e d ia (7 6 ) 3.5. P . p u t i d a UV4 in d o le c o n v e r s io n to indo x y l:
c o m p a r a t i v e a c tiv itie s (7 7 ) 3.6. C o m p a r a tiv e a c tiv itie s of J M 1 0 7 e x p r e s s in g p la s m id s
(pSS2), (pSS3), a n d (pSS4) (7 9 ) 3.7. . C o n v e rsio n of in d o le b y PA C lR (p S S 2 ) a n d
P A C 6 1 0 (p S S 2 ) (8 3 )
3.8. C o n v e rsio n of n a p h t h a l e n e b y P A C lR (p S S 2 ) a n d
P A C 61 0(p S S 2 ) in STRs (8 4 )
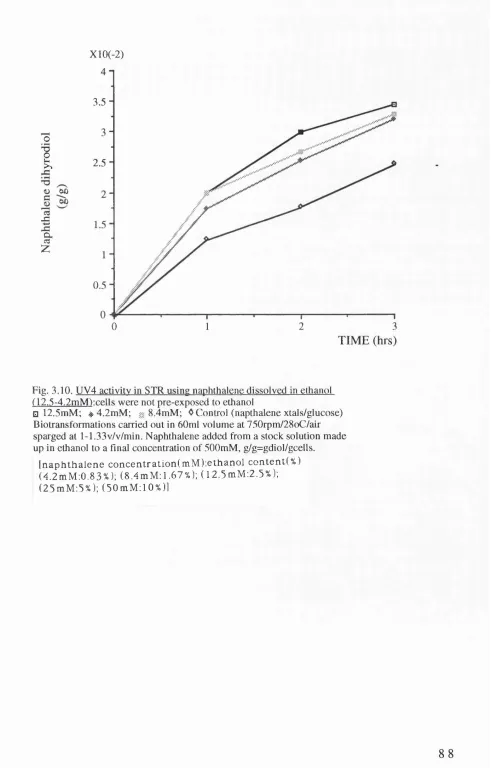
3.9. UV4 a c tiv ity in S IR u sin g n a p h t h a l e n e d is s o lv e d in e t h a n o l ( 1 2 . 5 - 5 0 mM) (8 7 ) 3.10. UV4 a c tiv ity in S IR using n a p h t h a l e n e d is s o lv e d in
e t h a n o l ( 1 2 .5 - 4 .2 m M ) (8 8 ) 3.11. JM 1 0 7 (p S S 2 ) a c tiv ity in STR u sin g n a p h t h a l e n e
d is s o lv e d in e th a n o l( 4 .2 m M - 8 .4 m M ) (8 9 ) 3.12. JM 1 0 7 (p S S 2 ) a c tiv ity a f t e r e x p o s u r e to p e n t a n o l
s a t u r a t e d p h o s p h a t e b u f f e r . (9 1 ) 3 .12.1. JM 1 0 7 (p S S 2 ) % a c tiv ity a f t e r e x p o s u r e to p e n ta n o l
s a t u r a t e d b u f f e r . (9 2 ) 3.13. JM 1 0 7 (p S S 2 ) a c tiv ity a f t e r e x p o s u r e to h e x a n e
s a t u r a t e d b u f f e r . (9 3 ) 3 .1 3 .1 . JM 1 0 7 (p S S 2 ) % a c tiv ity a f t e r e x p o s u r e to h e x a n e
s a t u r a t e d b u f f e r . (9 4 ) 3.14. J M 1 0 7 (p S S 2 ) a c tiv ity a f t e r e x p o s u r e to t e t r a d e c a n e
s a t u r a t e d b u f f e r . (9 5 ) 3 .1 4 .1 . J M 1 0 7 (p S S 2 ) % a c tiv ity a f t e r e x p o s u r e to t e t r a d e c a n e
s a t u r a t e d b u f f e r . (9 6 ) 3.15. UV4 a c tiv ity a f t e r e x p o s u r e to p e n t a n o l s a t u r a t e d
b u f f e r . (9 9 )
3 .1 5 .1 . UV4 % a c tiv ity a f t e r e x p o s u r e to p e n t a n o l s a t u r a t e d
b u f f e r . ( 1 0 0 )
3 .16. UV4 a c tiv ity a f t e r e x p o s u r e to h e x a n e s a t u r a t e d
b u f f e r . ( 1 0 1 )
3 .1 6 .1 . UV4 % a c tiv ity a f t e r e x p o s u r e to h e x a n e s a t u r a t e d
b u f f e r . ( 1 0 2 )
3.17. UV4 a c tiv ity a f te r e x p o s u r e to t e t r a d e c a n e s a t u r a t e d
3 .1 7 .1 . UV4 % a c tiv ity a f te r e x p o s u r e to t e t r a d e c a n e s a t u r a t e d
b u f f e r . (1 0 4 )
3.18. PA C lR (pSS 2) a c tiv ity a f t e r e x p o s u r e to p e n t a n o l
s a t u r a t e d b u f f e r . (1 0 6 ) 3.18.1. PA C lR (pSS 2) % a c tiv ity a f t e r e x p o s u r e to p e n ta n o l
s a t u r a t e d b u f f e r . ( 1 0 7 ) 3.19. PA C lR (pSS 2) a c tiv ity a f t e r e x p o s u r e to h e x a n e
s a t u r a t e d b u f f e r . ( 1 0 8 ) 3 .1 9 .1 . PA C lR (pSS 2) % a c tiv ity a f t e r e x p o s u r e to h e x a n e
s a t u r a t e d b u f f e r . ( 1 0 9 ) 3.20. PA C lR (pSS 2) a c tiv ity a f t e r e x p o s u r e to t e t r a d e c a n e
s a t u r a t e d b u f f e r . ( 1 1 0 ) 3 .2 0 .1 . P A C lR (p S S 2 ) % a c tiv ity a f t e r e x p o s u r e to t e t r a d e c a n e
s a t u r a t e d b u f f e r . ( I l l ) 3.21. JM 1 0 7 (p S S 2 ) a c tiv ity a f t e r e x p o s u r e to a
h e x a n e :b u f f e r 2 - liq u id p h a s e s y s te m (1 1 3 ) 3.22. UV4 a c tiv ity a f t e r e x p o s u r e to a h e x a n e :b u f f e r
2 - liq u id p h a s e s y s te m (1 1 4 ) 3.23. PA C lR (pSS 2) a c tiv ity a f t e r e x p o s u r e to a
h e x a n e :b u f f e r 2 - liq u id p h a s e s y s te m (1 1 5 ) 4.1. A c tiv ity r e t e n t i o n a f t e r e x p o s u r e to h e x a n e s a t u r a t e d
b u f f e r (1 2 9 )
4.2. A c tiv ity r e t e n t i o n a f te r e x p o s u r e to t e t r a d e c a n e
s a t u r a t e d b u f f e r ( 1 3 0 ) 4.3. P r o p o s e d m e c h a n is m of s o l v e n t a c tio n u p o n
th e cell ( 1 3 7 )
L IST OF TABLES
1.1. N a t u r a l ly o c c u rrin g d e g r a d a t i v e p la s m id s (3 9 )
2.1. O b ta in e d p la s m id s (4 9 )
2.2. C o n s tr u c te d p la s m id s (5 0 )
3.1. UV4 g r o w t h a n d a c tiv ity in v a r i o u s m e d ia (7 4 ) 3.2. Specfic a c t iv i t ie s f r o m JM 1 0 7 (p S S 2 ), (pSS3), a n d
(pSS4) (7 8 )
A B B R E V I A T I O N S
A p a m p ic illin
Cb c a r b e n ic illin
DNA d eo x yribonucleic acid
EDTA e th y le n e d ia m in e te tr a a c e tic acid IPTG i s o p r o p y l - p - D - th i o g a la c t o p y r a n o s i d e
k b p kilobase pairs
Kn k a n a m y c i n
LPS lip o p o ly s a c c h a r id e
MNNG N - m e t h y l - N ’- n i tr o - N - n i t r o s o g u a n id i n e PAGE po ly acry lam id e gel electro p h o resis
SDS sodium dodecyl sulphate
STR stirred tank reactor
S u p erscrip t r indicates resistance TEE tris - b o r a te - E D T A
TE tris-EDTA
T ris t r i s ( h y d r o x y m e t h y l ) a m i n o m e t h a n e UV u l t r a v i o l e t
XGal 5 - b r o m o -4 - c h l o r o - 3 - i n d o l y l- p - D - g a l a c to s id e
A . s i m p l e x : a r t h r o b a c t e r s i m p l e x E.coli: Esch eri ch ia coli
