1
SYNOPSIS
Even after illuminating most of the deep mysteries of brain, there is still a lot to know about this miraculous organ. The brain is a complex of pathways performing several different functions at a time. The molecular mechanism of these pathways needs insight to understand the complexity of the organ. The Olfactory Transduction is one such pathway which neuroscientists have wondered for its specificity to organisms. The pre-eminent objective of this project was to understand the molecular mechanism of this pathway by analysing different molecules involved in the pathway, their dominant functions and to study their evolutionary relationship among different vertebrates. The central idea involved in this comparative analysis was to correlate the molecular evolution with phylogenetic evolution. For the comparative analysis study, a comparison of 15 molecules involved in OT pathway namely; CALM (calmodulin), PKA(protein kinase A), CAMK2 (Calcium/calmodulin-dependent protein kinase), GNAL (guanine nucleotide binding proein), CNG (Cyclic nucleotide gated channel) A3, A4, B1, CLCA(calcium activated chloride channel) 1,2,3,4, PRKG(Protein kinase cGMP-dependent), ADCY3 (Adenylate cyclase 3), PDC( Phosducin), GUCA1(Guanylate cyclate activator 1) was done among eight different organisms namely; Human, Chimpanzee, Cow, Dog, Mouse, Chicken, Xenopus laevis and Zebrafish. Comparitive analysis showed that there is no significant relationship between molecular evolution and phylogenetic evolution. The OT pathway shows molecular evolution where number of components within the pathway varies among organisms which is specific to their ability to smell.
Apart from analysing OT pathway, another objective was to collect the brain molecules involved in OT pathway for different organisms and to update the available database, i.e. Molecular Database of Brain (MDB) freely available at http://bicmku.in/mdb, with these molecules. 390 molecules were collected for OT pathway. Currently, this database has 1251 brain molecules related to LTP, PT and OT pathway having the details of their gene, proteins and ligands.
In this short period of time, an additional objective was to study chimeric molecule of Outer membrane protein F (OmpF) and endolysin gene as a way for enzyme presentation by a molecule fixed to membrane. This idea would further help in bioengineering of molecular presentation systems. This is included in APPENDIX A.
2
In APPENDIX B, is also reported the crystallography of lysozyme crystallization which was done as a part of learning process as it is a best model to study crystallization and is also comfortable to work with. Different concentrations of lysozyme were tried to study its effect on morphology of crystals. Besides, this trehalose, a cryoprotectant was also used to study its interaction with lysozyme to understand the whole mechanism behind this property of trehalose. The inherent property of trehalose acting as a heat-stress compound was exploited to collect data to make the process more cost effective. Co-crystals of lysozyme with trehalose were obtained at varying concentrations of trehalose from 5%, 10%, 15%, 20%, 25%, 30% and 35%. Though after data processing the ligand, trehalose was not found to be bound with lysozyme protein.
3
4
1.1 Olfaction – Sense of smell
Olfaction or Olfactory perception is one of the most exquisite senses to communicate with the environment. It is the phenomenon of sensing smell, allowing detection of food, predators, mates, provides sensual pleasure of odour of flowers and perfumes and also gives warning alarms like that of chemical dangers and spoiled food. The sensory system used for olfaction is Olfactory System, which is one of the most pre-eminent senses, having remarkable capacity of detecting odorant molecules. These odorant molecules are volatile chemical molecules which may be air-borne molecules, detected by vertebrates, including humans, or water-soluble molecules detected by aquatic animals. Olfaction has also been reported in invertebrates like various insects including Drosophilla melanogaster [1], moths and honeybees and also in worms like C. elegans [2]. Reports are also there for olfaction in plants as their tendrils are sensitive to air-borne volatile compounds [3, 4]. In comparison to dogs and rhodents, though humans have poor olfactory ability, still they can specifically detect a wide range of odorant molecules.
Olfaction is a type of chemosensory process in which volatile odorant molecule binds to specific receptors present on olfactory epithelium in nose leading to a series of chemical reactions and finally transducing the signal to the brain. The brain not only analyses the signal and categorizes the odorant molecule but also memorizes the variety of odours each time it comes across. In 1991, Richard Axel and Linda Buck were honoured by the Nobel Prize in the field of Physiology and Medicine, for their aesthetic research on the interpretation of odorant patterns by brain where the discovered that brain can detect and memorize different odorant molecules because of the special receptors, called olfactory receptors present in the nasal epithelium [5].
Olfaction is also associated with some disorders like Anosmia- inability to smell, Hyperosmia- having acute sense of smell, Hyposmia-decreased sense of smell and Dysosmia-smelling things differently.
1.1.1 Olfactory Receptors
The ability of sensing millions of odorant molecules present in a different way is due to the presence of olfactory receptors. Olfactory receptors are largest multigene family in vertebrates and are chiefly located in the form of clusters in the posterior part of nasal cavity forming an olfactory epithelium, which is the primary sensing structure [6].
5
In insects, it is located on their third segment of antennae and other chemosensory organs [7]. There has also been a report suggesting that olfactory receptors are present on sperms, which detects a chemical called Bourgeonal, released by egg cell by chemotactic movement [8, 9, 10].
Basically, these olfactory receptors (ORs) belong to heptahelical G-protein coupled receptors (GPCRs) superfamily that is invariably seven transmembrane domain (7TM) proteins.
Figure 1.1 Structure of Olfactory Receptors showing seven transmembrane domain proteins.
1.1.2 Types of Olfactory Receptors
ORs can be classified into two types of classes based on their amino acid sequences and phylogenetic analysis:
Class I ORs referred as fish-like ORs specialized in recognizing water-soluble odorants, and,
Class II ORs referred as mammalian-like ORs recognizing volatile odorants.
The olfactory repertoire genes has been estimated to be 650 to 900 in humans [11, 12], 1200 to 1500 in mice [13, 14, 15], 1700 to 2000 in rats [16] and 1300 in dogs [17, 18,].
6
This may attribute to the reason why dogs and mice have extraordinary smelling power which humans don’t have [19].
1.1.3 Nomenclature of Olfactory Receptors
To avoid confusion in naming of these large numbers of olfactory genes, a nomenclature system has also been constructed for olfactory gene receptor family [20]. The name of the individual family are given in the format ORnXmY, where,
OR is the root name indicating Olfactory Receptor superfamily
n is an integer representing a family whose members have greater than 40% sequence identity
X is a single letter denoting a subfamily,
m is an integer representing an individual family member, and, Y is an integer denoting that it is a pseudogene.
1.1.4 Olfactory Transduction (OT)
Although every day we come across different types of odour which may be familiar or unfamiliar to us, but still we are able to detect it. So, the question arises that how the brain can identify and categorize every odorant molecule. The answer to this question lies in the cellular composition of the olfactory epithelium and specificity of the olfactory receptors to a particular class of odorant molecule.
Odorants are small volatile organic molecules of molecular weight less than 400 Da varying in size, shape, functional groups and charge [21]. These can be alcohols, aldehydes, ketones, esters or amines or can be aromatic, alicyclic, polycyclic or heterocyclic ring structures or can be substituted compounds of these types. Differences in the structure of an odorant can lead to modification in its quality [22].
7
1.1.4 (a)
Anatomical Organization of Olfactory Epithelium
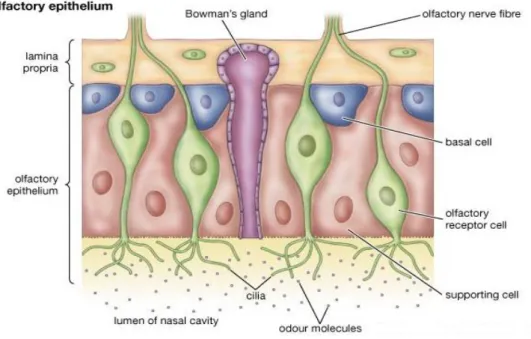
The olfactory system is well adapted structurally to perform its specific function. The olfactory epithelium present in the nasal cavity which is the primary sensory structure is basically made up of three types of cells:
Olfactory receptor neurons (ORNs) Supporting sustentacular cells Basal cells.
ORN is a bipolar neuron whose axons extends to the olfactory bulb present in the forebrain while its apical dendrites extend to the surface of neural epithelia forming cilia and detects odour [23, 24, 25, 26]
Figure 1.2 Structure of Olfactory epithelium showing olfactory receptor neurons, supporting cell and basal cells.
1.1.4 (b) Mechanism Of Olfactory Transduction
Though the mechanism is well known in vertebrates, it is still not well understood in invertebrates.
