Neuron-Like Differentiation of Bone Marrow-Derived
Mesenchymal Stem Cells
Keum Seok Bae,
1Joon Beom Park,
1Hyun Soo Kim,
2,4Dae Sung Kim,
1Dong Jun Park,
3and Seong Joon Kang
1Departments of 1Surgery, 2Hemato-Oncology and 3Otorhinolaryngology, Yonsei University Wonju College of Medicine, Wonju, Korea; 4FCB-Pharmicell Co., Ltd. Seongnam, Korea.
Received: April 17, 2010 Revised: July 25, 2010 Accepted: July 26, 2010
Corresponding author: Dr. Seong Joon Kang, Department Surgery, Yonsei University Wonju College of Medicine, 162 Ilsan-dong, Wonju 220-701, Korea.
Tel: 82-33-741-1306, Fax: 82-33-742-1815 E-mail: mdkang@yonsei.ac.kr
∙ The authors have no financial conflicts of interest.
© Copyright:
Yonsei University College of Medicine 2011
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/ licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Purpose: Mesenchymal stem cells (MSCs) are multipotent and give rise to distinct-ly differentiated cells from all three germ layers. Neuronal differentiation of MSC has great potential for cellular therapy. We examined whether the cluster of mechani-cally made, not neurosphere, could be differentiated into neuron-like cells by growth factors, such as epidermal growth factor (EGF), hepatocyte growth factor (HGF), and vascular endothelial growth factor (VEGF). Materials and Methods:BMSCs grown confluent were mechanically separated with cell scrapers and masses of sepa-rated cells were cultured to form cluster BMSCs. As described here cluster of BM-SCs were differentiated into neuron-like cells by EGF, HGF, and VEGF. Differenti-ated cells were analyzed by means of phase-contrast inverted microscopy, reverse transcriptase-polymerase chain reaction (RT-PCR), immunofluorescence, and immu-nocytochemistry to identify the expression of neural specific markers. Results: For the group with growth factors, the shapes of neuron-like cells was observable a week later, and two weeks later, most cells were similar in shape to neuron-like cells. Par-ticularly, in the group with chemical addition, various shapes of filament structures were seen among the cells. These culture conditions induced MSCs to exhibit a neu-ral cell phenotype, expressing seveneu-ral neuro-glial specific markers. Conclusion:
bone marrow-derived mesenchymal stem cells (BMSCs) could be easily induced to form clusters using mechanical scraping, not neurospheres, which in turn could dif-ferentiate further into neuron-like cells and might open an attractive possibility for clinical cell therapy for neurodegenerative diseases. In the future, we consider that neuron-like cells differentiated from clusters of BMSCs are needed to be compared and analyzed on a physiological and molecular biological level with preexisting neu-ronal cells, and studies on the possibility of their transplantation and differentiation capability in animal models are further required.
Key Words: Neuron-like cells, mesenchymal stem cell, epidermal growth factor, vascular endothelial growth factor, hepatocyte growth factor
INTRODUCTION
non-into a blood vessel in a bone marrow transplantation model.7
Mezey, et al.8 also discovered that when bone marrow cells
are injected into mouse peritoneum, these cells migrate to the brain and differentiate into neurons.
Recent studies also report that these marrow-derived mes-enchymal cells have a capacity to differentiate into neurons in ex vivo surroundings. Sanchez-Ramos, et al.20 used
retino-ic acid and brain-derived neurotrophretino-ic factors to differentiate BMSCs into neural cells, including neurons and astrocytes. Woodbury, et al.21 determined that using antioxidants such as
β-mercaptoethanol (BME), dimethyl sulfoxide (DMSO), and butylated hydroxyanisole (BHA), they differentiated mar-row-derived mesenchymal cells into neurons. The result was that within a few hours, most MSCs (80%) transformed into neuron-like shapes and expressed neuron-specific markers.
In the studies mentioned above concerning the potential of BMSC differentiation into neuron-like cells, the results from these studies suggest that these MSCs can be applied to the treatment of various brain and nerve diseases. There-fore, the goal of this study is to determine an effective meth-od of differentiating BMSCs into neuron-like cells. We used EGF, vascular endothelial growth factor (VEGF), and hepa-tocyte growth factor (HGF), and observed the conditions of differentiation.
EGF and VEGF are known to function as growth factors that stimulate proliferation of brain tissue-derived neural mesenchymal cells in culture.22-26 HGF is a heterodimer that
consists of a chain containing four kringles and a serine protease-like b chain and is also called as scatter factor. It binds to c-Met, the tyrosine kinase receptor,27 and it is a
growth factor that has pleiotrophic roles.28 Above all, within
the neural tissue, tyrosine kinase receptor and its ligands are expressed, and they play an important role in survival, dif-ferentiation and regeneration of neurons.
The ability of these cells to neurogenically differentiate has deep potential that is applicable to the field of cell ther-apy. Nerve tissue has a limited ability to repair itself after injury.29-32 Generally, groups of cells suggested for
neuro-genic cell therapy are embryonic stem cells and neuron stem cells acquired in embryo or adult brain tissue.33-36 However,
the use of these cells has limited application in clinical cir-cumstances due to ethical and legal issues.
BMSCs and their ability to differentiate into neuron-like cells play an important role in the treatment of degenerative nerve diseases such as Parkinson’s disease, Huntington’s disease, and Amyotrophic Lateral sclerosis.37-39 It is being
recognized that bone marrow that functions as storage for hematopoietic stem cells besides the most common stem
cells of hematopoietic origin. In 1968, Friedenstein et al proved that these cells can be formed into bone, and in 1985 Owen1 reported that cells with self renewal and
multi-differentiation capacity may be present in bone marrow on the basis of these cells’ similarity to hematopoietic cells. These have been named as bone marrow derived mesen-chymal stem cells (MSCs), or marrow stromal stem cells, and there are many researches who focus their aims on these cells.
