Bone Morphogenetic Protein Receptor in the Osteogenic Differentiation of Rat Bone Marrow Stromal Cells
6
0
0
Full text
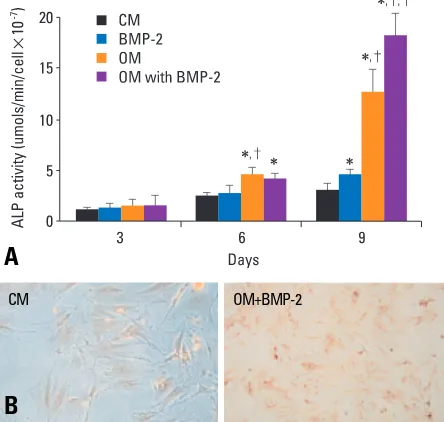
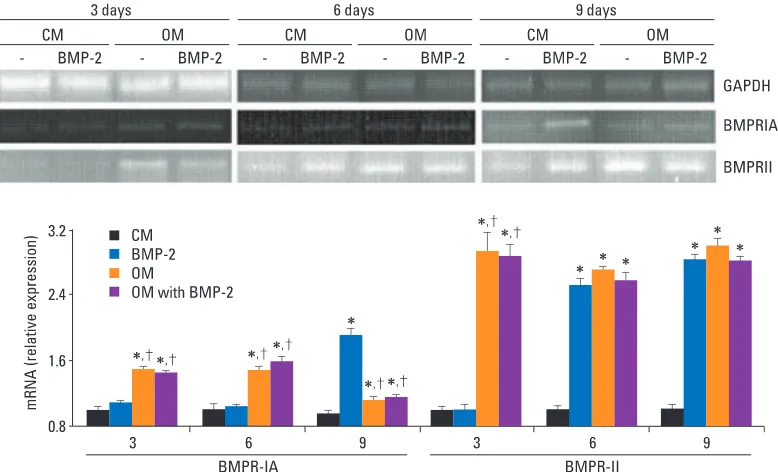
Figure
Related documents