J. Exp. Biol. (i97l)» 54. 737-744 7 3 7 With 6 text-figures
Printed in Great Britain
THE VISUAL MECHANISMS OF
TENEBRIO MOLITOR: CHANGES IN THE
ELECTRO-RETINOGRAM AS FUNCTION OF THE
STIMULUS DURATION
BY U. YINON*
The Vison Research Laboratory, Hadassah University Hospital, Jerusalem, Israel
{Received 27 October 1970)
INTRODUCTION
The yellow mealworm beetle Tenebrio molitor represents a group of insects living almost constantly under conditions of darkness. Some evolutionary changes pre-sumably occurred in the visual mechanism of this species during or after the adoption of its specific habitat. Various aspects of its visual mechanism have already been investigated (Yinon, 1969, 1970a; Yinon and Auerbach, 1969). In the present studies another aspect of the visual mechanism - the response as function of stimulus dura-tion - has been investigated. This has been compared with the response in diurnal or nocturnal insects not living in a permanently dark habitat.
The insect electroretinogram (ERG) is a complex of potential changes. Hartline (1928) partially explained the origin of the various components by demonstrating that the negative component of the insect ERG comes from the retina, while the positive component arises from the optic ganglion. The relative prominence of these two components varies with the anatomy of the eye. Therefore, in comparison to the vertebrate's ERG (Davson, 1962), there is less certainty about the insect's ERG pattern (Wigglesworth, 1965; Yinon, 1970 J). In the present studies the origin of various components of the ERG have been studied as well as their relative contribu-tion in the context of the duracontribu-tion and intensity of the stimulus.
METHODS
Results were calculated for 23 adults of the yellow mealworm beetle T. molitor (L.). The animals were dark-adapted for 5-10 min before each experiment. Between stimuli of different durations at each intensity, intervals of one minute were allowed for recovery in the dark, and between series of different intensities the intervals were of some minutes. Only one intensity level (log / = o-o) was used when exposures of longer than 1 s were given and the intervals were of 2-3 min between the stimuli.
The optical system was described previously (Yinon, 1970c). Exposures from 1 ms to is were given by Wollensak shutter and from 1 ms to 14 s by Exacta shutter. The signals from these shutters were displayed on the oscilloscope for each ERG. Relative
1-0 ms 8250 ms
100 ms
400 ms
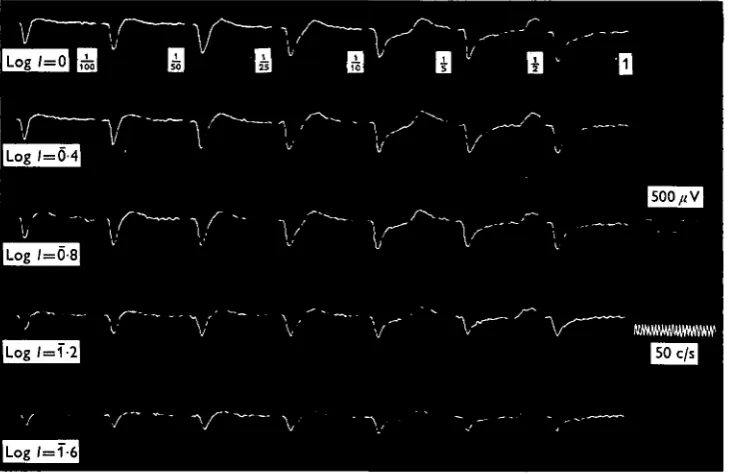
Fig. i. Changes in the ERG for a wide range of stimulus durations at a given intensity (log I = o-o). Stimulus durations for Group I are seen above the first column of calibration marks and for Group II above the second one. The left column of responses in Group II shows the 'on' potential of the same responses as the right column but on a different time scale.