MSCs are harvested from fat, cord blood, and embryos as well as from bone marrow. They have the potential to dif-ferentiate into marrow stromal cells, fat cells, osteoblastic cells, chondrocytes, tendinocytes, and myocytes, which are normally derived from mesenchymal stem cells.2-13 They
are also capable of differentiating into endodermal origin hepatocytes and ectodermal origin neurons. Therefore, these differentiation potentials of MSCsare used for research into possible clinical utilization in regenerative cell therapy.
Generally, the number of MSCs in the bone marrow is small. There are about 2 to 5 cells present in every 1×106
mononuclear cells. Normally, the human body is thought to have about 1×106 MSCs. However, they are relatively
easi-ly separated and grown in culture, and they have a great proliferation capability of a billion-fold increase in number
ex vivo without losing their stem cell characteristics. Nota-bly, Kuznetsov, et al.14 identified several growth factors that
are related to MSCs. They reported that MSCs under the presence of serum, platelet derived growth factor (PDGF), basic fibroblast growth factor (bFGF), transforming growth factor β (TGF-β), and epidermal growth factor (EGF) can form in vitro colonies.15
The Ancient Chinese once said that the “brain is the sea of bone marrow.” These ancient beliefs are now being prov-en one by one in recprov-ent times.16 Eglitis and Mezey17
report-ed bone marrow derivreport-ed cells discoverreport-ed in the all brain parts from the cortex to the brain stem. Particularly, in the case of human MSCs, they migrate and survive similarly to mice astrocytes when grafted in mice striatum, and they are discovered to lose their marrow mesenchymal cell mark-ers.18 When marrow-derived mesenchymal cells are
trans-planted into the lateral ventricles of a mouse, Kopen, et al.19
ROTEC), CD73-PE (BD), CD45-FITC (BD), CD34-FITC (BD), CD14-FITC (BD), and control (IgG1-FITC and IgG1-PE, BD)] for 45 min at room temperature. They were then washed three times with PBS and precipitated. The cells were dripped into columns and fixed with flow buffer (0.1% sodium azide, 1% paraformaldehyde, 0.5% bovine serum albumin). Finally, the cells were analyzed under Ep-ics XL (Beckman Coulter, Brea, CA, USA).
Differentiation capability test
Osteogenic differentiation of BMSCs
BMSCs (second passage) in the early phase of culture were suspended to DMEM. They were divided into 24 well plates (5×104 cells/well) and cultured for 24 hours. Then, the
me-dium was replaced with osteoblast differentiation meme-dium [High-glucose DMEM, 10% fetal bovine serum, 0.1 µM dexamethasone (Sigma, St. Louis, MO, USA), 10 mM β-glycerol phosphate, 50 µM L-ascorbic acid] and cultured for three weeks. The medium was replaced twice a week.
Chondrogenic differentiation of BMSCs
BMSCs were suspended in chondrogenic medium (6.25 μg/mL insulin, 6.25 μg/mL transferrin, 6.25 μg/mL sele-nous acid, 5.35 μg/mL linoleic acid, 1.25 μg/mL bovine se-rum albumin), 1 mM Pyruvate, 37.5 μg/mL Ascorbic acid, 10-7 M dexamethasone (Sigma)]. It was then centrifuged for 10 minutes in 450 xg. The precipitated cells were cul-tured in chondrogenic medium for three weeks as micro-mass. The medium was replaced for twice a week, and later stained to identify differentiation.
Adipogenic differentiation of BMSCs
BMSCs were suspended in DMEM, and divided into six well plates. These were cultured for 48 hours. The cultured cells were grown for three additional days under 100% den-sity. Adipogenic induction medium [high glucose DMEM, 10% fetal bovine serum, 1 μM dexamethasone (Sigma), 10 ug/mL insulin (Sigma), 100 μM indomethacin (Sigma), and 0.5 mM methyl-isobutylzanthine (Sigma)] was used for a 72-hour culture and to induce differentiation. The medium was removed and replaced with adipogenic maintenance medium [high glucose DMEM, 10% fetal bovine serum, 10 ug/mL insulin (Sigma)] to culture for 24 hours. The re-placement of the media was repeated for three times. Then, finally, the sample was cultured for one additional week with adipogenic maintenance medium.
hematopoietic stem cells and MSCs can replace embryonic stem cells in protection and cell therapy of dying nerves.40-43
In fact, acquiring autologous stem cells, inducing less pain in patients, proliferating stem cells in vitro, differentiating, and transplanting cells back into the same patient is a new concept, and has interesting possibility as a new treatment method.
In this study, we used growth factors usually found in the human body to induce differentiation to neuron-like cells. We expect that this revolutionizing autologous cell therapy will be promising for the treatment of neurodegenerative diseases.
MATERIALS AND METHODS
Bone marrow collection and mesenchymal stem cell culture
Patients with no notable pathologic history were chosen for this study. Human bone marrow of three healthy donors aged 21 to 40 years was obtained from Institute of cell therapy center before 2005, after they gave informed consent accord-ing to the approved procedure. About 10 mL of marrow were collected from the pelvis of each patient and stored in a hepa-rin containing test tube. 30 mL of phosphate buffered saline (PBS) was added to marrow sample. Then, the mixture was slowly streamed down on Ficoll-Plaque TM plus solution. The mixture was centrifuged using density gradient centrifu-gation for 20 minutes in 2,000 rpm. Next, the top layer and monocyte layer present in the interface of Ficoll-Plaque TM plus were retrieved and mixed again with 20 mL of PBS. Again, the blend was centrifuged in 1,800 rpm for 5 minutes, and this time only the monocyte was retrieved.
The separated monocytes were placed in Dulbecco’s Mod-ified Eagle’s Medium (DMEM, 10 % FBS, 1×penicillin-streptomycin). Then, appropriate amounts of monocytes (2×105) were inoculated into a culture dish (T-75), and
cul-tured in an incubator.
After seven to ten days, since only MSCs in monocytes proliferated on the floor of the culture dish, the medium was replaced. Later, when MSCs proliferated over 80% of the culture dish, it was transferred to other culture dish (T-175) for subculture. The medium was replaced once every three to four days to subculture 5 times.