S.a ur e us : S t a p h y l o c c u s a u r e u s
S . t y p h i m u r i u m : S a l m o n e l l a t y p h i m u r i u m P. putida: P s e u d o m o n a s p u t i d a
CONTENTS P a g e
i Title page (1)
il Abstract (3)
iii List of figures (4)
iv List of tables (6)
V Abbreviations (7)
vi Contents (8)
1. INTRO DUCTIO N ( 1 1 )
1.1 BIOCATALYSTS (11)
1.1.1. Biocatalysts in industry (11)
1.1.2. Advantages of using biocatalysts (11)
1.2 Two-liquid phase systems (13)
1.2.1. Introduction to tw o-liquid phase system s (13) 1.2.2. Classification of two-liquid phase systems (14) 1.2.3. Advantages of two-liquid phase system s (17) 1.2.4. D isadvantages of two-liquid phase system s (20)
1.2.5. Solvent selection criteria (20)
1.3. Effects of solvents on biocatalysts (21)
1.4. Prediction of solvent stablity (24)
1.5. Bacterial cell walls (26)
1.5.1. Gram positive cell wall (26)
1.5.2. Gram negative cell wall (27)
1.6. N ap hthalene hydroxylation (37)
1.6.1. N aphthalene degradation (37)
1.6.2. Plasmid encoded genes involved in naphthalene
d e g r a d a t i o n (39)
1.6.3. N aphthalene dioxygenase (40)
1.6.4. Toluene dioxygenase (42)
1.6.5. Regulation of the n a h and sa l opérons (44)
1.7. Previous work (45)
1.8. Aims of research program m e (46)
2. M ATERIALS AND METHODS ( 4 9 )
2.1. MATERIALS (49)
2.1.1. Organism s (49)
2.1.2. Reagents (50)
2.3. METHODS (57)
2.3.1. DNA manipulations (57)
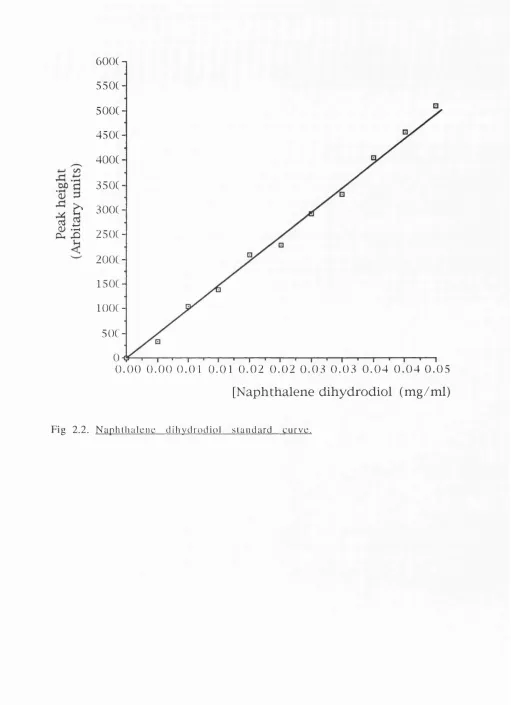
2.3.2. N aphthalene dioxygenase activity m easurem ents (62)
2.3.3. F erm en tatio n s (64)
2.3.4. Solvent exposure studies (65)
3. R ESU LTS ( 6 8 )
3.1. GENETIC MANIPULATIONS (68)
3.1.1. Restriction enzyme map of pQR113 (68) 3.1.2. Construction of plasmids p S S l, pQR155, and
PQR156. (68)
3.1.3. Construction of plasmids pSS2, pSS3, and pSS4 (70)
3.2. GROWTHS AND ACTIVITIES (70)
3.2.1. UV4 growth in complete and m inim al m edia (70) 3.2.2. UV4 dioxygenase activity com parisons (74) 3.2.3. JM107 growth in nutrient broth (75) 3.2.4. Comparisons of dioxygenase activity from JM107 harbouring plasmids pSS2, pSS3, and pSS4 (75) 3.2.5. E ffect of cell concentration on dioxygenase activity of
J M 1 0 7 (p S S 2 ) (80)
3.2.6. P A C IR and PAC610 growth in nutrient broth (81) 3.2.7. PA C lR (pSS 2) and PAC610(pSS2) conversion of
i n d o le (81)
3.2.8. PA C lR (pSS 2) and PAC610(pSS2) conversion of
n a p h t h a l e n e (81)
3.3. USE OF NAPHTHALENE DISSOLVED IN ETHANOL (82) 3.3.1. Explanation for use of ethanol (82)
3.3.2. UV4 (85)
3.3.3 JM 107(pSS2) (86)
3.4. SOLVENT EXPOSURE STUDIES (90)
3.4.1. Solvent choice (90)
3.4.2. E xposure to solvent saturated buffer (90) 3.4.3. Exposure to hexane:buffer two-liquid phase
s y s t e m s (112)
4. GENERAL DISCUSSIO N ( 1 1 6 )
4.1. DIOXYGENASE ACTIVITY LEVELS (116)
4.1.1. Variations in UV4 dioxygenase activity (116) 4.1.2. JM 107 dioxygenase activity variation with plasmids
4.1.3. Dioxygenase activity inhibition of JM 107(pSS2) (118) 4.1.4. Dioxygenase activities of UV4 and JM 107(pSS2) using
nap h th alen e in ethanol (118)
4.1.5. Dioxygenase activity comparisons o f P A C lR (p S S 2 ) and
P A C 6 1 0 (p S S 2 ) (122)
4.2. SOLVENT EXPOSURE STUDIES (124)
4.2.1. Factors influencing cell disruption (124)
4.2.2. Cell structure comparisons (131)
4.3. PROPOSED MECHANISM OF SOLVENT ACTION UPON WHOLE
CELLS. (135)
4.4. IMPLICATIONS FOR INDUSTRY (136)
5. C O N C L U SIO N S ( 1 3 9 )
6. FU T U R E W O R K (140)
7. A P P E N D I X (143)
1. I N T R O D U C T IO N
1.1. BIOCATALYSTS
1 .1 .1 . Biocatalysts in industry
Im proved und erstan d in g o f m icro b ial b io ch em istry and genetics has increased the potential for the use of both whole cells and isolated enzym es as biological catalysts (biocatalysts) in industrial processes. In addition to their ability to pro d u ce a num ber of u n iq u e c o m p o u n d s , th ey m ay also be u sed to s y n th e s iz e com pounds traditionally produced by ch em ical m eans. E xam ples o f products in which biocatalysts play a role in synthesis include, 1,2-chloropropionic acid (IC I), (S )-N ax o p re n ( ( S ) - 6 - m e t h o x y - a - m e th y l-2 -n a p h th alen e ac etica cid ) (In tern atio n al B io sy n th etics, BV- IBIS) [Pryce and Roberts,(1990)], polyphenylene (ICI), pinitol (ICI) [Ballard et al.(1983)1 ( F i g l . l . ) , and 3-fluoroveratole [Johnston e t a i,(1 9 8 7 )].
1 .1 .2 . A dvantages of using biocatalysts Regio- and stereospecificitv:
R eactions catalysed by isolated enzym es and w hole cells may co n fer strict regio- and stereospecificity [Ziffer et a l. ( 1 9 7 7 ) : G ibson and S u b ram anian,(1984); Brand et al.(1992)1 and so are p o t e n t i a l ly f a v o u r a b le fo r use in th e p h a r m a c e u ti c a l and a g ro c h em ica l in d u stries. N ot only does this c o n tro l the exact nature of the product, but in doing so, also reduces the amount of b y -p ro d u c t form ed.