The main mechanism behind olfaction involves cascade of events of signal transduction called Olfactory Transduction, where the chemical signal is transformed into electrical signals and sent to the brain which perceives it as smells [24, 25]. The whole event of signal
8
transduction involves the binding of odorant molecule to specific ORs leading to the activation of G-protein signal transduction [27]. Odorous ligands bind to specific olfactory receptors expressed on ORNs which subsequently gets coupled to an olfactory specific G-protein (Golf) and activates adenylyl cyclase type III. This reaction transduces chemical information into electrical involving amplification of signal. Activation of adenylyl cyclase increases intracellular cAMP (cyclic AMP) levels leading to the opening of cyclic nucleotide gated ion channel (CNG). This allows cations, mainly, calcium (Ca2+) and sodium (Na+) ion to flow down their electrochemical gradients into the cell leading to the depolarization of the cell membrane ORNs. The Ca2+ entering through CNG channel are also able to activated calcium activated chloride ion channel (CLCA) which is permeable to negatively charged chloride ion (Cl-), thus allowing it to flow out of the cell, leading to increase in depolarization. Furthermore, increase in Ca2+ concentration also adapts a negative feedback loop pathway which leads to two important steps:
Inhibition of adenylyl cyclase activation by calmodulin-dependent protein kinase (CAMK)
Downregulation of the affinity of CNG channel to cAMP.
Figure1.3 Flowchart showing the mechanism of Olfactory Transduction.
9
Figure 1.4 Diagrammatic representation of the mechanism of olfactory transduction pathway showing molecules involved in the pathway.
Biochemical studies have also shown that long stimulation to odorant molecules can lead to the elevation of cGMP levels [28, 29, 30]. Several Ca2+ ion independent mechanism have been proposed to mediate odour adaptation, including odour dependent phosphorylation of protein kinase A (PKA) and G-protein couples receptor kinase 3 (GSK3) [31, 32].
1.1.5 Molecules involved in OT Pathway
Broadly, there are 16 molecules present in olfactory transduction pathway.
1. Calmodulin (CALM)
CALcium MODULated proteIN is abbreviated as calmodulin which is abundantly expressed in cytoplasm of all eukaryotic cells and functions as a calcium-binding messenger
protein. It mediates the control of a large number of enzymes, ion channels and other proteins by Ca2+ and can stimulate number of protein kinase enzymes and phosphatases. Calmodulin also helps in the activation of adenylate cyclase in a calcium dependent manner and independent of direct stimulation of odorant molecules [33, 34].
10
2. Olfactory receptors (ORs)
These are G-protein coupled receptors (GPCRs) having seven transmembrane domains and act as primary sensory structure to detect odorant molecule. Binding of an odorant molecule then mediates olfactory transduction [35].
3. Protein kinase A (PKA)
Also known as cAMP-dependent protein kinase performs several functions in phosphorylating large number of substrates in the cytoplasm and nucleus [36]. It is a family consisting of four components, namely, PRKACA, PRKACB having catalytic alpha, beta subunit respectively and X-linked PRKX.
4. Calcium/calmodulin-dependent protein kinase (CAMK2)
Consists of four different subunits, CAMK2A, CAMK2B, CAMK2D, CAMK2G having catalytic subunits alpha, beta, delta and gamma. These molecules perform several functions like extracellular signal-regulated kinase and regulation of Ca2+ homeostasis [37].
5. Guanine nucleotide binding protein alpha subunit (GNAL)
These are basically adenylate cyclase stimulating G-alpha protein which are involved in modulating transmembrane signalling systems and mediates signal transduction within olfactory epithelium and basal ganglia [38].
6. Cyclic nucleotide gated channel (CNG)
It also consists of three subunits, namely, CNGA3, CNGA4 and CNGB1 having alpha3, alpha4 and beta1 catalytic subunits. It has ligand-gated ion channel activity and plays a central role in the transduction of odorant molecules [39, 40].
7. Calcium activated chloride channel (CLCA)
Its family consists of four different molecules: CLCA1, CLCA2, CLCA3, and CLCA4. It mainly has chloride channel activity and hydrogen ion transporting ATPase activity [41].
11
8. Protein Kinase cGMP dependent (PRKG)
It consists of two isoforms: PRKG1 and PRKG2 and is strongly expressed in smooth muscles, platelets and hippocampal cells. It mainly phosphorylates many cellular proteins and has ATP binding, cGMP binding activity.
9. Adenylate cyclase 3 (ADCY3)
It catalyzes the formation of the secondary messenger, cAMP. 10.Phosducin (PDC)
It is a phosphoprotein which mainly participates in regulation of visual phototransduction. It also shows phospholipase inhibitory activity. An odorant-induced rise of cAMP levels would activate protein kinase A leading to phosphorylation of phosducin which triggers the membrane targeting of GRK3 and phosphorylation of the receptor protein [42].
11.Guanylate cyclase activator 1 (GUCA)
It has two isoforms GUCA1A and GUCA1B and plays an important role in regulation of guanylyl cyclase by stimulating guanylyl cyclase1 when Ca2+ concentration is low and inhibits when free Ca2+ concentration is elevated [43].
12
Figure 1.5 Diagrammatic representation of all the molecules involved in OT pathway.
1.2 MOLECULAR DATABASE OF BRAIN
Of all the objects in the universe, human brain is the most complex. Despite of the recent advances in the science of the brain and mind, the mystery of the brain is still unrevealed. For better understanding of the brain, many databases have been developed for brain which has made the research easier. Nowadays, several databases of brain are available like Brain web: simulated Brain Database http://brainweb.bic.mni.mcgill.ca/brainweb/, The BRAINnet database http://www.brainnet.net/about/brain-resource-international-database/ Brain Resource Database http://www.brainresource.com/about-us/brain-resource-database, PMOD Normal Brain Database http://www.pmod.com/technologies/products/braindatabase_tool/braindatabase_tool.php, Sense lab http://senselab.med.yale.edu/ which are freely accessible for the researches and help them in understanding of various brain disorders and genetics and functional analysis of brain. None of these databases provides molecular information of the brain. The Molecular Database of Brain (MDB) http://bicmku.in/mdb, is one such novel database which deals with
13
the molecular aspects of brain. MDB is a unique database that has molecules related to different pathways of brain having details of all the genes, proteins and ligand involved in that pathway in a range of number of organisms like Humans, Chimpanzee, Cow, Dog, Mouse, Chick etc. MDB already has 317 molecules involved in Long Term Potentiation Pathway (LTP) [44] and Phototransduction Pathway (PT) [45].
The attempt was to update MDB with molecules related to Olfactory Transduction Pathway (OT). At present there are few databases for Olfactory Receptors (OR) namely, ORDBhttp://senselab.med.yale.edu/OrDB/,OdorDBhttp://senselab.med.yale.edu/OdorDB/, OdorMapDBhttp://senselab.med.yale.edu/OdorMapDB/,HORDEhttp://genome.weizmann.ac.il/hor de/, but all of them has information related to only ORs. MDB consists of all the molecules that are involved in the pathway.
Kyoto Encyclopedia of Genes and Genomes (KEGG) database (http://www.genome.jp/kegg/) was used to retrieve all the information related to Olfactory Transduction Pathway. About 390 molecules related to olfactory transduction pathway were collected. An attempt was made to understand olfactory perception process in detail by phylogenetic analysis of molecules involved in the pathway among eight organisms.
Presently, MDB has 1251 brain molecules related to different pathways. MDB is also being updated with the structural information of brain molecules which contains three categories: structure, model and Structural Genomics Initiative (SGI). This will again proof to be a great help for the neuroscientists for the functional analysis of brain molecules [46].
14
15
2.1 Analysis of Olfactory Transduction (OT) Pathway
2.1.1 Retrieval of sequences of molecules of OT from different organisms
For the analysis of OT pathway, 15 molecules were chosen- 1. CALM (Calmodulin)
2. PKA (Protein Kinase A)
3. CAMK2 (Calcium/calmodulin-dependent protein kinase) 4. GNAL (Guanine nucleotide binding protein alpha type) 5. CNGA3 (Cyclic nucleotide gated channel alpha 3) 6. CNGA4 (Cyclic nucleotide gated channel alpha 4) 7. CNGB1 (Cyclic nucleotide gated channel beta 1) 8. CLCA1 (Calcium-activated chloride channel alpha 1) 9. CLCA2 (Calcium-activated chloride channel alpha 2) 10. CLCA3 (Calcium-activated chloride channel alpha 3) 11. CLCA4 (Calcium-activated chloride channel alpha 4) 12. PRKG (Protein kinase cGMP dependent)
13. ADCY3 (Adenylase cyclase 3) 14. PDC (Phosducin)
15.GUCA1 (Guanylate cyclase activator 1)
These molecules were phlylogenetically compared among eight different classes of vertebrates.