Fluorescence Activated Cell Sorting (FACS) analysis
(SE-ville, MD, USA), HGF, 20 ng/mL, (R&D Systems, Minne-apolis, MN, USA), VEGF, 20 ng/mL, (R&D Systems), and was cultured with replacement of the medium at 3 day in-tervals for 14 days. The group with chemicals was cultured with the DMEM medium for seven days, and on the eighth day, a chemical compound (BHA 200 µM, KCl 5 mM, Val-proic acid 2 µM, Forskolin 10 µM, Hydrocortisone 1 µM, Insulin 5 µM) was added to the medium for an additional five days of culture.
Immunostaining
Immunochemical staining
The differentiated cells were suspended on a cm2 cover
slide at a 1×104 cells/cm2 concentration. After fixation, the
slide was washed with PBS for 5 minutes and fixed with 4% paraformaldehyde with PBS for 15 minutes. It was then washed twice with PBS for 5 minutes. Later, the sample was processed with PBS containing 1% BSA and 0.2% Tri-ton X-100 for 5 minutes. Primary antibody was mixed and allowed to react for 16 hours. Anti-human neuron-specific enolase (NSE); Chemicon Inc., Billerica, MA, USA), anti-human NeuN (Chemicon Inc.), anti-anti-human β-tubulin III (Sigma Co., St. Louis, MO, USA), anti-human glial fibril-lary acidic protein (GFAP); Sigma Co.), and anti-human microtubule-associated protein-2 (MAP-2) were used for primary antibodies. After the reaction with the primary anti-body, PBS with 0.5% BSA was used to wash twice for 15 minutes. The sample was cultured again with secondary an-tibody for 30 minutes. PBS with 0.5% BSA was used to wash twice for 5 minutes. Afterward, the Avidin-biotin re-action (Vectastain Elite ABC kit; Vector Laboratory Inc., Burlingame, CA, USA) was processed for 30 minutes. It was washed twice for 5 minutes with PBS. 3,3’-diamino-benzidine tetrahydrochloride dehydrate (DAB, Sigma Co.) was mixed as a staining substrate and left to react for five minutes. Processing with PBS for five minutes stopped the reaction and it was further washed with PBS two more times. The sample was dried and then washed with distilled water for five minutes. Finally, it was washed and dehydrat-ed with distilldehydrat-ed water, 70%, 80%, 95%, and 100% ethanol (in that order), and suspended.
Immunofluorescence staining
For immunofluorescent staining, the cells were transferred to a culture slide (four-chamber, BD Falcon, Franklin Lakes, NJ, USA) and cultured for 1-2 days. Using 4%
para-Confirmation of differentiation capability
Alkaline phosphatase staining of osteoblasts
Alkaline phosphatase staining method was used to confirm the differentiation of BMSCs into osteoblasts. Cells that had been cultured in a 24-well plate for three weeks were removed from the media and washed twice with PBS. Ice-cold methanol (99.9%) was added and fixed in room tem-perature for two minutes. Then, it was washed with tertiary distilled water. BCIP/NBP (Sigma) liquid substrate was add-ed and reactadd-ed with the sample for 10 minutes at an ambi-ent temperature. Again, the mixture was washed twice with tertiary distilled water. The stain results were later analyzed under an optical microscope.
Safranin O staining of chondrocytes
The cultured cells were suspended in ice-cold acetone for 1-2 minutes after washing PBS. Mayer’s hematoxylin solu-tion was used to stain for 10 minutes and washed with a bluing solution, and then with distilled water. 0.1% safranin O solution was added and allowed to react for five minutes. Every two minutes, the sample was added to the following solutions in order: 95% ethyl alcohol, absolute ethyl alco-hol, and xylene. The sample was mounted on a slide and their differentiation was studied under a microscope.
Oil red-O staining of adipocytes
Lipid staining using oil red O was used to confirm the dif-ferentiation of adipocytes from BMSCs. Cells that were cultured for 4 weeks in a six-well plate were mixed with 4% paraformaldehyde (in PBS) and fixed for 4 to 12 hours. The sample was then washed with 60% isopropanol (in PBS). It was stained with a 60% oil red-O solution (in PBS) for 45 minutes and washed with distilled water. The sample was tested under an optical microscope.
Neuron-like cell differentiation
The study was divided into two groups. One group was treat-ed with only growth factors, and the other was processtreat-ed with chemicals. First, BMSCs were cultured to occupy the entire culture medium and separated with a cell scraper. Masses of separated MSCs were then cultured to form 1-2 cluster/cm2. After 24 hours, when the BMSCs attached and
Rock-10 minutes at 55- 60°C.
cDNA synthesis
The total RNA that had been separated was treated with DN-ase. RNA 1 ug was added to Cyclescript RT preMix (dT20, Bioneer Inc.) and cycle in Table 1 was repeated for 15 times to synthesize cDNA.
PCR
Using each synthesized cDNA as a mold, PCR was per-formed with the primers mentioned in Table 2. A total of 30 cycles were repeated.
Electrophoresis
Electrophoresis was performed on the DNA fragments syn-thesized by PCR using 1.5% agarose gel on 1X TAE buffer under 50 V. It was stained with ethidium bromide and ob-served under a U.V. illuminator.
Western blotting
Using a cell scraper, the cells in the medium were separated after being washed twice with PBS. After being collected into a 1.5 mL tube, radioimmunoprecipitation assay(RIPA) buffer [1% triton X-100, 1% sodium deoxycholate, 50 mM NaCl2, 50 mM tris-HCl, 1 mM sodium vanadate, 2 mM phenylmethanesulfonylfluoride (PMSF)] was added to ob-tain lysate. The protein sample was quantified with a protein assay solution, and was next added to 4× sampling buffer. The sample was boiled for five minutes. After electrophore-sis, transfer buffer (25 mM Tris base, 0.2 M glycine, 20% methanol) was used to transfer for one hour under 30V us-ing Immobilon membrane (Millipore). The sample was then treated with blocking buffer (5% skim milk, 1X TBS, 0.1% formaldehyde in PBS, cells were fixed for 4-8 hours and
washed three times with PBS. The sample was then added to blocking solution (5% BSA in PBS) and left to react for 1 hour. Primary antibody was allowed to react for one hour at 4°C. After washing with PBS three times, mounting so-lution was added and covered with a cover glass. The cells were observed under a fluorescent microscope. To test for expression of antigens of neurons, immunofluorescent stain-ing was performed usstain-ing Neuronal Nuclei (NeuN, 1 : 100, Chemicon Inc.), GFAP (1 : 1,000, Chemicon Inc.), MAP-2 (1 : 500, Chemicon Inc.), Neuron Specific Enolase (NSE, 1 : 100, Chemicon Inc.), TH2 (1 : 1,000, Chemicon Inc.), Gal C (1 : 500, Chemicon Inc.) as monoclonal antibodies.