Mild operating conditions:
B io c a ta ly s ts w ork o p tim a lly u n d e r r e la tiv e ly m ild o p e ra tin g c o n d itio n s of tem perature, pressure and pH, reducing both the o p eratin g costs and the likelihood of un w an ted side reactions o c c u rrin g .
n
Benzene
P . p u t i d a
cis -Benzene glycol
C H C H3O
P o l y p h e n y l e n e ( + )-Pinitol
Reduced reaction stages:
Com pared to the chem ical conversion processes, the biologically c ataly sed o p eratio n s may be carried out m ore d irectly , often completing the conversion in a single step reaction [Johnston et al. (1987); Ballard et al.(1983)l. Loss of intermediates at each stage of the conversion process is thus reduced, as is the possibility of the occurrence of unwanted side reactions.
Abilitv to recognise a range of related compounds:
B iocatalysts are able to recognise and utilise a range of related com pounds, often differing m arkedly in their structure, yet still m aintain the same high degree of specificity [Ley et a l. ( 1 9 8 7 ) : Johnston and Reganathan,(1987); Gibson et al.(1989)1.
G ibson and colleagues (1990) identified 25 d ifferen t com pounds which could be recognised by the toluene dioxygenase of P . p u t i d a strain F39/D, and over 30 different compounds which the enzyme cloned into an E.co li strain, could utilise.
W h ilst the biotransform ation process can be carried out during the ferm e n ta tio n its e lf [W ith o lt et aL(1 9 9 0 ): F a v r e - B u lle e t a l,(1 9 9 1 )], separation of the two stages may often be desirable. W here a possibility exists of the substrate or p ro d u ct affecting c e llu la r grow th [Favre-B ulle eJL_al,(1991)], isolation of the two stages is beneficial to the overall process. This also allows for the g r o w t h an d b i o t r a n s f o r m a t i o n p h a s e s to be o p t i m i s e d in d iv id u a lly , w ith o u t co m p ro m ise, with c ells h arv ested at the stage where they have optim al enzym ic activity. Finally, if the b io tran sfo rm atio n is carried out over a relativ ely short period, new cell growth is unlikely to occur, thus the necessity for sterile conditions during the biotransform ation process is removed.
1.2. TWO-LIQUID PHASE SYSTEMS
1 .2 .1 . Introduction to two-liquid phase systems
M any biological reactions of interest employ substrates which are only sparingly soluble in water. Yet an aqueous environm ent is essential for biocatalytic activity, in both iso lated enzym e and
w hole cell systems; dried A . s i m p l e x cells were observed to be u n ab le to c ataly se stero id c o n v e rsio n , but a ctiv ity could be resto re d upon reh y d ra tio n of the cells [H o c k n u ll,(1 9 8 9 )]. For isolated enzym es, w ater m olecules are required to m aintain non- co v alen t interactions essential for activity. T hese include ionic bonds, van der W aals bonds, and hydrophobic interactions. The a m o u n t o f w ater req u ired appears to be d e p e n d e n t upon the nature of the enzyme. Klibanov (1989) proposed that only a single m onolayer of water around the enzym e was required for activity, and that a -c h y m o try p s in could retain activity in various solvents due to the presence of 50 mole equivalents of water bound to only the charged groups of the protein [Zaks and Klibanov,(1988)].
W ater m ay also be used as one of the active reagents in the rea c tio n , fo r e x am p le du rin g h y d ro ly sis, or to d isso lv e any cofactors present.
1 .2 .2 . Classification of two-liquid phase systems
Classification of two-liquid phase systems is im portant in order to u nderstand the m echanism s of solvent d istrib u tio n betw een the aqueous and organic phases. This allow s the m ass tran sfer of substrate and product in the reactor to be studied, factors that need to be taken into account when designing specific two-liquid phase b io tran sfo rm atio n system s.
A c la ss ific a tio n sy stem based upon c o n c e n tra tio n p ro file s for poorly w ater-soluble substates dissolved in the organic solvent, has been proposed by Lilly and W oodley (1985). Three theoretical concentration profiles dependent upon solubility of the biocatalyst in the aqueous phase and the aqueousiorganic phase ratio can be distinguished (Fig 1.2.).
Profile A
Organic Phase Biocatalyst (soluble)
A
Organic Phase Aqueous Phase Biocatalyst (insoluble)
B
Organic Phase Biocatalyst (insoluble)
F ig .1.2. Theoretical concentration profiles for transfer o f substrate
dissolved in organic phase in a two-liquid phase bioreactor (from Lilly and Woodley, (1985)).
Interface and film shown by a continuous line and a dashed line, r e s p e c t iv e ly .
Profile B
W hen insoluble biocatalysts are used (im m obilised enzym es, free or immobilised whole cells), a further set of partition coefficients and concentration gradients may be introduced by the biocatalyst and its support, in addition to those created at the liquid:liquid in te r f a c e .
Profile C
If the phase ratio is altered such that the level o f aqueous phase is decreased, so that it no longer forms a discrete phase between the organic phase and the insoluble b io cataly st, then in effect, only an aqueous film is retained around the b io cataly st. The concentration gradient produced by the aqueous phase becom es less significant, and the profile depicted by profile C is predicted to be observed.
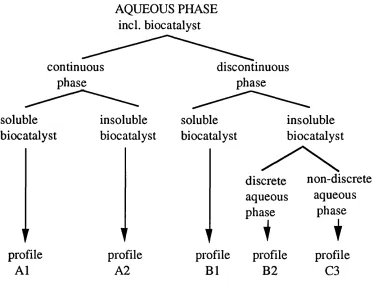
D erived from the reacto r concentration p rofiles, a classification sy stem based upon the b io cataly st d istrib u tio n and the phase ratio, can be distinguished (Figs 1.3. and 1.4.). The biocatalyst is assumed to be p resent in the aqueous phase either in a soluble (free enzyme), or insoluble form (free or imm obilised whole cell, or im m obilised enzym e). The aqueous phase can exist either as the continuous phase, a discrete discontinuous phase, or as a non discrete phase.
Use of water-miscible organic solvents to increase the solubility of poorly w ater-m iscible substrates, has been studied [Sonom oto e t al,(1979;198/f); Y okozeki et al.(l982)1. The main disadvantage of this system is that high levels of dissolved solvent in the aqueous phase can result in a loss of operational stability and reduced activity. W hen high concentrations of dissolved solvents are used, even immobilisation of the biocatalyst may not be able to prevent solvent disruption of the biocatalyst [B utler,(1979)]. Furtherm ore, a d d itio n o f c o -s o lv e n t is unable to a lle v ia te the p ro b le m s a sso c iated w ith su b stra te and /o r p ro d u c t in h ib itio n , and m ay exacerbate the situation by m aintaining close p roxim ity o f these com pounds with the biocatalyst.
biotransform ations. The organic phase acts as a reserv o ir and/or sink into w hich the su b strate an d /o r p ro d u c t d isso lv e s. T he aqueous layer retains the b io ca ta ly st and any h y d ro p h ilic c o factors, acting as the site in which the reaction occurs. This system m ay enable b io tran sfo rm atio n to occur w ith o u t the d e leterio u s effects associated with using a water-miscible solvent.
1 .2 .3 . Advantages of two-liquid phase systems.
i) T he a d d itio n o f so lv e n t to fo rm a d is c r e te p h a se can significantly increase the concentration of substrate present in the re a c tio n v essel, th ereb y red u cin g the v o lu m e of the re a c to r necessary to form a given am ount of product. The am ount of dissolved substrate can reach high levels; 170g/L of cholesterol d isso lv e d in carbon tetrac h lo rid e was re p o rte d to have been obtained by B uckland and co-w orkers (1975). A lternatively, the substrate m ay be used to form the organic phase [Brazier e b
al ,(1990); Harbron et al.(1986)1.