I. Class: Mammalia Order: Primates – Humans (Homo sapiens) II. Class: Mammalia Order: Primates – Chimpanzee (Pan troglodytes) III. Class: Mammalia Order: Artiodacyla – Cow (Bos taurus)
IV. Class: Mammalia Order: Carnivora – Dog (Canis familiaris) V. Class: Mammalia Order: Rhodentia – Mouse (Mus musculus) VI. Class: Aves Order: Galliformes – Chicken (Gallus gallus)
VII. Class: Amphibia Order: Anura – African clawed frog (Xenopus laevis) VIII. Class: Actinopterygii Order: Cypriniformes – Zebra fish (Danio rerio)
16
2.1.2 Phylogenetic Analysis of Pathway
Multiple sequence alignment (MSA) of eight organisms was done using ClustalW (http://www.ebi.ac.uk/2can/tutorials/nucleotide/clustalw.html). Phylogenetic trees were constructed using neighbour joining method as implemented in MEGA 5.1.4 (http://www.megasoftware.net/) for 15 molecules in the pathway. The Neighbour joining method gave tree topologies with minimum branch length. It resulted in producing un-rooted, bifurcating trees [31]. The clustering in the constructed tree were then validated using the Bootstrap (a statistical method which provides the repeatability of clustering observed in randomized trials), where the number of trials was set to 100. Nodes with bootstrap value greater than 50% were used for analysis.
2.2 Molecular Database of Brain (MDB)
2.2.1 Data collection and Updating the Database
Different databases were accessed like KEGG Pathway Database
(http://www.genome.jp/kegg/pathway/), NCBI-PubMed (www.ncbi.nlm.nih.gov/pubmed), Uniprot/Swissprot (web.expasy.org/docs/swiss-prot_guideline.html), Protein Data Bank (PDB- www.rcsb.org ) for collection of molecules and various details like gene name, gene symbol, chromosome location, functions etc.
The relational database management system MySQL is used for updating the database. HyperText Markup Language (HTML) and Cascading Styling Sheets (CSS) were used for styling and designing. PHP:Hypertext Preprocessor was used as the interface. This helped in providing a user friendly resource to access information on brain related molecules.
17
2.2.2 Sequence Alignment Studies
Jalview (www.jalview.org) was used for alignment studies. The alignment score using nucleotide sequences was calculated according to the following formula:
Alignment Score = Percentage Identity/ Alignment length.
Dividing the percentage identity by alignment length ensures that, small alignments with high identity do not bias the result.
Percentage identity was also calculated using jalview which was used for making scatter plot of average % identity of molecules involved in the pathway among human, chimpanzee, dog, mouse and fish.
Standard deviation (SD) was calculated for average percentage identity of the molecules involved in the pathway among these organisms. Then the range of standard deviation was calculated by adding and subtracting the standard deviation value to obtain significant range of variation among the molecules.
Maximum Deviation = Mean value + SD Minimum Deviation = Mean value – SD
Scatter plot was made to plot the graph between molecules involved in the pathway against Average % identity. The plot signifies the conservation or variation of a molecule among different organisms.
18
19
3.1Analysis of the olfactory transduction (OT) pathway
The OT pathway was understood based on: Molecular analysis Phylogenetic analysis.
3.1.1 Molecular Analysis of OT Pathway
The OT pathway is comprised of different molecules which work as a cascade to help in olfactory perception. These molecules consist of different components which vary in different organisms of different families or within the same family. The OT pathway can actually be
referred as DUAL SECONDARY MESSENGER PATHWAY, as it involves two different
secondary messengers, i.e. cAMP and cGMP for signal transduction, which are triggered by different concentration of odorant molecules. The first secondary messenger pathway involves cAMP while the second secondary messenger pathway involves cGMP which is triggered at high concentration of odorant molecules after its long exposure.
The overall OT pathway is shown as in flowchart diagram. Figure3.1.
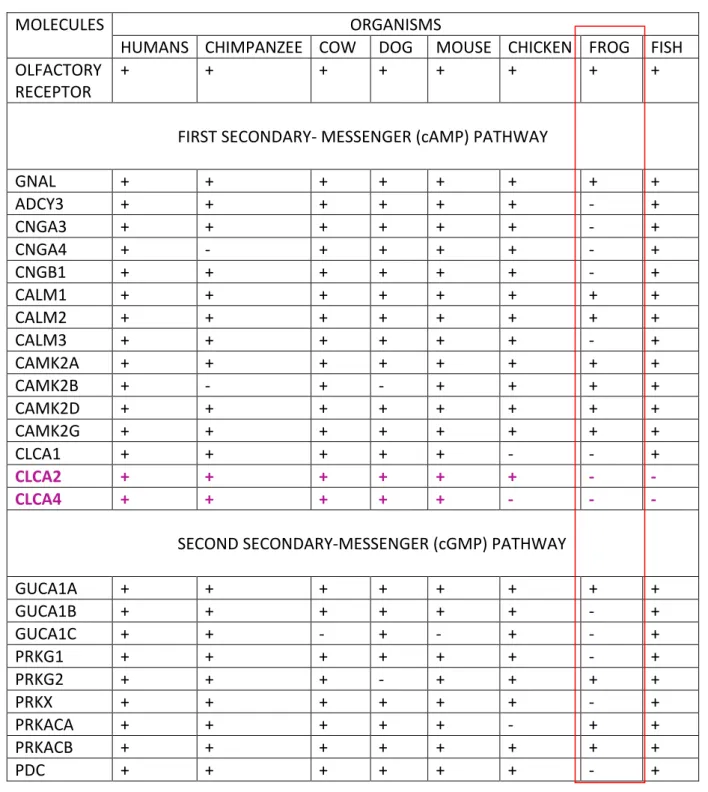
The molecules and their different components involved in OT pathway were analysed among eight organisms, namely, Human, Chimpanzee, Cow, Dog, Mouse, Chicken, Frog and Fish. The molecules were analysed along the pathway for both First secondary-messenger pathway and Second secondary-messenger pathway.
After the analysis of OT pathway in above selected organisms, it was found that no orthologs are present in chimpanzee for the molecule CNGA4 and CAMK2B. No ortholog of CAMK2B was found in dogs also. CLCA2 and CLCA4 are not present in frog and fishes. Most of the molecules involved in OT pathway in other organisms are as per the KEGG Database, are not present for frogs.
20 FLOWCHART DIAGRAM
21
Molecular analysis of the components involved in the pathway is shown as in Table 3.1.
MOLECULES ORGANISMS
HUMANS CHIMPANZEE COW DOG MOUSE CHICKEN FROG FISH
OLFACTORY RECEPTOR
+ + + + + + + +
FIRST SECONDARY- MESSENGER (cAMP) PATHWAY
GNAL + + + + + + + + ADCY3 + + + + + + - + CNGA3 + + + + + + - + CNGA4 + - + + + + - + CNGB1 + + + + + + - + CALM1 + + + + + + + + CALM2 + + + + + + + + CALM3 + + + + + + - + CAMK2A + + + + + + + + CAMK2B + - + - + + + + CAMK2D + + + + + + + + CAMK2G + + + + + + + + CLCA1 + + + + + - - + CLCA2 + + + + + + - - CLCA4 + + + + + - - -
SECOND SECONDARY-MESSENGER (cGMP) PATHWAY
GUCA1A + + + + + + + + GUCA1B + + + + + + - + GUCA1C + + - + - + - + PRKG1 + + + + + + - + PRKG2 + + + - + + + + PRKX + + + + + + - + PRKACA + + + + + - + + PRKACB + + + + + + + + PDC + + + + + + - +
Table 3.1 Dual secondary messenger pathway analysis of the molecules involved along the pathway. As per the pathway databases, the molecules involved in the pathway are denoted as ‘+’ while the molecules absent are denoted as ‘-’sign. The molecules highlighted in red and pink are discussed further.