RT-PCR
Total RNA separation
Each cell from 175 mm culture dish was washed twice with PBS. Then, 1 mL of AccuZol (Bioneer Inc., Alameda, CA, USA) was added and cells were harvested into 1.5 mL cen-trifuge tube using a cell scraper. After adding 200 µL of chlo-roform and shaking vigorously for 15 seconds, the mixture was left in ice for five minutes. The mixture was next centri-fuged for 15 minutes at 12,000 rpm at 4°C. After the super-natant was transferred to a 1.5 mL tube, an equal amount of isopropyl alcohol was added, and the mixture was left at -20°C for 10 minutes. At 4°C the mixture was centrifuged again for 15 minutes at 12,000 rpm. The supernatant was re-moved and was washed with 1 mL of 80% ethanol. The so-lution was centrifuged once again for 10 minutes in 12,000 rpm at 4°C, and the supernatant was removed. The ribonu-cleic acid (RNA) pellet was dried at room temperature for 10 to 20 minutes. It was dissolved into DEPC-water and left for
Table 1. cDNA Synthesis
Step Reaction Temperature Time
Step 1 Primer annealing 37°C 30 sec
Step 2 cDNA synthesis 48°C 4 min
Step 3 Melting secondary structure & cDNA synthesis 55°C 30 sec
Table 2. Prime Sequence for PCR
Primer sequence
GFAP for: GTG GGC AGG TGG GAG CTT GAT TCT rev: CTG GGG CGG CCT GGT ATG ACA NSE for: CCC ACT GAT CCT TCC CGA TAC AT rev: CCG ATC TGG TTG ACC TTG AGC A
Map2 for: CCA TTT GCA ACA GGA AGA CAC rev: CAG CTC AAA TGC TTT GCA ACT AT
RESULTS
Marrow collection and MSCs culture
After seven days of culturing marrow derived eukaryocytes in the medium, cell clusters were confirmed under a micro-scope. The attached cells were forming typical mesenchy-mal stem cell shapes (form of neuroglial cells in a pyramid shape with many processes). The primary subculture was completed 15 days later (Fig. 1).
FACS analysis
FACS analysis was performed to confirm bone marrow-de-rived MSCs. Cells from the fourth subculture were collect-ed and testcollect-ed for SH2 and SH4 expressions, which are markers specific to mesenchymal stem cells. The test re-sults were 99.8% positive in cell culture. The test was nega-tive for antibodies (CD45, CD34, CD14), which are specif-ic markers to hematopoietspecif-ic stem cells, in the negative control group. Thus, it was confirmed that the sample of cells that had been grown consisted purely of mesenchymal stem cells, with no hematopoietic cells (Fig. 2).
Differentiation capability test
Alkaline phosphatase and silver nitrate staining of osteoblasts
When replaced with and cultured under high-glucose DMEM [10% fetal bovine serum, 0.1 µM dexamethasone (Sigma), 10 mM β-glycerol phosphate, 50 µM L-ascorbic acid] for three weeks, the stain for alkaline phosphatase and silver nitrate was positive. This confirmed the possibility of mes-enchymal stem cells’ differentiation to osteoblasts (Fig. 3).
Safranin O staininig of chondrogenic cells
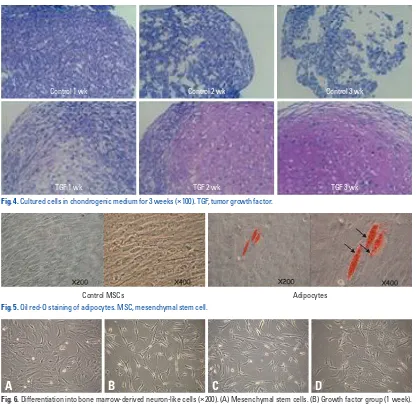
After forming BMSCs into a micromass shape using a cen-trifuge, the cells were cultured in chondrogenic medium for three weeks. As a result of Safranin O stain, differentiation into chondrocytes was observed (Fig. 4).
Oil red-O staining of adipocytes
After culturing with adipogenic induction medium, the re-sult of Oil red-O staining showed and confirmed adipocyte differentiation (red fat vacuoles stained with Oil red-O in cells) (Fig. 5).
Differentiation into neuron-like cells
When cultured with the addition of growth factors or chem-TWEEN 20, polyoxyethylene (20) sorbitan monolaurate) for
[image:6.595.57.283.218.353.2]one hour. At 4°C, antibody [GFAP, NSE, galactocerebroside (Gal C), and NeuN] was allowed to react for 12 hours. Then, washing buffer [1X TBS, 0.1% TWEEN 20, poly-oxyethylene (20) sorbitan monolaurate)] was used three times for five minutes each. Horseradish peroxidase bound secondary antibody was added to the reaction for one hour. It was washed three times with washing buffer. The result was verified with enhanced chemiluminescence (ECL) sys-tem (Habersham Bioscience).
Fig. 1. Mesenchymal stem cell culture (×100). (A) After 7 days of culturing. (B) After 14 days of culturing. (C) Subculture (4 times). (D) Cluster of BMSCs.
A
C
B
D
Fig. 3. Possibility of mesenchymal stem cells’ differentiation to osteoblasts (×100). (A) Alkaline phosphatese staining. (B) Silver nitrate staining.
[image:6.595.59.281.388.596.2]A
B
Fig. 2. Fluorescence Activated Cell Sorting (FACS) analysis.