Sim ilarly, the amount of product which can be formed, may also be increased as the reaction equilibrium is altered.
ii) W here the su b strate is a solid at re a c tio n te m p e ra tu re s, solubilisation by the solvent may make it more accessible to the biocatalyst [Ceen et al.(1987)1.
iii) Hydrophobic products formed are readily p artitioned into the organic layer, reducing the possibility of su p ersatu ratio n in the aq u eo u s p h ase, and any c ry s ta llisa tio n o f p ro d u c t u pon the biocatalyst surface that may arise.
iv) W h ere high levels of substrate or p ro d u c t m ay lead to b io cataly st inhibition, partitioning away from the aqueous phase can lim it these detrimental effects.
v) F in ally , d o w n stream p ro ce ssin g m ay be g reatly sim p lified . Isolation of the organic phase in which p ro d u ct and unreacted substrate accumulate, may ease the isolation and recovery of the b io c a ta ly s t.
AQUEOUS PHASE incl. biocatalyst
continuous phase
discontinuous phase
soluble biocatalyst
profile A1
insoluble biocatalyst
t profile
A2
soluble biocatalyst
insoluble biocatalyst
discrete non-discrete aqueous aqueous phase
1 phase1
1'
i
profile profile profile
B1 B2 C3
A l B1
A2
° °; °
B 2
o
OŒ
D
Fig 1.4. Theoretical two-liquid phase profiles: biocatah si solu b ility and phase ratio (Adapted from Lilly and Woodley, (19 8 5 ))
■ Insoluble biocatalyst; H Organic phase; O Aqueous phase
Also, reco v ery of p ro d u ct from the reactio n m ed iu m m ay be sim plified by virtue of the low boiling points o f m any organic s o lv e n ts .
1.2.4. D isadvantages of two-liquid phase system s.
The m ain disadvantage of tw o-liquid phase bio cataly tic systems res u lts fro m the d e n a tu rin g e ffe c t of the so lv e n t upon the biocatalyst. This can occur either at the liquidiliquid interface, or within the aqueous phase itself, due to the presence of dissolved solvent. The actual mechanism(s) by which inhibition occurs is not clearly understood, and will be discussed later (section 1.3.).
1 .2 .5 . Solvent selection criteria
The final selection of solvent to be em ployed m ust take into consideration a num ber of different factors [Lilly,(1982)]:
i) Reactant/product solubility in the the solvent.
ii) Reactant/product partitioning between the two phases.
T h e n e ed to p a rtitio n s u b s tr a te s a n d /o r p r o d u c ts e x e rtin g in h ib ito ry effects upon the b io c a ta ly st, has been m en tio n ed . C om plete partitioning of product into the organic layer is also desirable for dow nstream processing. How ever, high affinities of the su b stra te for the so lv e n t ph ase m ay n o t n e c e ssa rily be b en eficial, if this hinders su b strate tra n s fe r into the aqueous p h a s e .
iii) Biocatalyst inhibition by organic solvent.
The solvent itself can exert inhibitory effects upon the biocatalyst. However, these effects can be minimised by im m obilisation of the b io cataly st to reduce d énaturation at the liq u id iliq u id interface [Hocknull and Lilly,(1990)].
The solvent to be used m ust be safe with respect to its toxicity and flammability. This m ust also be taken into consideration when deciding upon methods of disposal.
vi) Solvent cost.
1.3. EFFECTS OF SOLVENT ON BIOCATALYSTS
The actual m echanism (s) by which solvents affect biocatalysts is not clearly understood, with a wide range o f effects observed [Lilly et al.(1987)1. The resultant effects are dep en d en t upon a num ber of factors including the nature of the cell and solvent, the mode of presentation of solvent, the period of exposure, and also the ag itatio n rate. B ar (1986), d istin g u is h e s b etw een so lv e n t effects resulting from the presence of a liq u id iliq u id interface (phase toxicity), and those caused by solvent dissolved in the aqueous solution (m olecular toxicity) (F ig l.5 .).
Only a few of the negative effects of solvents on the biocatalyst are illustrated, with a greater range of effects know n to occur, both at the m orp h o lo g ical and the p h y sio lo g ic al levels [Lilly, (1982); Lilly et al.(1987)1.
Loss of cell viabilitv
Loss of viability of cells exposed to a range of organic solvents has been reported in both E. coli. and P s e u d o m o n a s [Jackson and D e M o ss,(1 9 6 5 ); M oses and R ic h a rd so n ,(1970); De Sm et et al. (1 9 8 3 )].
Loss of cell morphologv
Several changes in cellular m orphology due to solvent dam age have been observed by electron m icroscopy. Solvent treated cells show ed c y to p lasm ic shrinkage [Jackson and De M o s s ,(1 9 6 5 )], chrom osom al displacem ent [De Smet et al.(1978)1 and m em brane d isru p tio n [Favre-B ulle et al.(1991): D eSm et et al. ( 1 9 7 8 : 1 9 8 3 ) 1 . w ith d isru tio n o c cu rin g in both the c y to p la s m ic and o u te r m e m b r a n e s
Loss of intracellular proteins
In tact cells w ere o b served to lose in tra c e llu la r p ro tein s in a selective m an n e r [D eSm et et al.(1978)1. M a la te d e h y d ro g e n a s e was released in toluene treated E . c o l i , bu t o th er c y to p la s m ic p r o t e i n s , g l u c o s e - 6 - p h o s p h a t e d e h y d r o g e n a s e and g l u ta m a te dehydrogenase, were retained within the cell. This m ay reflect a p o s s ib le p e r i p h e r a l l o c a tio n f o r m a la te d e h y d r o g e n a s e , or selective release by the peptidoglycan or outer m em brane layers.
Loss of RNA
RN A loss was shown to be independent of the level of solvent presen t (1 .5p l/m l to SOpl/ml), with up to 85% of the total cellular R N A b ein g e x tr a c te d in to the m ed iu m . T h is m ay lead to im p a irm e n t o f en zy m e sy n th esis req u ired fo r m a in te n a n c e and repair of the cell [Jackson and DeMoss,(1965)].
SOLVENT
Molecular Toxicity Enzyme inhibition Protein dénaturation Membrane modification
-membrane expansion
-structure disorder -permeability change -etc.
Phase Toxicitv Extinction of nutrients Disiiiption of cell wall
-extinction of outer cellular components
Limited access to nutrients -cell attraction to interface -emulsion formation -cell coating
F i g l .5 . T oxic effects due to solvent
a) d is s o lv e d in the aqueous medium (m olecu lar toxicity);
b) forming a distinct second phase (phase toxicity),
Loss of lipopolysaccharide (LPS) and phospholipid
Exposure of cells to toluene was observed to result in rem oval of up to 41% o f o u ter m em b ran e L P S . In c o n tra st, the outer m em brane p h o spholipid rem ained relativ ely u naffected [De Sm et et a l .(1978)1. D e s p ite th is, the o u t e r m e m b r a n e a p p e a re d relatively intact, and the difference in the amount of the two lipid species lost m ay reflect the m ore exposed location o f the LPS com pared with the phospholipids.
Loss of selective perm eabilitv
The role of the outer m em brane includes selectively limiting the flux o f m ateria ls betw een the cell and its e n v iro n m en t. M any solvents can disrupt this function. Cells treated with toluene were unable to concentrate C ^ ^ -th io m e th y lg a la c to sid e w ithin the cell, and in sta n ta n e o u s loss o f this c o m p o u n d was o b serv ed upon exposure to the solvent [Jackson and D eM oss,(1965)]. Sim ilarly, increased su scep tib ilty to hydrophobic a n tib io tics was observed following treatm ent of cells with polymyxin B nonapeptide, due to disorganisation of the outer m em brane structure by the antibiotic [Vaara et al.(1984)1.
Loss of substrate uptake
Non-com petitive inhibition by solvents has been reported to affect sugar uptake m echanism s [Loureiro-Dias and Peinado,(1982); Leao and van U nden,(1982)]. Inhibition in these cases was attributed to a lte r a t io n s in the lip id e n v ir o n m e n t d i s r u p t in g th e c lo s e l ip i d : p r o t e i n i n t e r a c t i o n s r e q u ir e d to m a i n t a i n th e a c tiv e conformation of the enzymes. The rate of loss of activity appeared to be correlated with the lipophilicity of the solvent used.