22
3.1.2 Sequence Alignment studies
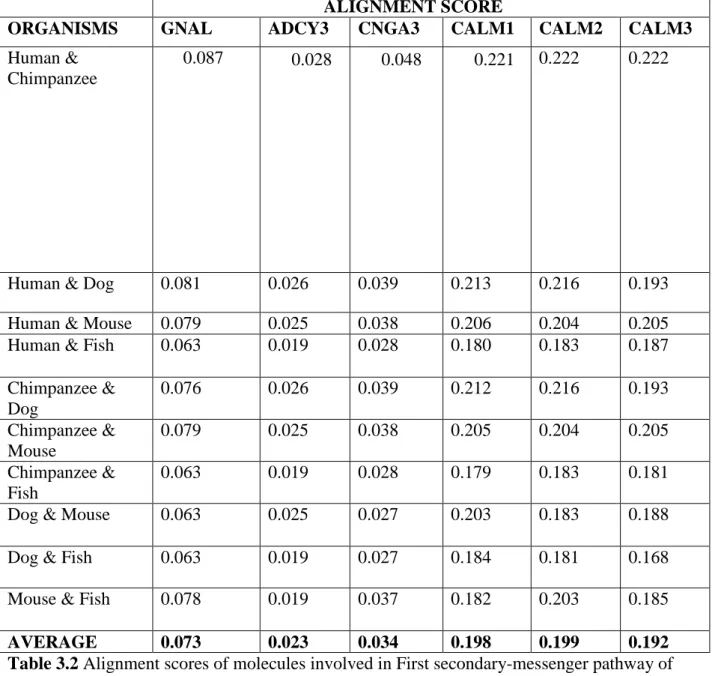
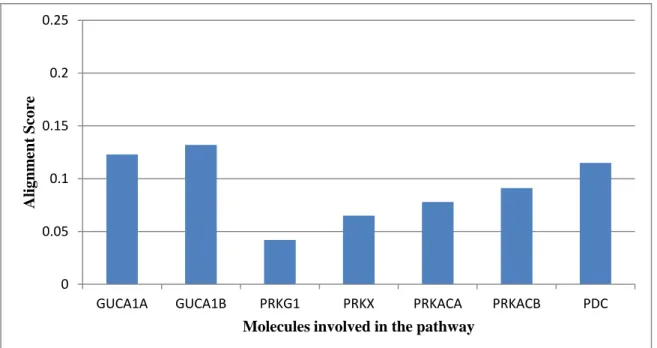
The alignment scores were calculated for looking the conservation or variation of a particular molecule involved along the pathway which is shown here in the Table 3.2, 3.3 and 3.4. Only those molecules were considered for calculating the alignment scores for which sequences are present. Similarly only those organisms were considered for sequence alignment studies, which has sequences of all the molecules considered above. Then the average scores were taken for these molecules involved in the pathway, which were then denoted graphically as a plot of average alignment score for each molecule along the pathway which is shown in Figure 3.2 and 3.3.
ALIGNMENT SCORE
ORGANISMS GNAL ADCY3 CNGA3 CALM1 CALM2 CALM3
Human & Chimpanzee
0.087 0.028 0.048 0.221 0.222 0.222
Human & Dog 0.081 0.026 0.039 0.213 0.216 0.193
Human & Mouse 0.079 0.025 0.038 0.206 0.204 0.205
Human & Fish 0.063 0.019 0.028 0.180 0.183 0.187
Chimpanzee & Dog 0.076 0.026 0.039 0.212 0.216 0.193 Chimpanzee & Mouse 0.079 0.025 0.038 0.205 0.204 0.205 Chimpanzee & Fish 0.063 0.019 0.028 0.179 0.183 0.181
Dog & Mouse 0.063 0.025 0.027 0.203 0.183 0.188
Dog & Fish 0.063 0.019 0.027 0.184 0.181 0.168
Mouse & Fish 0.078 0.019 0.037 0.182 0.203 0.185
AVERAGE 0.073 0.023 0.034 0.198 0.199 0.192
Table 3.2 Alignment scores of molecules involved in First secondary-messenger pathway of Olfactory Transduction
23
ALIGNMENT SCORE
ORGANISMS CAMK2A CAMK2D CAMK2G CLCA1
Human & Chimpanzee
0.064 0.066 0.058 0.036
Human & Dog 0.061 0.063 0.049 0.030
Human & Mouse 0.114 0.060 0.055 0.020
Human & Fish 0.053 0.053 0.040 0.017
Chimpanzee & Dog 0.059 0.065 0.066 0.030
Chimpanzee & Mouse
0.111 0.060 0.051 0.020
Chimpanzee & Fish 0.051 0.051 0.053 0.017
Dog & Mouse 0.095 0.051 0.039 0.020
Dog & Fish 0.055 0.053 0.044 0.017
Mouse & Fish 0.127 0.60 0.051 0.017
AVERAGE 0.079 0.058 0.050 0.022
Table 3.3 Alignment scores of molecules involved in First secondary-messenger pathway of Olfactory Transduction.
Figure 3.2 Sequence conservation and variation in molecules along the First secondary-messenger pathway of olfactory transduction.
0 0.05 0.1 0.15 0.2 0.25
GNAL ADCY3 CNGA3 CALM1 CALM2 CALM3 CAMK2A CAMK2D CAMK2G CLCA1
A li gnm ent sc ore
24
ALIGNMENT SCORE
ORGANISMS GUCA1A GUCA1B PRKG1 PRKX PRKACA PRKACB PDC
Human & Chimpanzee
0.158 0.165 0.049 0.088 0.084 0.126 0.169
Human & Dog 0.143 0.143 0.045 0.067 0.087 0.082 0.123
Human & Mouse
0.139 0.141 0.046 0.066 0.086 0.084 0.115
Human & Fish 0.113 0.113 0.037 0.061 0.074 0.068 0.080
Chimpanzee & Dog 0.139 0.143 0.045 0.067 0.082 0.120 0.144 Chimpanzee & Mouse 0.135 0.140 0.046 0.066 0.073 0.109 0.153 Chimpanzee & Fish 0.113 0.112 0.037 0.059 0.070 0.098 0.094
Dog & Mouse 0.050 0.112 0.037 0.065 0.075 0.075 0.083
Dog & Fish 0.112 0.112 0.037 0.055 0.074 0.068 0.082
Mouse & Fish 0.137 0.145 0.045 0.059 0.084 0.086 0.116
AVERAGE 0.123 0.132 0.042 0.065 0.078 0.091 0.115
Table 3.4 Alignment scores of molecules involved in Second secondary-messenger pathway of Olfactory Transduction.
Figure 3.3 Sequence showing conservation and variation in molecules along the Second secondary-messenger pathway of olfactory transduction.
0 0.05 0.1 0.15 0.2 0.25
GUCA1A GUCA1B PRKG1 PRKX PRKACA PRKACB PDC
A
li
gnm
ent
Score
25
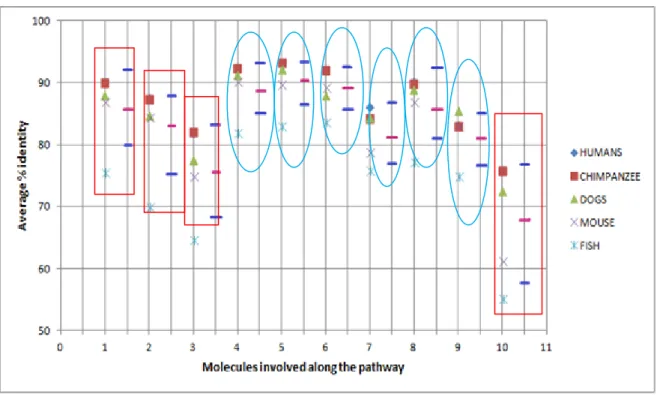
The alignment score and the graphical plot denoted that the molecules having higher alignment score are more conserved and molecules having less alignment score shows variation.
In the first secondary-messenger pathway, sequences of molecules like CALM 1, 2, 3 were found to be conserved while others shows some variation. Similarly, in second secondary-messenger pathway molecules like GUCA1A, GUCA1B were showing conservation while others were showing some variation.
The significance of the variation is measured in terms of standard deviation and was depicted in the scatter plots.
Two scatter plots were made for both secondary messenger pathways. It shows the molecules that are conserved or varying along the pathway among different organisms. Scatter plots are shown in Figure 3.4 and 3.5.
It was found that in first secondary messenger pathway, sequences of CALM1, 2, 3 and CAMK2A, 2D, 2G were conserved while GNAL, ADCY3, CNGA3 and CLCA1 were varying among organisms.
Similarly, in second secondary messenger pathway, it was observed that the molecules like GUCA1A, GUCA1B, PRKACA, PRKACB and PRKG1 were found to be conserved while PRKX and PDC were showing significant variation among organisms.
26
Figure 3.4 Plot showing the conservation and variation of molecules along the First Secondary messenger (cAMP) pathway. The numbering on X-axis indicates the molecules involved along the pathway (1-GNAL, 2-ADCY3, 3-CNGA3, 4-CALM1, 5-CALM2, 6-CALM3, 7-CAMK2A, 8-CAMK2D, 9-CAMK2G, and 10-CLCA1) and Y-axis denotes the average percentage identity. The pink line indicates the Mean value and the blue line indicate the range of standard deviation (SD), i.e. Maximum deviation (Mean + SD) and Minimum deviation (Mean – SD). The blue circle indicates the molecules that are conserved along the pathway among organisms and red boxes indicate molecules that show variation among the organisms.
27
Figure3.5 Plot showing the conservation and variation of molecules along the Second Secondary messenger (cGMP) pathway. The numbering on X-axis indicates the molecules involved along the pathway (1-GUCA1A, 2-GUCA1B, 3-PRKG1, 4-PRKX, 5-PRKACA, 6-PRKACB and 7-PDC) and Y-axis denotes the average percentage identity. The pink line indicates the Mean value and the blue line indicate the range of standard deviation (SD), i.e. Maximum deviation (Mean + SD) and Minimum deviation (Mean – SD). The blue circle indicates the molecules that are conserved along the pathway among organisms and red boxes indicate molecules that show variation among the organisms.