CD
19
-P
E
CD
4-PE
CD
14
-P
E
CD
4-PE
CD34-FITC CD45-FITC CD3-FITC CD3-FITC
CD34
Co
un
ts
0 20 40 60 80 100
100 101 102 103 104
[image:6.595.56.281.619.712.2]with for two weeks. When the grown and proliferated cells were immunohistochemically tested for neurocyte markers such as NSE, NeuN, and GFAP, the number of positive cells for each stain was very small in the group that was treated with EGF alone. However, when treated with both EGF and HGF, cells that showed positive stains for each staining method were clearly observed. Based on the results mentioned above, the medium treated with both EGF and HGF was much higher in positive immunohistochemical stain ratio than the medium processed with EGF only when investigated regarding the im-munohistochemical staining ratio that was positively stained in each condition (Table 3).
icals, each group showed that, as differentiation progressed, the cells’ processes became thinner and longer, similar to neurocytes, and morphologic changes were observed, such as the area of cytoplasm around the nucleus growing small-er. For the group with growth factor, the shape of neuron-like cells was observable a week later, and two weeks later, most of the cells were similar in shape to neurocytes. Particu-larly, in the group with chemical additions, filament struc-tures of various shapes were seen among the cells (Fig. 6).
Next, to compare and to observe the effects of growth fac-tors used in differentiation into neuron-like cells, EGF alone or a combination of EGF and HGF was given, to be followed up Fig. 5. Oil red-O staining of adipocytes. MSC, mesenchymal stem cell.
Control MSCs Adipocytes
Table 3. Immunohistochemically Tested for Neurocyte Markers
NSE NeuN GFAP
EGF 0.9 % 0.8 % 1.2 %
EGF and HGF 56 % 75 % 24 %
GFAP, glial fibrillary acidic protein; NSE, neuron specific enolase; NeuN, neuronal nuclei; EGF, epidermal growth factor; HGF, hepatocyte growth factor.
Control 1 wk
TGF 1 wk
Control 2 wk
TGF 2 wk
Control 3 wk
TGF 3 wk
[image:7.595.98.514.63.467.2]Fig. 4. Cultured cells in chondrogenic medium for 3 weeks (×100). TGF, tumor growth factor.
Fig. 6. Differentiation into bone marrow-derived neuron-like cells (×200). (A) Mesenchymal stem cells. (B) Growth factor group (1 week). (C) Growth factor group (2 weeks). (D) Chemical group (2 weeks).
godendrocyte was a strong posititve only in the group with growth factors. Through these results, we discovered that MSCs are capable of differentiating into neurocytes and glial cells; in addition, we could also confirm the differenti-ation of part of MSCs populdifferenti-ation into oligodendrocytes (Fig. 8).
RT-PCR
Although all the groups showed expression of the gene in the case of GFAP, the group with growth factor only showed increased expression of the gene. For NSE, the control group was expressed minutely. NSE was much more highly ex-pressed in the group with growth factor only. The group treated with chemicals also showed higher expression than the control group, but less than the group treated with growth factor. It was discovered that MAP2, NF-M, and GAP43 were only expressed in the group with chemical mediators. Based on these results, according to particular differentia-tion condidifferentia-tions, specific markers of some neuron-like cells show differences in expression level (Fig. 9).
Immunostaining
Immunochemical staining
For the group treated with growth factors for two weeks, im-munohistochemical staining of each neurocyte-specific mark-er (NSE, NeuN, and GFAP) was pmark-erformed. It was confirmed that all the markers were positively stained. Namely, it was verified that inducing MSCs differentiation into neuron-like cells by growth factors caused the cells to differentiate most-ly to neurocytes and glial cells (Fig. 7).
Immunofluorescence staining
In the case of immunofluorescent staining in glial fibrillary acidic protein (GFAP), the group treated with growth fac-tors for two weeks showed positive results. In the case of NeuN, the group with growth factors also resulted in posi-tive reactions and even the staining of the control group showed weakly fluorescent results. In the case of Microtu-bule-associated protein 2 (MAP2), the group with growth factors was a strong positive. Gal C that is a marker of
oli-Fig. 8. Immunofluorescence staining (×200). (A and B) GFAP, glial fibrillary acidic protein. (C and D) NeuN, neuronal nuclei. (E and F) MAP2, microtubule-associated protein 2. (G and H) Gal C, galactocerebroside.
B
A
D
C
F
E
H
G
N
eg
at
ive
c
on
tro
l
N
eu
ro
na
l d
iff
er
en
tia
tio
n
(G
ro
w
th
fa
ct
or
s 2
w
ee
[image:8.595.63.496.558.713.2]k)
Fig. 7. Immunochemical staining (×200). (A) NSE, neuron specific enolase. (B) NeuN, neuronal nuclei. (C) GFAP, glial fibrillary acidic protein.
BMSCs using growth factors such as EGF, VEGF, and HGF, we can presume the following three possibilities.
First, there exists a neurocyte subpopulation that can be differentiated into neurocytes in BMSCs that are proliferat-ed and culturproliferat-ed ex vivo. Second, BMSCs can be trans-dif-ferentiated using a method such as mesenchymal epithelial transformation (MET). Finally, as we already know from the results of the RT-PCR and western blotting, under par-ticular conditions of differentiation, it is possible to for cells to differentiate into neuron-like cells, each with different characteristics.
Bossolasco, et al.44 postulated that, since nestin and beta
tubulin III, together with O4 and GFAP showed a positive reaction, MSCs already have a subpopulation that has a ca-pability to differentiate into neuron-like cells. In addition, Ji-ang, et al.45 reported that selectively proliferated cells can be
differentiated into multipotent precursor cells with the char-acteristic shapes of neurocytes, astrocytes, and oligodendro-cytes. Chamberlain, et al.46 also proved the possible
trans-differentiation of mesenchymal stem cells.
Western blotting
With the exception of the control group, all the groups in-duced differentiation showed expression of the protein GFAP. The group with growth factor showed markedly higher ex-pression of protein than the group with chemicals. For NSE, the entire group showed expression of the protein, and the expression was particularly higher in the group with growth factor. In the Gal C test, the control group re-sults were weakly positive for Gal C expression, and higher than the expression found in the group with chemicals. NeuN was expressed in high quantity in both groups with growth factor and with chemicals. Based on these results, according to particular differentiation conditions, specific markers of some neuron-like cells show a difference in the level of expression (Fig. 10).