Loss of m em brane associated functions
An intact m em brane to generate an electro n tran sp o rt chain is essential for the formation of ATP and NADH. The NADH co-factor recycling step was shown to be the p rim ary targ et of organic solvent disruption of steroid dehydrogenation in A . s i m p l e x , rather than the a ctu al d e h y d ro g e n a s e e n z y m e its e l f [H o c k n u ll and L illy ,(1988)]. Sim ilarly, (N a+/K +)-A T P ase activity was lost upon addition of alcohols, due to disruption of the ability to form a
m em b ran e pro to n p o ten tial. As a result, res p ira tio n d e p en d e n t lactose and proline transport functions were inactivated [Patel e t a l,(1 975)].
Lim ited access to nutrients
Aggregation of cells at the liquidiliquid interface and form ation of emulsions may hinder the uptake of nutrients. In addition, coating of cells with an organic layer can lead to blockage of nutrient diffusion into the cell [Cho and Shuler,(1986)].
1.4. PREDICTION OF SOLVENT STABILTY
The wide array of effects produced by solvents upon biocatalysts m akes the correlation of stability and activity with the nature of solvent more d ifficu lt. F o r exam ple, Crem onesi and co-w orkers (1975) observed that P -h y d ro x y s te ro id d e h y d ro g e n a s e sh o w ed a significant decrease in activity in carbon tetrachloride, yet still rem ain ed stable.
V a r io u s w o rk e rs h a v e lo o k ed at th e r e la ti o n s h i p b e tw e e n b io c a ta ly st stab ility and so lv e n t p o larity . B rin k and T ra m p e r ( 1 9 8 5 ) r e p o r t e d an a s s o c i a t io n b e tw e e n s o l v e n t p o l a r i ty (m ea su red in term s of the H ild e b ra n d c o e f f ic ie n t (Ô)), and m olecular weight of the solvent, with the biocatalytic activity of free and im m obilized cells. Low activities were observed when solvents of high polarity (ô > 8 ) and low m olecular weight (less than 200) w ere em ployed. Sim ilarly, L aane and co -w o rk ers (1985), looking at work carried out by various groups using a range of biocatalytic systems (free and im m obilised cells and enzym es), in the p resen ce of a variety of organic so lv e n tiaq u e o u s system s, suggested that polarity of the solvent could be more satisfactorily d efin ed in term s of logP, where logP is re p re se n te d by the e q u a tio n :
LOG P= [solvent] in octanol [solvent] in water
A co rre latio n betw een the logP value of the so lv en t and the biocatalytic activity retained was found in w hich, solvents with logP values greater than 4 resulted in m inim al dam age to the biocatalyst, and those with LogP values less than 2 caused total loss o f activity. This correlation has been used successfully by several workers. However, discrepancies can still occur; Hocknull and Lilly (1989) found that solvents of logP greater than 4 still caused inactivation of the im m obilised bio cataly st, but that the rate of loss of activity increased with increasing water miscibility of the solvent.
A lthough the results of L aa n e ’s group may be used as a general guideline in the selection o f solvents, it does not give us an understanding of the m echanism (s) involved in the disruption of biocatalysts. A closer inspection o f c ell:so lv en t in teractio n s is required in order to elucidate this problem.
1.5. BACTERIAL CELL WALLS
B acterial cells can be classified upon the structure o f their cell walls, and the resu ltan t ability to take up certain dyes. G ram positve and Gram negative cells share many features such as the p e p tid o g ly c a n la y e r and the te ic h o ic a c id s, a lth o u g h the proportions of these components found in the two classes of cells d iffer. G ram n e g ativ e c ells are fu rth e r d istin g u is h e d by the p r e s e n c e o f a s e c o n d m e m b r a n e s t r u c t u r e , d i f f e r e n t in c o m p o sitio n to the c y to p lasm ic m em b ran e , and o v e rly in g the p ep tid o g ly ca n layer.
1 .5 .1 . Gram positive cell walls
The Gram positive cell wall is m orphologically distinct from its Gram negative equivalent. The m ajor com ponent of the cell wall is the peptidoglycan, contributing up to 50% of the total cell wall w e ig h t, with the re m a in d e r being m ade up of a v a rie ty of p o l y m e r s .
The Peptidoglycan
Teichoic acids.
T e ic h o ic acids are the m ain p o ly m e rs a s s o c ia te d w ith the p e p tid o g ly can in G ram positive bacteria. T hese co m p o n en ts are the m ajor cell surface antigens of the Gram positive cells, and may also play a role in the movement of ions into the cells.
Teichoic acids consist of glycerol or ribitol phosphate units joined by p hosphodiester linkages, with up to 40 such units linked to form the polymer. Sim ilar su g a r-1-phosphate units are also found in some organisms which, although not strictly teichoic acids, are closely related in terms of their properties and functions (Fig. 1.7). The teichoic acids are joined to the peptidoglycan frame via a linkage unit which joins the teichoic acid chain to the 6' position of the muramic acid residue of the peptidoglycan.
T eichuronic acids.
T e ic h u r o n ic acid s, lik e the te ic h o ic a c id s, fo rm a lin e a r p o ly s a c c h a r id e b e a rin g u ro n ic acid r e s id u e s . T h ey are n o t p h o sp h o ry la te d , but p robably bind to the p e p tid o g ly ca n via a single pho sp h o d iester bond.
1 .5 .2 . Gram Negative cell wall
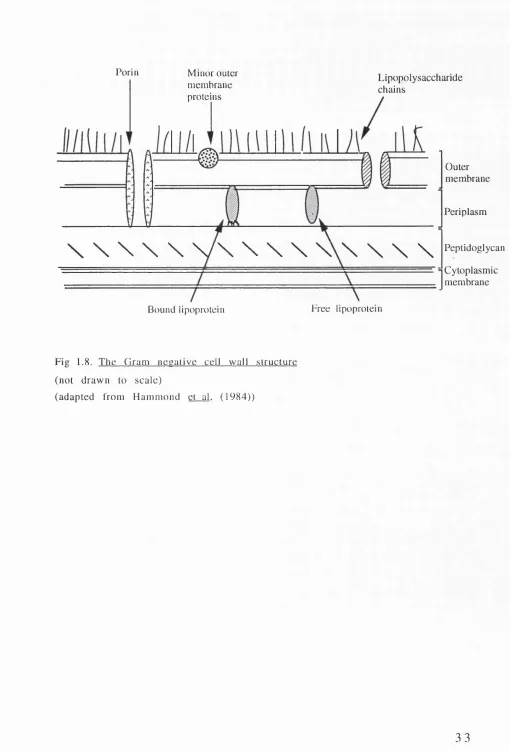
T ransverse sections o f cell walls from G ram positive and Gram n e g a tiv e o rg an ism s show sev eral m a jo r d if f e r e n c e s in th eir stru ctu res. In G ram negative cells, the p e p tid o g ly c a n layer is sig n ific a n tly sm aller, but reta in s its ro le in m a in ta in in g the integrity of the cell. In addition, overlying the peptidoglycan layer is a second m em b ran e stru ctu re co m p o sed o f a lip id /p ro te in bilayer, the outer m em brane (Fig 1.8.). A ssociated with the outer m e m b ra n e are a series of novel p r o te in s, and the co m p lex am p h ip ath ic LPS m o lecu le. The p e rip la s m o ccu p ies the space betw een the inner and outer m em branes. This region contains a nu m b er o f enzym es associated with tran sp o rt, m o d ificatio n and d e g ra d atio n functions.
The p e rip lasm
T he p e r ip la s m d e fin e s the sp ace b e tw e e n the o u te r and cytoplasmic membranes. Estimates of the volume occupied by the
e U l
p e r i p l a s m r a n g e f r o m 5% [ D e c a d , ( l 9 7 6 ) ] to 4 2 % [ S t o c k , ( 1 9 7 7 ) ] d e p e n d e n t u p o n t he g r o w t h p h a s e o f t he c e l l [ D e c a d , ( 1 9 7 6 ) ] .