28
3.1.3 Phylogenetic Analysis of the Olfactory Transduction Pathway
The relationship of different molecules involved in the pathway among different class of organism was studied by constructing a Phylogenetic tree. From the phylogenetic tree, those branching pattern was considered significant which had the bootstrap value greater than 50%. Phylogenetic trees of molecules involved along the pathway are shown in Figure 3.7. Also clustering of molecules was done based on the phylogenetic tree which is shown in Figure 3.8. 1. GNAL 2. ADCY3 GNAL ADCY3
Figure 3.6 Standard taxonomic Tree
Mouse Dog
29 3. CNGA3 4. CNGA4 5. CNBG1 CNGA4 CNBG1 CNGA3
30 6. CALM1 7. CALM2 8. CALM3 CALM1 CALM2 CALM3
31 9. CAMK2A 10.CAMK2B 11.CAMK2D 12.CAMK2G CAMK2A CAMK2B CAMK2D CAMK2G
32 13.CLCA1 14.CLCA3 15.CLCA4 16.PDC CLCA1 CLCA3 CLCA4 PDC
33 17. GUCA1A 18.GUCA1B 19.PRKG1 20.PRKG2 GUCA1A GUCA1B PRKG1 PRKG2
34
21.PRKX
22.PRKACA
23.PRKACB
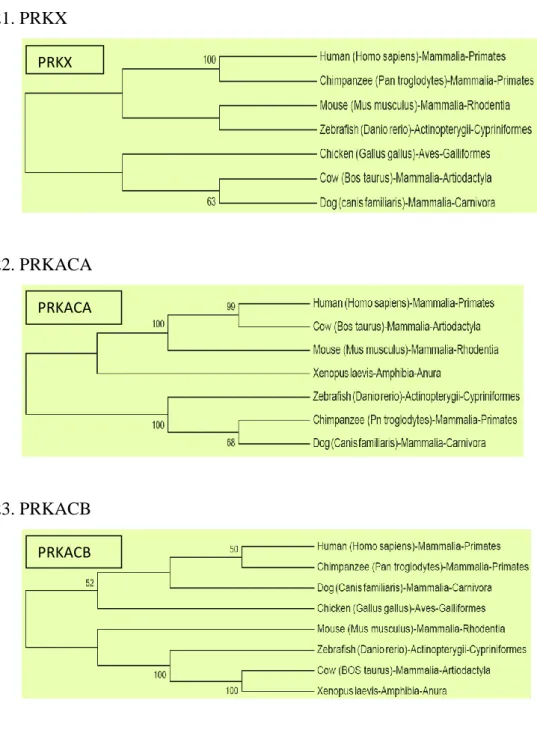
Figure 3.7 Evolutionary Trees generated using NJ method. Bootstrap values greater than 50% only given. (Mammals: Human, Chimpanzee, Dog, Cow, Mouse, Aves: Chicken, Amphibia:
Xenopus laevis, Fishes: Zebrafish). Components of same molecule are shown in same colour.
PRKX
PRKACA
35
Molecules Organisms
Human Chimp Mouse Dog Cow Chick Frog Fish
GNAL ADCY3 CALM1 CALM2 CALM3 CAMK2A CAMK2B CAMK2D CAMK2G
36
Molecules Organisms
Human Chimp Mouse Dog Cow Chick Frog Fish
CNGA3 CNGA4 CNBG1 CLCA1 CLCA2 CLCA4 GUCA1A GUCA1B PRKG1
37
Molecules Organisms
Human Chimp Mouse Dog Cow Chick Frog Fish
PRKG2
PRKX
PRKACA
PRKACB
PDC
Figure3.8 Based on the phylogenetic trees, clustering of the organisms has been made for those having bootstrap value greater than 50%. The figure compares the clustered obtained with the standard taxonomic relationship.
Thus from the Figure 3.8 it is shown that there is no reasonable comparison between the clustering of molecules based on their sequences and the standard taxonomic clustering.
3.2. Molecular Database of Brain (MDB)
Molecular database of brain gives information on both functional and structural part of the selected molecules. Gene, protein and ligand details of a particular pathway of brain can be easily accessed by simply giving the molecule name in the search bar. Figures 3.9, 3.10, 3.11 and 3.12 present the home page, view mode and search mode screenshots of MDB.
38
Figure 3.9 Home page of MDB
39
Figure 3.11 Search mode of MDB
40
3.3 Analysis of the Olfactory Transduction pathway
Molecular analysis of the OT pathway shows that though same molecules are involved in the pathway among different vertebrates, the number of components of these molecules vary from organism to organism which may contribute towards their specific olfaction power. The absence of many molecules in amphibians suggests that the function of these molecules in olfactory transduction might have been lost though these molecules may be present in frogs performing different functions. Molecular analysis of the components also shows that amphibians only have second secondary messenger pathway and the first secondary messenger pathway is not developed in them. The components of the molecules like CLCA2 and CLCA4 which are only present in terrestrial vertebrates signifies that these are terrestrial modifications and are not necessary for olfaction in aquatic life.
Sequence alignment studies shows that the initial molecules and final molecules in first secondary- messenger pathway shows variation among organisms while the molecules in the middle of the pathway are conserved. Though this pattern is not observed in second secondary-messenger pathway. Only molecules like PRKX and PDC showed significant variation among organisms. The molecules showing variation can be attributed as specific molecules evolved for olfactory transduction.
Comparison of phylogenetic tree of different molecules with standard taxonomic tree by clustering them suggested that evolution of the components of the olfactory transduction pathway does not follow the whole organism phylogenetic relationship. It is as if the molecules have evolved independently as per the need of time and function. The pathway has not evolved as one unit. The modification in the molecular components of the OT pathway in vertebrates has no significant match with their taxonomic relationship.
41
42
The olfactory transduction pathway was analysed on both molecular basis and phylogenetically among different classes of vertebrates and it was observed that there is no significant relationship between molecular evolution and phylogenetic evolution. The OT pathway showed molecular evolution where number of components within the pathway varied among organisms probably conferring specificity to their ability to smell.
Molecular Database of Brain (MDB) available at the URL http://bicmku.in/mdb, has been updated with 390 molecules involved in olfactory transduction pathway. Presently, this database has 1251 brain molecules related to three pathways namely, Long Term Potentiation (LTP), Phototransduction (PT) and Olfactory Transduction (OT).
43
REFERENCES
1. Benjamin Kaupp U. (2010). Olfactory signalling in vertebrates and insects: differences and commonalities. Nature Reviews Neuroscience 11:188-200.
2. Cornelia I. Bargmann, Hartweig E., Hortvit H. R. (1993). Odorant-selective genes and neurons mediate olfaction in C.elegans. Cell 74:515-527.
3. Wayne A., Fromm H. (2001). Calmodulin as a versatile calcium signal tranducer in plants. New Phytologist 151:35-66
4. Heil M., Karban R. (2010). Explaining evolution of plant communication by airborne signals. Trends in ecology and evolution 25: 137-144.
5. Buck L, Axel R. (1991). A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell 65 (1): 175–87.
6. Rinaldi A. (2007). The scent of life. The exquisite complexity of the sense of smell in animals and humans. EMBO Rep. 8 (7): 629–33.
7. Hallem EA, Dahanukar A, Carlson JR. (2006). Insect odor and taste receptors. Annu. Rev. Entomol. 51: 113–35.
8. Spehr M., Schwane K., Riffell JA., Zimmer RK, Hatt H. (2006). Odorant receptors and olfactory-like signalling mechanisms in mammalian sperms. Mol. Cell Endocrinol. 250 (1-2): 128-136
9. Olsson P., Laska M. (2010). Human male superiority in Olfactory Sensitvity to the sperm attractant odorant, Bougeonal. Chemical senses 10: 1-5.
10.MarcSpehr., Katlen S., Jeffrey A. Riffell, Jon Barbour, RichardK.Zimmer, Eva M. Neuhaus., Hanns H. (2004). Particulate Adenylate Cyclase Plays a Key Role in Human Sperm Olfactory Receptor-mediated Chemotaxis. Journal of Biochemistry 279: 40194-40203.
44
11.Malnic B., Godfrey PA., Buck LB. (2004). The human olfactory receptor gene family. Proc Natl Acad Sci USA 101:2584-2589.
12.Glusman G., Yanai I., Rubin I., Lancet D. (2001). The complete human olfactory subgenome. Genome Res 11:685-702.
13.Zhang X., Firestein S. (2002). The olfactory receptor gene superfamily of the mouse. Nat. Neuroscience 5: 124-133.
14. Godfrey PA., Malnic B., Buck LB. (2004). The mouse olfactory receptor gene family. Proc Natl Acad Sci USA 101:2156-2161.
15.Young JM., Friedmam C., Williams EM., Ross JA., Tonnes-Priddy L., Trask BJ. (2002). Different evolutionary processes shaped the mouse and human olfactory receptor gene families. Human Mol. Genet. 11: 535-546.