DISCUSSION
Through the results of neuron-like cell differentiation from
GFAP
NSE
Gal C
NeuN
[image:9.595.124.515.365.539.2]Control group Growth factors 1 week group Growth factors 2 week group Chemical group
Fig. 10. Western blotting. GFAP, glial fibrillary acidic protein; NSE, neuron specific enolase; NeuN, neuronal nuclei; Gal C, galactocerebroside.
GFAP
NSE
MAP2
NF-M
GAP 43
[image:9.595.123.514.566.722.2]Control group Chemical group Growth factors 1 week group Growth factors 2 week group
mal cerebral infarct models, symptoms has resulted in re-covery and decreased neurological deficit when injected with MSCs.8,61-63 It has also been discovered that when
BM-SCs are injected into blood vessels, rather than directly into damaged brain tissue, the cells have shown an ability to pass through the blood-brain barrier and to migrate to the damaged part of the brain.2,20,21,61,64 In a clinical study, Bang,
et al.65 proliferated and introduced autologous MSCs
intra-venously to patients with cerebral infarction. They reported that injecting these stem cells had the effect of improving neurological symptoms, such as reducing the size of the in-farct and decreasing the atrophy of the ventricle.66
In this study, BMSCs’ ability to proliferate and differenti-ate into neuron-like cells by has indicdifferenti-ated a possibility for application to cell therapy for a wide range of diseases such as neurodegenerative disease and cerebral infarct. Previous-ly known methods of inducing neuron-like cell differentia-tion using several chemicals have limits because the toxici-ty of these chemicals makes them too risky to be applied in humans. However, the EGF, VEGF, and HGF used in this study are secreted in human body, and can be safely manip-ulated to induce differentiation. Furthermore, the use of dif-ferentiated neuron-like cells in clinical therapeutic methods may have fewer limitations. These cells also have autolo-gous marrow origin, and are thus free from severe compli-cations; for example, immune rejection reaction. Because large quantities of neuron-like cells can be acquired from a small amount of bone marrow, these cells’ application in clinical therapy will be very useful.
In the future, it will be necessary to compare and analyze the characteristics of neuron-like cells originating from MSCs with other neurons in molecular biological and phys-iological aspects. Further studies are required to uncover the possibilities of transplanting them into humans and differen-tiating the cells within animal.
REFERENCES
1. Owen M. Lineage of osteogenic cells and their relationship to the stromal system. In: Peck WA, editor. Bone and mineral research. Amsterdam: Elsevier; 1985. p.1-25.
2. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, et al. Multilineage potential of adult human mesenchy-mal stem cells. Science 1999;284:143-7.
3. Tremain N, Korkko J, Ibberson D, Kopen GC, DiGirolamo C, Phinney DG. MicroSAGE analysis of 2,353 expressed genes in a single cell-derived colony of undifferentiated human mesenchy-mal stem cells reveals mRNAs of multiple cell lineages. Stem
When BMSCs proliferate and differentiate, as we discov-ered in the immunohistochemical stain positive ratio of the two groups, one treated with EGF only and the other with both EGF and HGF, it is our understanding that EGF stim-ulates cell proliferation and HGF stimstim-ulates differentiation into neuron-like cells. EGF along with bFGF is used as a factor to grow and proliferate neural stem cells or neural precursor cells derived from mouse or human brain tissue. Because previous reports already discovered that BMSCs express receptors for EGF,47 EGF in this study is also thought
to be associated with the proliferation and differentiation of BMSCs into neuron-like cells.22-26 HGF has a pleiotropic
function and plays an important role in the organogenesis of various epithelial cells30 including renal, pulmonary,
gas-tric, bowel mucosa, corneal, and skin epithelium and tissue regeneration.48-50 It also plays an important role in growth
and differentiation of stromal cells such as osteoblastocytes and myocytes.51-53 HGF is a rare neurotrophic factor that is
expressed all over the brain tissue.54-57 HGF improves the
survivability of neurons in the hippocampus and midbrain, and induces the growth of neurite in neocortical explants.54,57
In the peripheral nervous system, HGF functions as a sur-vival factor of motor neurons. Particularly, in the develop-ment process, it functions as a axonal chemoattractant to spinal motor neuron,58 and is also associated with the growth
and survival of sensory and parasympathetic neurons.59
Currently, surgical and medical treatments for central nervous system diseases, including degenerative, traumatic, and ischemic damage, are very limited, and it is difficult to expect recovery from nervous system damage. To regener-ate the damaged nervous tissues, a method called neuron transplantation is a raised possibility. A study that transplant-ed fetal dopaminergic neurons to an adult who was suffering from Parkinson’s disease showed positive results.34
Howev-er, its use is controversial because there are still many ethi-cal problems with the application of fetal or embryo stem cells to patient treatment. Also, techniques for obtaining a sufficient quantity of adult neuronal stem cells to (NSCs) are still limited.29,32 Therefore, this cannot be considered a
proper method for application in clinical circumstances.60
However, MSCs have excellent proliferation capabilities that can overcome the adult neuronal stem cells’ limitation of quantity. It has also been confirmed that MSCs are capa-ble of differentiating into cells that have similar characteris-tics to neurogenic cells, and it is possible for these cells to be used in the recovery of damaged neurons.
ani-striatal embryonic progenitor cell produces neurons and astro-cytes. J Neurosci 1992;12:4565-74.
23. Reynolds BA, Weiss S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Sci-ence 1992;255:1707-10.
24. Reynolds BA, Weiss S. Clonal and population analyses demon-strate that an EGF-responsive mammalian embryonic CNS pre-cursor is a stem cell. Dev Biol 1996;175:1-13.
25. Richards LJ, Kilpatrick TJ, Bartlett PF. De novo generation of neuronal cells from the adult mouse brain. Proc Natl Acad Sci U S A 1992;89:8591-5.