A l a r g e p e r c e n t a g e o f t h e t o t al c e l l u l a r p r o t e i n c o n t e n t is l o c a t e d in t h e p e r i p l a s m , w i t h t h e a c t u a l a m o u n t d e p e n d e n t u p o n g r o w t h c o n d i t i o n s a n d s t r a i n t y p e [ O x e n d e r a n d Q u a y , ( 1 9 7 5 ) ] . A n u m b e r o f d i f f e r e n t e n z y m i c f u n c t i o n s a r e f o u n d in t h i s r e g i o n , a n d t h e s e m a y be c l a s s i f i e d i n t o t h r e e g r o u p s :
i) B i n d i n g p r o t e i n s ;
t h i s g r o u p o f p r o t e i n s m e d i a t e s t r a n s p o r t o f s o l u t e s , i n c l u d i n g s u g a r s a n d a m i n o a c i d s , to c y t o p l a s m i c m e m b r a n e p e r m e a s e s , w h e r e t h a y c a n t h e n be t r a n s f e r r e d i n t o t h e c y t o p l a s m .
ii) H y d r o l y t i c e n z y m e s ;
t h e h y d r o l y t i c e n z y m e s c a t a l y s e t h e d e g r a d a t i o n o f a n u m b e r o f m e t a b o l i s a b l e c o m p o u n d s u n a b l e t o t r a v e r s e t h e c y t o p l a s m i c m e m b r a n e d u e to t h e i r s i z e o r c h a r g e . P a r t i a l d e g r a d a t i o n c a n t h e n a l l o w t h e s e c o m p o u n d s to b e t r a n s l o c a t e d i n t o t he c y t o p l a s m .
iii) D e t o x i f y i n g e n z y m e s ;
c e l l s a r e a b l e t o d e g r a d e o r m o d i f y a n u m b e r o f a n t i m i c r o b i a l a g e n t s in t h e p e r i p l a s m . A c c u m u l a t i o n o f t h e s e e n z y m e s in t h i s r e g i o n p r o d u c e s a c o n c e n t r a t i n g e f f e c t , l o w e r i n g t h e a m o u n t o f e n z y m e r e q u i r e d to c o m p l e t e d e t o x i f i c a t i o n . C o m p a r a t i v e e n z y m e s p r o d u c e d b y G r a m p o s i t i v e o r g a n i s m a r e s e c r e t e d i n t o t h e m e d i u m , so t h a t a g r e a t e r a m o u n t o f t h e s e e n z y m e s a r e r e q u i r e d to b r i n g a b o u t t he s a m e e ff ect .
iv) D e o x y r i b o n u c l e o s i d e c a t a b o l i s i n g e n z y m e s a n d n u c l e a s e s ;
a n u m b e r o f e n z y m e s in t h e p e r i p l a s m a r e i n v o l v e d in t h e m o d i f i c a t i o n o f d e o x y r i b o n u c l e o t i d e s a n d r i b o n u c l e o t i d e s .
T h e o u t e r m e m b r a n e
T h e o u t e r m e m b r a n e s e r v e s a v a r i e t y o f f u n c t i o n s i n c l u d i n g :
m o le c u la r w e ig h t h y d ro p h ilic m o le c u le s can p ass; p ro v id in g re c e p to r sites fo r b a cterio p h ag e and b a cte rio c in s; d e te rm in in g strain and species specificity of the cell; p ro d u cin g the m ain endotoxic effects associated with the cell.
The outer m em brane is m ade up of three main components: the phospholipids; the proteins; and the lipopolysaccharides.
i) Phospholipid.
T h e m a in s p e c i e s o f p h o s p h o l i p i d i d e n t i f i e d in th e E n tero b actericeae is p h o sp h atid y leth an o lam in e, with low er levels o f p h o s p h a tid y lg ly c e r o l and c a r d io lip in a lso p re s e n t. F r e e z e fra c tu re studies in d ic ate th at the p h o sp h o lid s are d istrib u te d asym etrically in the outer m em brane, with the LPS co m p o n en t f o u n d e x c l u s i v e l y on the o u t e r l e a f l e t [ M u h l r a d t and Golecki,(1975); Van Gool and Nanninga,(1971)]. It is thought that the o u te r le a fle t m ay be c o m p o s e d e n tire ly o f L P S , w ith phospholipids forming the inner leaflet, or that small patches of pho sp h o lip id may exist together with LPS in this outer leaflet [Nikaido and Vaara,(1985)].
i i) P r o te in s L p o / O m p I
The m ost abundant protein isolated from the outer m em brane of E.coli is Braun's lipoprotein [Braun,(1975)]. It can exist either in a free form, or covalently bound to the peptidoglycan [Inouye e t a l , (1972)]. This protein is thought to anchor the outer m em brane to the p e p tid o g ly c a n . M u ta n ts d e fe c tiv e in this lip o p ro te in d isso ciate their outer m em brane when treated with E D T A and detergents, and form vesicles at their surface due to the reduced stability of the m em brane [Suzuki et__al,(1978)]. An a n alo g o u s protein has been identified in P . a e r u g i n o s a . Protein I shows a n u m b er of sim ilarities to B raun's lip o p ro tein , in clu d in g sim ilar m obility in SD S-polyacrylym ide, and the presen ce of covalently b o u n d g l y c e r o l and f a t t y a c id r e s i d u e s [ M iz u n o an d K a g ey a m a,(1 9 7 8 )]. In ad d itio n , sim ila rités in the am in o acid co m p o sitio n and secondary stru ctu re, as well as stretch es of sequence hom ology betwen the two proteins, have been reported [Cornelis et al.(1989)1.
Chain 1 Chain 2
CH20H
NAMA
CH20H
“0 0
NHC0CH3 NHC0CH3
CH3-CH-C0
—— mtmmm mm — mmb bbm mbm «bm mm
NH
L-alanine CH3-CHI
CO
NAG — 0 — NAMA— 0 —
D-glutamic acid
MM* MMM «MM «MM M »M M«M
NH I
CH-COOH I
CH2 I CH2
I CO
^^M MM» MM# MM« M^M ^^M «MM^ ^^M MM MM* ^^M MM ^^M
NH
S * r r - Ç H -C H 2-C H 2-C H 2-C H
CO
MBM MM» MM »MM M^ MM *^M ^^M MM #^M ^^M
NH
I
D-alanine COOH- CH CH3
COOH
NH2
/
L-ala
D-glu
m -dapa
D-ala
r \
1) O f c Œ
RO
-0.
CH2<
y
-,0CH2 RO"
-CH2
y
\ )
-OCH2 R 0
--c m n
R=H, alanyl, glycosyl
2) CH2OH
CH2O
OCH2 OCH2
CH2O CH2O'
R= a or p N-acetyl glucosaminyl Ala= alanine
Fig 1.7 The structure of teichoic acid
consisting o f 1) glycerol phosphate, or 2) ribitol phosphate units, (adapted from Hammond et al. (1984))
O m p A /O p rF
T hese analogous proteins play an im portant role in m aintaining the structural stability of the E . c o l i and P . a e r u g i n o s a cells, respectively [Hancock et al.(1980)1.
OprF deficient P s e u d o i n o n a d s produce a near spherical shape in contrast to the rod shape of the wild type cell. Furtherm ore, the cells are m ore pro n e to leakage of a n u m b er o f p erip lasm ic proteins and are m ore susceptible to osm otic shock [Gotoh e_t al,(1989)]. E.coli cells bearing a lipoprotein or an OmpA mutation ap p ea r m o rp h o lo g ic a lly norm al, bu t w here a d o u b le m u ta tio n involving both these proteins occurs ( I p o ' / o m p A ' ) , the cells take on a spherical form. This defect in E.coli can be c o rrected by expression of the o p r F gene from P . a e r u g i n o s a [W o o d ru ff and H a n c o c k ,(1989)] indicating the degree of sim ilarity th at exists between the OmpA and OprF proteins.
The porins
The porins m ake up the third m ajor group of outer m em brane pro tein s. These proteins are no n -co v alen tly associated with the peptidoglycan, and form non-specific channels for the m ovem ent of small hydrophilic molecules across the mem brane.