16.Richard A. Gibbs, George. MW. (2004). Rat Genome Sequencing Project Consortium: Genome sequence of the Brown Norway rat yields insights into mammalian evolution. Nature 428:493-521.
17.Quignon P., Kirkness E., Cadieu E., Touleimat N., Guyon R., Renier C., Hitte C., Andre C., Fraser C., Galibert F. (2003). Comparison of the canine and human olfactory receptor gene repertoires. Genome Biol 4:R80.
18.Olender T, Fuchs T, Linhart C, Shamir R, Adams M, Kalush F, Khen M, Lancet D (2004). The canine olfactory subgenome. Genomics 83:361-372.
19.Pascale Q., Mathieu G., Maud R. , Patricia L., Sandrine Tache E M. (2005). The dog and rat olfactory receptor repertoires, Genome Biology 2 6:R83.
20.Glusman G, Bahar A, Sharon D, Pilpel Y, White J, Lancet D (2000). The olfactory receptor gene superfamily: data mining, classification, and nomenclature. Mamm. Genome 11 (11): 1016–2.
45
21.Amoore J. E., Hautala E. (1983). Odor as an aid to chemical safety: odor thresholds compared with threshold limit values and volatilities for 214 industrial chemicals in air and water dilution. J. Appl. Toxicol. 3: 272–290.
22.Gaillard I., Rouquier S., Giorgi D. (2003). Olfactory Receptors. Cellular and Molecular Life Sciences 61: 456-469.
23.Stuart F. (2001). How the olfactory system makes sense of scents. Nature 413: 211-218.
24.Frank Zufall, Trese Lienders Zufall (2000). The Cellular and Molecular basis of Odor Adaptation. Chemical Senses 25:473-481.
25.David.T.Moran, Carter Rowley J, Bruce W. Jafek, Mark A.Lowell (1982). The fine structure of olfactory mucosa in man. Journal of neurocytology 11:721-746.
26.Gabriele V. Ronnett1 and Cheil Moon (2002). G-proteins and olfactory signal transduction. Annual Review of Physiology 64: 189-222.
27.Breer H, Shepherd G.M (1993). Implications of the NO/cGMP system for olfaction, Trends in Neurosciences 16:5-9.
28.Kroner C., Boekoeff .I, Lohmann S.M, Breer H (1996). Regulation of olfactory signalling via cGMP dependent protein kinase. European Journal of Biochemistry 236: 632-637.
29.Moon C., Jaberi. P,Otto Bruc A., Baehr W., Ronett. G.V. (1998). Calcium sensitive particulate guanyl cyclase as modulator cAMP in olfactory receptor neurons. Journal of Neurosciences 18: 3195-3205.
30.Dawson T.M., Arriza J.L., Jaworsky D.E., Borisy F.F., Ronett G.V. (1993). β-adrenargic receptor kinase 2 and β-arrestin 2 as mediators of odorant induced desensitization. Science 259: 825-829.
46
31.Peppel K., Boekhoff I., Mc Donald P., Breer H., Caron M.G. (1997). G-protein coupled receptor kinase 3 (GSK3) gene disruption leads to loss of odorant receptor desensitization. The Journal of Biological Chemistry 272: 25425-25428.
32.Robert R. H. Anholt , Ann M. Rivers (1990). Olfactory transduction: cross-talk between second-messenger systems. 29 (17): 4049-4054.
33.Jonathan B., Wolfgang B., King-Wai Y., Stephan F. (2004). Calmodulin permanently associates with rat olfactory CNG channels under native conditions. Nature Neruoscience 7: 705-710.
34.Jones DT., Reed RR (1989). Golf: an olfactory neuron specific-G protein involved in odorant signal transduction. Science 244: 790-795.
35.Robert RH. , Susanne M. Mumby , Doris A. Stoffers , Peggy R. Girard , JF. Kuo , Solomon H. Snyder (1987). Transduction proteins of olfactory receptor cells: identification of guanine nucleotide binding proteins and protein kinase C. Biochemistry 26 (3): 788-795.
36.Jia Wei, Allan Z. Zhao, Guy CK. Chan, Lauren P. Baker, Soren Impey, Joseph A. Beavo, Daniel R. Storm (1998). Phosphorylation and Inhibition of Olfactory Adenylyl Cyclase by CaM Kinase II in Neurons: a Mechanism for Attenuation of Olfactory Signals. Neuron 29: 495-504.
37.Allen MS.(1987). Signal transduction by guanine nucleotide binding proteins. Molecular and cellular endocrinology 49: 1-16.
38.Zagotta W.N, Siegelbaum SA. (1996). Structure and Function of Cyclic Nucleotide-Gated Channels. Annual Review of Neuroscience 19: 235-263.
39.Lisa J Brunet, Geoffrey G., John N. (1996). General Anosmia Caused by a Targeted Disruption of the Mouse Olfactory Cyclic Nucleotide–Gated Cation Channel. Neuron 17: 681-693.
47
40.Aaron B. Stephan, Eleen Y. Shum, Sarah Hirsh, Katherine D. Cygnar, Johannes Reisert, and Haiqing Zhao (2009). ANO2 is the cilial calcium-activated chloride channel that may mediate olfactory amplification. PNAS 106 : 11776-11781.
41.Boekhoff I, Touhara K, Danner S, Inglese J, Lohse MJ, Breer H, Lefkowitz RJ. (1997). Phosducin, potential role in modulation of olfactory signalling. The Journal of Biological Chemistry 272(7): 4606-4612.
42.Trese Leinders-Zufall, Renee E. Cockerham, Stylianos Michalakis , Martin Biel, David L. Garbers , Randall R. Reed , Frank Zufall , and Steven D. Munger (2007). Contribution of the receptor guanylyl cyclase GC-D to chemosensory function in the olfactory epithelium. PNAS 104 (36): 14507–14512.
43.Pathania M. (2012). Molecular Understanding of Brain : Analysis of Long TermPotentiation(LTP ). M.Sc dissertation submitted to Madurai Kamaraj University 1–54.
44.Ramamoorthy (2012). Molecular Understanding of the Brain: Analysis of Phototransduction pathway. M.Sc dissertation submitted to Madurai Kamaraj University 1–46.
45.Mulakrishanan A. (2013). Structural genomics initiative for brain molecules and enhancing molecular database of brain. M.Sc dissertation submitted to Madurai Kamaraj University 1-61.
48
SUPPLEMENTARY TABLE
ORGANISMS MOLECULES KEGG Accession No.
Homo sapiens GNAL 2774 ADCY3 109 CNGA3 1261 CNGA4 1262 CNGB1 1258 CALM1 801 CALM2 805 CALM3 808 CAMK2A 815 CAMK2B 816 CAMK2D 817 CAMK2G 818 CLCA1 1179 CLCA2 9635 CLCA4 22802 GUCA1A 2978 GUCA1B 2979 GUCA1C 9626 PRKG1 5592 PRKG2 5593 PRKX 5613 PRKACA 5566 PRKACB 5567 PDC 5132 Pan troglodytes GNAL 455285 ADCY3 459070 CNGA3 470517 CNGA4 466413 CNGB1 454126 CALM1 738819 CALM2 459213 CALM3 466008 CAMK2A 462189 CAMK2D 461445 CAMK2G 738330 CLCA1 456640 CLCA2 736939 CLCA4 469372 GUCA1A 748651 GUCA1B 427007 GUCA1C 460572 PRKG1 744467
49 PRKG2 461217 PRKX 449495 PRKACA 455776 PRKACB 469367 PDC 469614 Mus musculus GNAL 14680 ADCY3 104111 CNGA3 12790 CNGA4 233649 CNGB1 333329 CALM1 12313 CALM2 12314 CALM3 12315 CAMK2A 12322 CAMK2B 12323 CAMK2D 108058 CAMK2G 12325 CLCA1 12722 CLCA2 80797 CLCA4 229927 GUCA1A 14913 GUCA1B 107477 PRKG1 19091 PRKG2 19092 PRKX 19108 PRKACA 18747 PRKACB 18749 PDC 20028 Canis familiaris GNAL 607658 ADCY3 482994 CNGA3 481341 CNGA4 476830 CALM1 480416 CALM2 474584 CALM3 476428 CAMK2A 612941 CAMK2D 610764 CAMK2G 489050 CLCA1 490178 CLCA2 612510 CLCA4 612508 GUCA1A 609180 GUCA1B 481801 GUCA1C 487968 PRKG1 609616 PRKG2 100855930
50 PRKX 610031 PRKACA 403556 PRKACB 479975 PDC 403624 Bos taurus GNAL 100124520 ADCY3 535603 CNGA3 281701 CNGA4 516000 CNGB1 281702 CALM1 617095 CALM2 100297344 CALM3 520277 CAMK2A 530719 CAMK2B 525416 CAMK2D 532713 CAMK2G 282162 CLCA1 507643 CLCA2 534256 CLCA4 507504 GUCA1A 282243 GUCA1B 286851 PRKG1 282004 PRKG2 533330 PRKX 505773 PRKACA 282322 PRKACB 282323 PDC 287007 Gallus gallus GNAL 474379 ADCY3 422013 CNGA3 396144 CNGA4 430630 CNGB1 768759 CALM1 396523 CALM3 416692 CAMK2A 374175 CAMK2B 374174 CAMK2D 422688 CAMK2G 423737 CLCA2 771009 CLCA4 424523 GUCA1A 374216 GUCA1B 419864 GUCA1C 427960 PRKG1 423682 PRKG2 428753 PRKX 481656
51 PRKACB 424542 PDC 429078 Xenopus laevis GNAL 378630 CALM1 606721 CALM2 380558 CAMK2A 399284 CAMK2B 397789 CAMK2D 399160 CAMK2G 380508 GUCA1A 44645 PRKG2 444025 PRKACA 100101276 PRKACB 380388 Danio rerio GNAL 492467 ADCY3 571825 CNGA3 100334711 CNGA4 568123 CNGB1 568211 CALM1 406660 CALM2 336121 CALM3 327379 CAMK2A 550436 CAMK2B 562064 CAMK2D 445208 CAMK2G 791855 CLCA1 334289 GUCA1A 140430 GUCA1B 140431 PRKG1 394005 PRKG2 100125912 PRKACA 564064 PRKACB 100538234 PDC 386613
Supplementary Table shows KEGG accession number for nucleotide sequences of the molecules considered for the pathway analysis for different organisms.