26. Carpenter MK, Cui X, Hu ZY, Jackson J, Sherman S, Seiger A, et al. In vitro expansion of a multipotent population of human neural progenitor cells. Exp Neurol 1999;158:265-78.
27. Nakamura T, Nishizawa T, Hagiya M, Seki T, Shimonishi M, Sugimura A, et al. Molecular cloning and expression of human hepatocyte growth factor. Nature 1989;342:440-3.
28. Matsumoto K, Nakamura T. Hepatocyte growth factor (HGF) as a tissue organizer for organogenesis and regeneration. Biochem Biophys Res Commun 1997;239:639-44.
29. Gage FH. Mammalian neural stem cells. Science 2000;287:1433-8. 30. Björklund A, Lindvall O. Self-repair in the brain. Nature 2000;
405:892-3, 5.
31. Rakic P. Adult neurogenesis in mammals: an identity crisis. J Neu-rosci 2002;22:614-8.
32. Temple S, Alvarez-Buylla A. Stem cells in the adult mammalian central nervous system. Curr Opin Neurobiol 1999;9:135-41. 33. Bain G, Kitchens D, Yao M, Huettner JE, Gottlieb DI. Embryonic
stem cells express neuronal properties in vitro. Dev Biol 1995;168: 342-57.
34. Freed CR, Greene PE, Breeze RE, Tsai WY, DuMouchel W, Kao R, et al. Transplantation of embryonic dopamine neurons for se-vere Parkinson’s disease. N Engl J Med 2001;344:710-9. 35. Lindvall O, Brundin P, Widner H, Rehncrona S, Gustavii B,
Frackowiak R, et al. Grafts of fetal dopamine neurons survive and improve motor function in Parkinson’s disease. Science 1990;247: 574-7.
36. McKay R. Stem cells in the central nervous system. Science 1997; 276:66-71.
37. Hüttmann A, Li CL, Dührsen U. Bone marrow-derived stem cells and “plasticity”. Ann Hematol 2003;82:599-604.
38. Isacson O, Bjorklund LM, Schumacher JM. Toward full restora-tion of synaptic and terminal funcrestora-tion of the dopaminergic system in Parkinson’s disease by stem cells. Ann Neurol 2003;53 Suppl 3:S135-46.
39. Silani V, Cova L, Corbo M, Ciammola A, Polli E. Stem-cell thera-py for amyotrophic lateral sclerosis. Lancet 2004;364:200-2. 40. Clement AM, Nguyen MD, Roberts EA, Garcia ML, Boillée S,
Rule M, et al. Wild-type nonneuronal cells extend survival of SOD1 mutant motor neurons in ALS mice. Science 2003;302:113-7. 41. Holden C, Vogel G. Stem cells. Plasticity: time for a reappraisal?
Science 2002;296:2126-9.
42. Mejia-Aranguré JM, Fajardo-Gutiérrez A, Flores-Aguilar H, Mar-tinez-García MC, Salamanca-Gómez F, Palma-Padilla V, et al. Environmental factors contributing to the development of child-hood leukemia in children with Down’s syndrome. Leukemia 2003;17:1905-7.
43. Svendsen CN, Langston JW. Stem cells for Parkinson disease and ALS: replacement or protection? Nat Med 2004;10:224-5. 44. Bossolasco P, Cova L, Calzarossa C, Rimoldi SG, Borsotti C, Cells 2001;19:408-18.
4. Le Blanc K, Pittenger M. Mesenchymal stem cells: progress to-ward promise. Cytotherapy 2005;7:36-45.
5. Lee RH, Kim B, Choi I, Kim H, Choi HS, Suh K, et al. Character-ization and expression analysis of mesenchymal stem cells from human bone marrow and adipose tissue. Cell Physiol Biochem 2004;14:311-24.
6. Wang G, Bunnell BA, Painter RG, Quiniones BC, Tom S, Lanson NA Jr, et al. Adult stem cells from bone marrow stroma differenti-ate into airway epithelial cells: potential therapy for cystic fibrosis. Proc Natl Acad Sci U S A 2005;102:186-91.
7. Brazelton TR, Rossi FM, Keshet GI, Blau HM. From marrow to brain: expression of neuronal phenotypes in adult mice. Science 2000;290:1775-9.
8. Mezey E, Chandross KJ, Harta G, Maki RA, McKercher SR. Turning blood into brain: cells bearing neuronal antigens generat-ed in vivo from bone marrow. Science 2000;290:1779-82. 9. Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, et al.
Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng 2001;7:211-28.
10. Katz AJ, Tholpady A, Tholpady SS, Shang H, Ogle RC. Cell sur-face and transcriptional characterization of human adipose-derived adherent stromal (hADAS) cells. Stem Cells 2005;23:412-23. 11. Panepucci RA, Siufi JL, Silva WA Jr, Proto-Siquiera R, Neder L,
Orellana M, et al. Comparison of gene expression of umbilical cord vein and bone marrow-derived mesenchymal stem cells. Stem Cells 2004;22:1263-78.
12. In ‘t Anker PS, Scherjon SA, Kleijburg-van der Keur C, de Groot-Swings GM, Claas FH, Fibbe WE, et al. Isolation of mesenchy-mal stem cells of fetal or maternal origin from human placenta. Stem Cells 2004;22:1338-45.
13. Campagnoli C, Roberts IA, Kumar S, Bennett PR, Bellantuono I, Fisk NM. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood 2001;98:2396-402.
14. Kuznetsov SA, Friedenstein AJ, Robey PG. Factors required for bone marrow stromal fibroblast colony formation in vitro. Br J Haematol 1997;97:561-70.
15. van den Bos C, Mosca JD, Winkles J, Kerrigan L, Burgess WH, Marshak DR. Human mesenchymal stem cells respond to fibro-blast growth factors. Hum Cell 1997;10:45-50.
16. Morse WR. Chinese Medicine. New York: Hoeber; 1938. 17. Eglitis MA, Mezey E. Hematopoietic cells differentiate into both
microglia and macroglia in the brains of adult mice. Proc Natl Acad Sci U S A 1997;94:4080-5.