A n u m b e r o f p o rin like p ro te in s have b een id e n tifie d in P . a e r u g i n o s a although they appear to have a greater exclusion lim it co m pared with the en te ro b ac te rial porins. P s e u d o m o n a s strains show a g rea ter resistan c e to several h y d ro p h o b ic and h y d ro p h ilic d ru g s than m any o f the E n te r o b a c te r ic e a e . T h is greater resistance of P s e u d o m o n a s strains has been attributed to the inability of these compounds to traverse the porins. This may reflect either the highly selective nature of these proteins, or the lower percentage of these porins that are open at any given time in th e p s e u d o m o n a d s [A ngus et a l.( 1 9 ^ 2 ) : Y o s h i m u r a an d N ik a id o ,(1 9 8 2 )].
Porin Minor outer membrane proteins
Lipopolysaccharide chains
Outer membrane
Periplasm
\ \ \ \ \ VN \ \ \ \ \ \ \ \ \
PeptidoglycanCytoplasmic membrane Free lipoprotein
Bound lipoprotein
Fig 1.8. The Gram necalive cell wall structure (not drawn to scale)
(adapted from Hammond et al. (1984))
Other Pseu d om o na s major proteins
The function of Protein G is not yet clear, but it may be involved in uptake of fluoroquinolone or iron [Chamberland,(1989); Yates e t a l,(1 9 8 9 )].
P rotein H I is o v erex p ressed w hen cells are grow n in m edia deficient in Mg2+, Ca^+, Mn^+, and Sr2+ [Bell and H ancock,(1989); Bell et_ai,(1991); Nicas and H an co ck ,(1 9 8 3 )]. T hese cells show increased resistance to antibiotics which are th o u g h t to act by chelating the divalent cations bound to the LPS. Protein H I is th o u g h t to bind lip o p o ly sa c c h a rid e s in p lac e o f the catio n s norm ally present. The protein cannot be rem oved or substituted by the antibiotics, and so renders the cell resistant to them.
iii) The lipopolysaccharide (LPS)
LPS is a com plex amphipathic m acrom olecule m ade up of three d istin c t regions jo in e d by co v alen t linkages (Fig 1.9.). LipidA forms the innerm ost region of the LPS m olecule integrating into the lipid bilayer. Joined to this is an oligosaccharide unit which can be separated into a low and a high m olecular weight fraction [Fensom and M eadow ,(1970); Koval and M eadow ,(1977)]. The low m o lecu lar w eight fraction consists of the core region which is linked directly to the lipidA. The high m olecular weight fraction is m ade up of the 0-an tig en ic saccharide units of the LPS, consisting of a number of unique repeating oligosaccharide units.
L ip id A
LipidA forms the endotoxic centre of the LPS, and appears to be indispensible for cell survival, since no mutants deficient in this fraction have yet been identified. A strong interaction with the matrix protein 0-8 in E . c o l i appears to be essential to maintain the overall lattice netw ork of the outer m em brane, which forms the basic fram ew ork from which the rem aining outer m em brane com ponents can orientate them selves.
P seu dom onaceae and the E n tero b actericeae, with antigenic cross reactivity observed between the lipidA com ponents of E . c o l i and P .a r e u g i n o s a [R ivera and M c G ro a rty ,(1 9 8 9 )]. K u lsh in and c o workers (1991) have characterised the lipidA fraction of two wild type and one rough m utant of P . a e r u g i n o s a , and shown them to share the same disaccharide ‘b a ck b o n e ’, but differ in the nature and the number of fatty acids present.
S a l m o n e l l a species and E . c o l i lipidA display a highly ordered hexagonal lattice network of their fatty acyl com ponents, which is p o stu lated to influence the hydrophobic barrier p ro p erty of the cells [Labischinski et al/1 9 8 5 ): Kato et al.(1990)1.
Core structure
T he core unit is a com plex p o ly sacch arid e structure w hich is c o m m o n to m any m em b ers o f the same sp ecies [R ow e and M eadow ,(1983); W ilkinson and G albraith,(1975)].
A 2-keto-3-deoxyoctonic acid (KDO) residue links lipidA to the core unit in both Enterobactericeae and Pseudom onaceae. The core unit of P . a e r u g i n o s a has been shown to contain a num ber of in v arian t sugars, although the m olar ratio of these com ponents and the order of linkage m ay d iffer betw een strains. S im ilar v a ria tio n s are know n to e x is t a m o n g s t the e n te ric b a c te ria [Jansson et al.(1981)1.
0 -antigenic region
T he 0 - a n tig e n ic region is m ade up o f a series o f rep eatin g o ligom er units whose length and com position is highly variable, thus forming the basis for immunological typing of cells.
T he 0 - a n tig e n ic p o ly sa cc h arid e unit can be reso lv ed into an amino-sugar rich region and a neutral-sugar rich region [Yokota e t a l,(1 9 8 7 ); Saw ada et al.(1985)1. w ith the sh o rte r n e u tra l-s u g a r region being antigenically related in P . a e r u g i n o s a of d ifferent s e r o t y p e s .
M utants defective in synthesis of the LPS are defined upon the e x ten t to which the 0 - s id e chain and core are affected. Figure
1.10.1. illu stra te s the series of LPS m u tan ts iso lated from S . t y p h i m u r i u m . These are termed rough mutants, since the cells
P o ly sacch arid e ch ain LPS
KDO
G lu co sam in e d isa c c h a iid e
Lipid A
F atty acid ch ain s
take on a grainy edged m orphology in co n trast to the sm ooth appearance shown by wild type cells. A sim ilar series of mutants have been derived from P . a e r u g i n o s a strain P A C l and P A C IR , a spontaneous m utant of P A C l but which shares its LPS structure [Chester and Meadow, (1975)] (Fig 1.10.2.).
LPS from smooth strains of P . a e r u g i n o s a have identified two distinct forms, a low m olecular w eight A band LPS, and a high m olecular weight B band LPS. The two bands differ significantly in composition. The larger B band is thought to be the main antigenic structure exposed on the cell and is serotype specific, whilst the A band of P . a e r u g in o s a w a s found to be antigenically related to a num ber of cells differing in their serotype. No amino-sugars have been detected in the A band LPS but a high rhamnose content was observed [Arsenault et al.(1991)1. A further difference lies in the lipidA com ponent of both LPS. L ipidA antibody raised in E . c o l i was shown to cro ss-react with the lipidA o f B band LPS of P . a e r u g i n o s a , but not with that from the A band. This may be a result of the low level of phosphate groups found in the A band LPS to which the antibodies bind.
1.6. NAPHTHALENE HYDROXYLATION 1 .6 .1 . N aph th alen e degradation
T h e c o m p l e t e d e g r a d a tio n o f n a p h th a le n e by m a n y soil pseudom onads, is a m ultistep process involving the form ation of the i n te r m e d ia te h y d ro c a rb o n , c a te c h o l. C a te c h o l u n d e rg o e s fu rth e r d eg rad atio n via a ring cle av a g e p ro ce ss to form the tr ic a rb o x y lic in te rm e d ia rie s used in the p ro d u c tio n of ATP. C leavage of catechol can either lead to the form ation o f c i s , c i s -m uconic acid (-m ^tfl-cleav ag e) (Fig 1.11), or 2 -h y d ro x y -m u co n ic s e m ia ld e h y d e ( o r f A o -cleav ag e). The final T C A p ro d u c ts are dependent upon which cleavage pathway is used.
N a p h t h a l e n e u n d e rg o e s h y d r o x y la tio n by b o th n a p h th a le n e dioxygenase and toluene dioxygenase via sim ilar processes. Each enzym e is able to recognise a variety of compounds related to its own n atural substrate, and c ataly se the reactio n in the same ste reo sp e cific m anner.