52
APPENDIX A
Cloning of endolysin (ybcS) gene in Loop 5 of Outer
Membrane Protein F (OmpF)
53
A1 INTRODUCTION
A1.1 Membrane Proteins
Biological membranes exist as a dynamic structure allowing the cells to communicate with its environments. It has two essential components, i.e. lipids and proteins. Membrane proteins are interesting scientifically because of their key roles in various biological processes of life. They control cell adhesion to form tissues, help in development of plants and animals, control important metabolic processes including salt balance, energy production and transmission, and photosynthesis. Nowadays, they are also targets of most of the pharmaceuticals and becoming important across medicine and agriculture.
An insight into the protein database indicates that approximately 20% of all the genes in most of the genomes encodes for membrane proteins [1]. Broadly, membrane proteins are classified as:
Intergral membrane proteins Peripheral membrane proteins.
Peripheral membrane proteins are attached to the lipid bilayer either by electrostatic or Van der Waals interactions or by covalent interactions [2]. Integral membrane proteins are permanently attached to the membrane and span the membrane [3]. Basically, integral membrane proteins are also of two types-
α-Helical β-barrels.
α-helical membrane proteins are most abundantly found in all types of biological membranes and contain one or several α-helices. The helices are hydrophobic in nature.
β-barrel proteins are mainly found in outer membranes of Gram-negative bacteria, lipid rich cell walls of some Gram-positive bacteria and in outer membranes of mitochondria and chloroplasts. They consist of β-sheets and form a β-barrel like structure. The residues alternatively face outwards towards the lipids or inwards towards the proteins such that they form a pattern having alternatively hydrophobic and polar. Thus a porous channel is formed through which water-soluble molecules can cross. It has been estimated that about 2-3% of all the genes in Gram-negative bacteria encodes for β-barrel membrane proteins [4].
54
A1.2 Porins
Porins belong to the β-barrel class of membrane proteins which were discovered in 1976 by T. Nakae in Salmonella as protein complexes that forms trans-membrane channels through which molecules can diffuse. These are large channels that are specific to different types of molecules. These are present in Gram-negative bacteria, Gram-positive bacteria of the group Mycolata and in mitochondria and chloroplasts of eukaryotes [5]. Generally, porins are present as oligomers mostly trimers [6].
OmpF. OmpC and PhoE (phosphorin) are generally referred as ‘Classical porins’. They show a general preference for charge and size of solute, OmpC and OmpF preferring cations over anions and PhoE preferring anions [7].
X-ray crystallization studies have shown that porins consists of antiparallel β- strands, the number of which differs in different porins.
A1.3 OmpF
The outer membrane protein F (OmpF) is one of the three major porins found in Gram-negative bacteria. It is a non specific transport channel that allows passive diffusion of small, polar molecules like water, ions, glucose and waste products, of 600-700 Da in size through the cell’s outer membrane [8, 9]. OmpF porins functions to regulate osmotic pressure between cell and its surroundings.
The OmpF porin gene contains 1086 nucleotides which correspond to 362 amino acids. Out of these 362 amino acids, 22 amino acids make up the signal peptide ad form a precursor sequence to OmpF porins which are eventually cleaved off [10].
OmpF of Salmonella typhii has 37KDa molecular weight and it is made up of 371 amino acids. It shows 57.4% identity with E.coli OmpF. The structure of S.typhi OmpF was solved and submitted to PDB and the PDB ID is 3NSG [11].
OmpF has 16 antiparallel β-strands and 8 loops that connect β-helices to each other. Contact among the monomers is stabilized by hydrophobic and polar interaction, where Loop 2 (known as the ‘latching loop’) tends to bend over the wall of the barrel of the neighbouring
55
subunit, playing a significant role in stabilization. The pore size is however reduced by the insertion of Loop 3 into the barrel, forming the “eyelet” [12].
Figure A1 Structure of OmpF porin
monomer.
56
Figure A2 Structure of OmpF porin of Salmonella typhii as a trimer where red regions shows loop 5 of OmpF.
A1.4 Clinical and Industrial manifestations of Porins
The exposed external loops of porins are known to interact with antibodies, phages and colicins. OmpF, along with LPS, is the receptor for phage K20 [13, 14].
Porins also acts as potential target as an antigen that can further used for vaccination. Porins predominantly induce the production of IgG and IgM antibodies. It has been reported that OmpC is one of the major antibody inducer and confers a stronger long lasting bactericidal activity than OmpF [15].
It has also been showed that E.coli porins can be used as bioadsorbents as Yeast metallothioneins can be effectively expressed in E.coli as attachments to LamB protein which enhances E.coli natural capability to cadmium ions to about 15-20 fold [16].
Bacterial outer membrane porins can also be used as efficient drug delivery system. OmpF can help in diffusion of a prodrug into vesicles acting as functional nanoreactor [17].
57
A1.5 Endolysin
Endolysin, generally termed as “lysins” are double stranded DNA bacteriophage encoding enzymes produced during the late phase of gene expression in the lytic cycle. It degrades the peptidoglycan layer which is the main component of bacterial cell wall, enabling the liberation of progeny virus [18].
Endolysins lack secretory signals, thus their access to peptidoglycans from inside the cells is dependent on small hydrophobic proteins called, Holins, which enable endolysin molecule to cross the inner membrane. Holins themselves do not have any enzymatic activity and are localized to the cytoplasmic membrane.
The endolysin gene used for the study is ybcS which is found in defective prophage, DLP12 of E.coli K-12 as a part of two component lysis cassette along with essd, a putative holin. Endolysin structure of Bacillus anthracis is known and it contains an N-terminal amidase and a C-terminal membrane binding site. Some contain an N-terminal SAR (Signal Arrest Release) domain, that aids in holin independent release of the protein into the periplasmic space [18].
58
A2. Materials and Methods
A2.1 Plasmid isolation by Alkaline Lysis Method
Requirements- Solution I 50 mM glucose
25 mM tris chloride (pH 8.0) 10 mM EDTA (pH 8.0)
This solution was filter sterilized and stored at 4°C. Solution II
0.2N NaOH (freshly diluted from 10N stock)
1% (w/v) SDS
This solution was freshly prepared and used at room temperature. Solution III
3M sodium acetate (pH 5.2, adjusted using glacial acetic acid) This solution was autoclaved and stored at 4°C.
Procedure
Inoculation of bacterial culture was done in LB broth (10ml) with their respective antibiotics i.e. Kanamycin (50µg/ml) for pOKompFL5H+E and ampicillin (100µg/ml).
Overnight culture was then pelleted out at 12000 rpm for 5 minutes.
The pellet was then suspended in 200µl of ice-cold Solution I by vigorous vortexing.
Then 400µl of freshly prepared Solution II was added and the contents were mixed gently by inversion, and incubated on ice for 5 minutes.
59
Then 300µl of ice-cold Solution III was added, mixed by inversion and stored on ice for 5 minutes.
Then after centrifugation at 12000 rpm for 10 minutes, supernatant was collected in a fresh eppendorf.
DNAse free RNAse was then added at a concentration of 2µg/ml and incubated at 370C for 45 minutes.
Then equal volume of chloroform was added and mixed by inversion.
It was then spun at 12000 rpm for 5 minutes and the upper aqueous layer was collected in a fresh tube.