18. Azizi SA, Stokes D, Augelli BJ, DiGirolamo C, Prockop DJ. En-graftment and migration of human bone marrow stromal cells im-planted in the brains of albino rats--similarities to astrocyte grafts. Proc Natl Acad Sci U S A 1998;95:3908-13.
19. Kopen GC, Prockop DJ, Phinney DG. Marrow stromal cells mi-grate throughout forebrain and cerebellum, and they differentiate into astrocytes after injection into neonatal mouse brains. Proc Natl Acad Sci U S A 1999;96:10711-6.
20. Sanchez-Ramos J, Song S, Cardozo-Pelaez F, Hazzi C, Stedeford T, Willing A, et al. Adult bone marrow stromal cells differentiate into neural cells in vitro. Exp Neurol 2000;164:247-56.
21. Woodbury D, Schwarz EJ, Prockop DJ, Black IB. Adult rat and human bone marrow stromal cells differentiate into neurons. J Neurosci Res 2000;61:364-70.
Brain Res Mol Brain Res 1995;32:197-210.
56. Yamagata T, Muroya K, Mukasa T, Igarashi H, Momoi M, Tsuka-hara T, et al. Hepatocyte growth factor specifically expressed in mi-croglia activated Ras in the neurons, similar to the action of neuro-trophic factors. Biochem Biophys Res Commun 1995;210:231-7. 57. Hamanoue M, Takemoto N, Matsumoto K, Nakamura T,
Nakaji-ma K, Kohsaka S. Neurotrophic effect of hepatocyte growth factor on central nervous system neurons in vitro. J Neurosci Res 1996; 43:554-64.
58. Ebens A, Brose K, Leonardo ED, Hanson MG Jr, Bladt F, Birch-meier C, et al. Hepatocyte growth factor/scatter factor is an axonal chemoattractant and a neurotrophic factor for spinal motor neu-rons. Neuron 1996;17:1157-72.
59. Davey F, Hilton M, Davies AM. Cooperation between HGF and CNTF in promoting the survival and growth of sensory and para-sympathetic neurons. Mol Cell Neurosci 2000;15:79-87.
60. Aboody KS, Brown A, Rainov NG, Bower KA, Liu S, Yang W, et al. Neural stem cells display extensive tropism for pathology in adult brain: evidence from intracranial gliomas. Proc Natl Acad Sci U S A 2000;97:12846-51.
61. Zhao LR, Duan WM, Reyes M, Keene CD, Verfaillie CM, Low WC. Human bone marrow stem cells exhibit neural phenotypes and ameliorate neurological deficits after grafting into the isch-emic brain of rats. Exp Neurol 2002;174:11-20.
62. Chen J, Li Y, Katakowski M, Chen X, Wang L, Lu D, et al. Intra-venous bone marrow stromal cell therapy reduces apoptosis and promotes endogenous cell proliferation after stroke in female rat. J Neurosci Res 2003;73:778-86.
63. Chen J, Li Y, Wang L, Lu M, Zhang X, Chopp M. Therapeutic benefit of intracerebral transplantation of bone marrow stromal cells after cerebral ischemia in rats. J Neurol Sci 2001;189:49-57. 64. Li Y, Chen J, Chen XG, Wang L, Gautam SC, Xu YX, et al.
Hu-man marrow stromal cell therapy for stroke in rat: neurotrophins and functional recovery. Neurology 2002;59:514-23.
65. Bang OY, Lee JS, Lee PH, Lee G. Autologous mesenchymal stem cell transplantation in stroke patients. Ann Neurol 2005;57:874-82. 66. Nagai A, Kim WK, Lee HJ, Jeong HS, Kim KS, Hong SH, et al.
Multilineage potential of stable human mesenchymal stem cell line derived from fetal marrow. PLoS One 2007;2:e1272. Deliliers GL, et al. Neuro-glial differentiation of human bone
mar-row stem cells in vitro. Exp Neurol 2005;193:312-25.
45. Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002;418:41-9.
46. Chamberlain JR, Schwarze U, Wang PR, Hirata RK, Hankenson KD, Pace JM, et al. Gene targeting in stem cells from individuals with osteogenesis imperfecta. Science 2004;303:1198-201. 47. Deans RJ, Moseley AB. Mesenchymal stem cells: biology and
potential clinical uses. Exp Hematol 2000;28:875-84.
48. Noji S, Tashiro K, Koyama E, Nohno T, Ohyama K, Taniguchi S, et al. Expression of hepatocyte growth factor gene in endothelial and Kupffer cells of damaged rat livers, as revealed by in situ hy-bridization. Biochem Biophys Res Commun 1990;173:42-7. 49. Defrances MC, Wolf HK, Michalopoulos GK, Zarnegar R. The
presence of hepatocyte growth factor in the developing rat. Devel-opment 1992;116:387-95.
50. Sonnenberg E, Meyer D, Weidner KM, Birchmeier C. Scatter fac-tor/hepatocyte growth factor and its receptor, the c-met tyrosine ki-nase, can mediate a signal exchange between mesenchyme and epi-thelia during mouse development. J Cell Biol 1993;123:223-35. 51. Sato T, Hakeda Y, Yamaguchi Y, Mano H, Tezuka K, Matsumoto
K, et al. Hepatocyte growth factor is involved in formation of os-teoclast-like cells mediated by clonal stromal cells (MC3T3-G2/ PA6). J Cell Physiol 1995;164:197-204.
52. Grano M, Galimi F, Zambonin G, Colucci S, Cottone E, Zallone AZ, et al. Hepatocyte growth factor is a coupling factor for osteo-clasts and osteoblasts in vitro. Proc Natl Acad Sci U S A 1996;93: 7644-8.
53. Allen RE, Sheehan SM, Taylor RG, Kendall TL, Rice GM. Hepa-tocyte growth factor activates quiescent skeletal muscle satellite cells in vitro. J Cell Physiol 1995;165:307-12.
54. Jung W, Castren E, Odenthal M, Vande Woude GF, Ishii T, Dienes HP, et al. Expression and functional interaction of hepatocyte growth factor-scatter factor and its receptor c-met in mammalian brain. J Cell Biol 1994;126:485-94.