P.Etn
O-SIDE CHAIN
GlcNAc D.Gal I I _ ^ ^
I
\
, , .► D.Glc — D.Gal — D .G lc_L L D .H epJ-L D .H epJ- KDO— K D O ^ LIPID/*
SLPS RaLPS
I
RcLPS
ReLPS Rd2LPS
RdlLPS
Fig. 1.10.1 Core polvsacharides o f rough mutants o f S . t y p h i m u r i u m (Ra-Re) P=phosphate; P .E tn =0 -phosphorylethanolamne;
P .P .E tn = 0 -pyrophosphorylethanolam ne (adapted from Hammond et al , (1984))
Glc Glc
Rha Glc
I I
GalN — >■ Hep — >■ dOclA — dOcl A
1
1
0-ANTIGEN Alanine
1 . 6 . 2 . P la s m id e n c o d e d g e n e s in v o v e d in n a p h th a le n e d e g r a d a t i o n
Genes encoding the enzymes involved in naphthalene degradation are often found encoded upon plasmids. However, these genes may also be encoded by the chrom osom e [W eightm an et al. (1984)], or distributed between the plasm id and chrom osom e such that part of the degradative process is encoded by genes on the chrom osom e and the rem ainder by those on the plasm id [Reineke ^ ,(1980); Fennewald et al.(1979)1.
A num ber of plasmids have been isolated from pseudom onads able to degrade naphthalene, the best studied being plamid NAH7 isolated from P . p u t i d a strain PpG7 [Dunn and Gunsalus,(1973)]. Table 1.1. lists some of the plasmids found, not including those obtained by cloning or mutagenesis. Transposon Tn5 mapping of NAH7 has enabled the genes to be ordered into two distinct opérons, nah and sa l ; n a h {nah A-nah F), encodes the upper p a th w ay e n zy m es c a ta ly sin g b rea k d o w n o f n a p h th a le n e to salicylate, and sal (nah G-n ah K), the lower pathway enzymes encoding salicylate degradation via the meta c le a v a g e p ath w ay [Yen and Gunsalus,(1982); Yen et al.(1983)1.
P la s m id C om pound degraded
CAM c a m p h o r
OCT octane, decane
SAL 1 sa lic y la te
NAH n a p h t h a l e n e
TOL to lu e n e
NIC nicotin, nicotinam ide
pEG s t y r e n e
p W R l 3-chlorobenzoic acid
pAC25 3-chlorobenzoic acid
Table 1.1. Naturally occurring degradative plasm ids
1 .6 .3 . Naphthalene D ioxygenase
The first step in the degradation o f naphthalene is its hyd roxylation by the enzym e naphthalene d io x y g en a se. The enzym e c a ta ly ses the addition o f tw o o x y g en atom s from m olecular oxygen at positions C l and C2, in a stereosp ecific manner, to form (-\-)-cis ( 1 R , 2 S ) d i h y d r o x y 1 , 2 d i h y d r o -naphthalene [Jeffrey et al.n975V. Jerina et al.(197111.
The isolated enzym e is unstable upon exposure to air [Catterall and W illiam s,(1971)], but can be stabilised by inclusion of ethanol, g ly c e r o l and d ith io th r e ito l in the e x tr a c tio n b u ffe r [Laborde,(1979); Ensley et_al,(1982)].
Subsequent isolation and purification of naphthalene dioxygenase [E nsley et al. d 98211. indicated that the enzym e was in fact a multimeric protein made up of three subunits designated A, B and C. The subunits contain iron-sulphur chromophores at their active centre which coordinate the transfer o f electrons from a suitable donor to the final acceptor m olecule. Electrons are transferred se q u en tia lly from ferredoxin(nap)reductase (com ponent A ) to ferredoxin(nap) (c o m p o n e n t C ), and finally to the terminal iron- sulphur protein (component B) (Fig 1.12.).
Component A: Ferredoxintnap lR ed uctase.
F e r r e d o x in (n a p )r e d u c ta s e is the primary unit o f naphthalene d ioxygen ase, being a 2Fe-2S flavoprotein o f m olecular w eight 36,000. In the presence of NADH, the enzyme binds the pyridine nu cleotid e transferring one or two electrons to a variety o f electron acceptors including cytochrom e C [Ensley et__al,(1982); H aigler and G ibson,( 1990a)], ferricyanide and Nitrotetrazolium B lue [Haigler and G ibson,( 1990a)]. NADPH can also act as an electron source, although the resultant activity was observed to be less than half that using NADH [Haigler and Gibson,( 1990a)].
Component C: Ferredoxin£n ap ).
nah
o
OO
I
CD.H
n
CHo OOOH
CH3 GOOH
(jxm
PYRUVATE +
ACETALDEHYDE
sal
Fig 1.11. Naphthalene degradation via the m e t a -cleavage pathway Enzymes involved:
A , Naphthalene dioxygenase; B ,cfj-naphthalene dihydrodiol dehydrogen ase; C , 1,2-dihydroxynaphthalene dioxygenase; D , 2-hydroxychrom ene-2-carboxylate isomerase; E , 2-hydroxybenzalpyruvate aldolase; F , salicyl aldéhyde dehydrogenase; G , salicylate hydroxylase; H , catechol-2,3-d ioxy genase; I, hydroxymuconic semi aldehyde dehydrogenase; J , 4-oxalo-crotonate tautomerase; K , 4-oxalo4-oxalo-crotonate decarboxylase; L , 2-oxopent-4-enoate hydratase; M , 2-oxo-4-hydroxypentanoate aldolase.
ferredoxin(nap) is esen tial for activity o f naphthalene d io x y genase, and its function cannot be substituted by analogous ferred o x in s from sim ilar sy stem s [A ndrew et al, ( 1 9 7 6 ) ; Subramanian et_al,(1979); Gunsalus and L ipscom b,(1973)]. This suggests that a sp ecific interaction between ferredoxin(nap) and the terminal com ponent may be required for the transfer o f electrons. H owever, the interaction with ferredoxin(nap>reductase appears to be less stringent, and spinach(nap>reductase is able to substitute its function, but at a reduced level o f activity.
Component B: Iron-Sulpur Protein(nap)_HSP(n^p)l.
The terminal com ponent o f the naphthalene dioxygenase has a m olecular w eight o f 158,000 which can be resolved into two p o ly p ep tid e subu nits o f 5 5 ,0 0 0 and 2 0 ,0 0 0 [E n sley and G ib son ,(1 9 8 3 )]. This is su ggestive o f an a 2 p 2 quarternary structure. Both fe r r e d o x in (n a p )r e d u c ta se and f e r r e d o x in ( n a p ) activities are required for ISP(nap) to accept electrons from NADH, and no activity is observed if only one or other of these functions is p r esen t.l^ C -n a p h th a len e stu d ies sh o w ed that I S P ( n a p ) reversibly binds the substrate, most likely at the enzyme's active site.
1.6.4. T oluene dioxygen ase
electron movement
NADH-NAD
Reductase (red)
(nahA)
Reductase (ox)
Ferredoxin (red)
(nahC)
Ferredoxin (ox)
TQP
DIOL
NAP
C om p arison o f the n u c le o tid e se q u en ce s o f nap hth alen e d io x y g en a se and benzene d io x y g en a se from P. put i da s t r a in s identified regions o f D N A show ing more than 60% hom ology [Kurkela et al.(1988)1. However, the amino acid hom ology was less stringent, su ggestin g that both enzym es had em erged from a common ancestral gene, but that the gene products had diverged over time.
1 .6 .5 . Regulation o f the nah and s a l o p é r o n s
Enzym es involved in naphthalene degradation were found to be active at low lev els in the absence o f any exogen ou sly added factors [Barnsley,(1974); Yen and G unsalus,(1982); S ch ell,(1985)]. H owever, when salicylate or its analogue, 2-am inobenzoate, was added to P s e u d o m o n a s strains, N C IB 9 8 1 6 , A T C C 17483 and PpG277 harbouring the Nah plasmids, induction o f a number o f
n a h encoded enzym es produced a 20fold increase in activity [B arn sley,(1 9 7 4 )].
Salicylate is able to induce all the enzymes encoded by the n a h
and s a l opérons, with an intact n a h R gene essen tial for this coordinated induction to occur [Sch ell,(1983); Schell and Wender, (1986); Grund and Gunsalus,(1983); Yen and G unsalus,(1982)].
n a h R ' cells produce a pleiotropic effect resulting in a n a h ' / s a l '
phenotype, which can be corrected by in t r a n s com p lem en tation using a second, intact n a h R a llele [S ch ell,(1 9 8 5 ); Grund and G un salus,(1 9 8 5 )].