After this, equal volume of isopropanol was added and the tubes were incubated for 10 minutes at room temperature.
It was again centrifuged at 12000 rpm for 15 minutes.
The pellet was then washed with 70% ethanol and the mixture was again centrifuged for 5 minutes at 12000 rpm.
After then, the pellet was kept for air drying.
Finally, the pellet was resuspended in 20µl of MilliQ and then stored at -200C.
A2.2 Agarose Gel Electrophoresis
The isolated plasmid was then checked by using Agarose Gel Electrophoresis. Running buffer (1X TAE)
• Tris : 0.025 M
• EDTA : 1 mM
The pH was adjusted to 8.0 using acetic acid.
Sample buffer (10X)
• Glycerol : 50%
• Bromophenol blue : 0.0%
60
1 % Agarose gel was casted with the addition of ethidium bromide (1μl/mL) in 1 X TAE buffer. Samples were loaded with the loading dye. Electrophoresis was carried out at 100V till the dye front reaches the end of gel. After electrophoresis the gel is visualized under UV transilluminator.
A2.3 Restriction Digestion
Double digestion and sequential digestion were performed and the digested products were analyzed by agarose gel electrophoresis.
Sequential Digestion
Digestion mixture was prepared by adding DNA, enzyme I, buffer and MilliQ and the mixture was then spun at 12000 rpm for 1 minutes for completely mixing of the contents.
The digestion mix was kept in water bath for 3 hours at 370C.
After then, one-tenth volume of sodium acetate (pH 5.2) and twice the volume of isopropanol were added.
It was then incubated at -200C for one hour.
Then it was centrifuged at 14000 rpm for 10 minutes.
The supernatant was discarded and the mixture was washed with 80% ethanol. It was again spun for 14000 rpm for 10 minutes.
The supernatant was removed and the pellet was air dried completely.
The pellet was again resuspended in Enzyme II, buffer and MilliQ according to the reaction volume and then the contents were again mixed by spinning at 12000 rpm for 1 minute.
The mixture was again incubated in water bath at 370C for 3 hours.
The digestion mixture was finally run in agarose gel to analyze the digested products.
A2.4 Gel Elution
The agarose block containing the desired DNA fragment was taken and the DNA was eluted by using SIGMA GelEluteTM Gel Extraction Kit as per the manufacturer’s instructions.
61
A2.5 Ligation
Reactions were set up with 5 Weiss Unit of T4 DNA Ligase (Fermentas). Ligation was allowed to take place at 40C over.
A2.6 Competent Cell Preparation by Calcium Chloride Method
A single colony of E. coli DH5α, pET20b+ or BL21DE3 was inoculated into 2 ml LB broth and kept in a shaker at 180 rpm at 370C overnight.
Starter culture (1%) was then subcultured into 50ml of LB medium and was grown again at 370C for two hours.
When O.D. reaches to 0.6. the culture is taken out and kept in ice for 10 minutes. Then the culture is spun at 5000rpm for5 minutes at 40C.
The supernatant was then discarded and the pellet was suspended into ice-cold 1ml 100mM calcium chloride followed by gentle shaking.
Then the volume was made upto 7ml by adding more calcium chloride. Then it was again centrifuged at 4000 rpm for 5 minutes at 40C.
After discarding the supernatant again 1ml of calcium chloride was added to the pellet and was mixed by gentle shaking. The volume was made upto 7ml by adding more calcium chloride,
It was then incubated for half an hour on ice and then centrifuged at 3000 rpm for 5 minutes at 40C.
Finally, the pellet was resuspended into 500µl of calcium chloride.
A2.7 Transformation
100μl of competent cells were taken in a 1.5 mL eppendorf.
To this 100ng of plasmid DNA to be transformed was added or the ligation mixture can be added.
The mixture was then incubated in ice for 30’.
The heat shock was then given for 90 seconds at 42°C in a water bath and immediately the tubes are transferred in ice.
62
It was then left in the shaker for expression for 90 minutes at 370C at 180 rpm.
The cells were then plated on LB-agar plates with suitable selection conditions and incubated at 370C in an incubator overnight.
A2.8 Quick screening
An overnight culture was taken in 1.5 ml eppendorf and pelleted at 12000rpm for 5 minutes.
The pellet was then suspended into 20µl of MilliQ by vortexing.
Then 50µl each of DNA loading dye and phenol was added to the contents. The mixture was again vortexed strongly.
It was again centrifuged at 14000 rpm for 10 minutes.
After this three layers will be formed. The uppermost layer contains plasmid DNA was taken and check by Agarose Gel Electrophoresis.
A2.9 Protein expression
SDS-PAGE Separating gel (10 ml) - 12% 30% acrylamide – bisacrylamide : 4 ml Tris HCl (pH 8.8 and 1.5M) : 2.5 ml SDS 10% : 0.1 ml APS 10% : 0.1 ml TEMED : 4 µl Distilled water : 3.4 ml
63 Stacking Gel (10 ml) - 5% 30% acrylamide – bisacrylamide : 1.7 ml Tris HCl (pH 6.8 and 1M) : 2.5 ml SDS 10% : 0.1 ml APS 10% : 0.1 mg TEMED : 8 µl Distilled water : 5.7 ml
Sample loading Buffer (2X)
Tris-HCl (pH 6.25) : 0.125 Bromophenol blue : 0.05% SDS : 4% Glycerol : 20% β-mercaptoethanol : 10% Running Buffer (pH 8.3) Glycine : 0.192 M Tris-HCl : 0.025 SDS : 0.1% Staining Solution (0.1%)
Coomassive Brilliant Blue (G 250) : 0.25%
Methanol : 50%
Acetic acid : 10%
Destaining Solution Acetic acid 10%
64
The SDS PAGE was performed with 12% separating gel and 5% stacking gel. A separating gel was cast in the gel assembly system and a small column of stacking gel was poured over it and an appropriate comb was placed and kept for polymerization. Samples were prepared by boiling with an equal volume of the sample buffer for 5 minutes. Electrophoresis was done at 100V, till the dye front reached bottom of the gel. After electrophoresis, the gel was kept in staining solution (Coomassie staining) for 1 hr and the excess stain was removed by keeping in the destaining solution till the bands were clear.
65
A3. RESULTS AND DISCUSSION
A3.1 Introduction of endolysin (
ybcS
) into loop 5 of OmpF
In this study, engineering of ybcS (endolysin)gene into loop 5 of OmpF was done. Loop 5 is one of the loops of OmpF that is away from the trimer interface. So, it is taken for engineering studies as it will not affect in trimer formation.
Further, endolysin is an unstable enzyme, so its engineering into loop 5 will provide stability to its structure which can be further be utilized for various applications including endolysins as an antimicrobial agent.
ybcS was obtained as plasmid from Genobase-ASKA library. It has an internal EcoRV site in its reading frame which was removed by inverse PCR mutating the restriction site and keeping the codon intact [19, 20]. It was cloned into TA cloning vector with HindIII at 5’ end and EcoRV site at 3’ end. After sequencing TA clone showed deletion mutation of two CC nucleotides within the endolysin which was again corrected by inverse PCR. Now the TAendolysin CC corrected has 549 bp endolysin as insert which is released after HindIII and EcoRV digestion. pOKOmpF-YFP in loop F construct was used as backbone.
So, pTA-endolysinCC corrected and pOKOmpFl5-YFP was used for this part of study.
66
A3.1.2 Isolation of plasmid DNA
To isolate endolysin from TA vector, plasmid DNA of pTAendolysinCC corrected was isolated.
For the engineering of endolysin into OmpF, plasmid DNA of pOKOmpFL5-YFP was also isolated.
The isolated plasmids are shown in Figure A4.
Lane 1: pTAendolysinCC corrected Lane 2: pOKompFL5-YFP
Figure A4 Plasmid isolation.
A3.1.3 Restriction Digestion
pTA-endolysinCCcorrected has HindIII and EcoRV restriction sites engineered at its 5’ and 3’ end of endolysin gene. TA vector also has two HindIII sites. So, sequential digestion was performed for pTA-endolysinCC corrected. The expected bands were 2.7 kb for TA backbone and 549 bp of endolysin.
pOKOmpFL5-YFP also has HindIII and EcoRV sites engineered in its loop 5. So, to obtain backbone for the insertion of endolysin vector, double digestion was performed for pOKOmpFL5-YFP. After digestion the backbone of pOKOmpFL5 was expected to give band of 3.2 bp length.
67
Both the restriction digestion results gave the expected band patterns which are shown in Figure A5.
Lane 1: Undigested pTA-endolysinCC corrected
Lane2: pTA-endolysinCC corrected digested with HindIII & EcoRV Lane 3: 1 Kb ladder
Lane 4: pOKOmpFL5-YFP digested with HindIII and EcoRV Lane 4: Undigested pOKOmpFL5-YFP
Figure A5 Restriction digestion products. 1 2 3 4 5