Visual mechanisms of Tenebrio molitor 739
light intensity was determined by Ilford neutral circular wedge (3-0 log units) and ^ r Kodak optical density filters. Schott heat filter was used to exclude infrared radia-tion. The maximum intensity (log / = 00) was 70 lux over the eye.
The animal was connected through a pre-amplifier to the recording system which was synchronized with the shutter oscilloscope and the camera. The spontaneous activity was monitored by an additional oscilloscope. In some of the experiments an averaging device (computer of Average Transients, 400 B, TMC), connected to the electronic counter was used and was triggered by a photo-cell placed near the eye.
For other experimental conditions see Yinon and Auerbach (1969) and Yinon (1970A).
RESULTS
[image:3.451.46.411.264.501.2]The ERG pattern for short stimulus durations (less than 100 msec) is composed of a fast negative potential which is followed by a slow positive one. In longer durations a faster positive component appeared coupled with the end of the stimulus, thus called
Fig. 2. The combined effect of stimulus duration and intensity on the ERG.
740
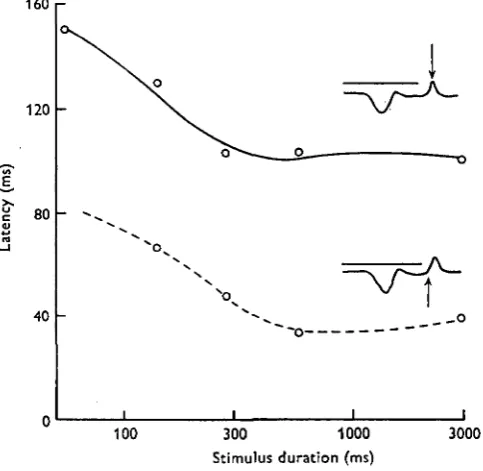
For a given intensity and above a certain threshold of stimulus duration, the exposure time and the latent period of the 'on' are independent. It is not dead however, why the latency and the peak latency (the period from the beginning of th? stimulus to the appearance of the response peak) are still constant in their length in
10000
1000
100
10
10 100 1000 Stimulus duration (ms)
[image:4.451.80.377.129.356.2]10000
Fig. 3. The latent periods of the ERG components as functions of stimulus duration (I = ' off' II = 'on'-peak, A = 'on').
160
120
80
40
100 300 1000 Stimulus duration (ms)
3000
[image:4.451.104.346.390.625.2]Visual mechanisms of Tenebrio molitor
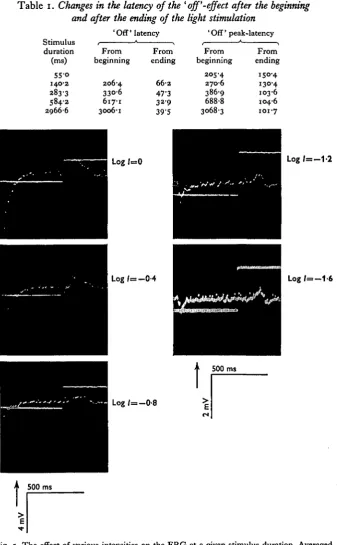
Table i. Changes in the latency of the ' off'-effect after the beginning
and after the ending of the light stimulation ' Off' latency ' Off' peak-latency S t i m u l u s i * > i * >
duration From From From From (ms) beginning ending beginning ending
55-o
140-2
283-3 584-2 2966-6
206-4 330-6 617-1 3006-1
6 6 2
47-3 32-9 39-5
205-4 270-6
386-9 688-8 3068-3
150-4 130-4 103-6 104-6 101-7
Log / = 0
L o g / = - 0 - 4
Log 1= - 0 - 8
L o g / = - 1 - 2
Log /=—1-6
[image:5.451.54.392.78.624.2]500 ms
742
[image:6.451.47.403.163.442.2]presence of subthreshold stimuli (Fig. 3). The latent period of the 'off' is clearly related to the stimulus extinction, yielding a linear ratio (as function of duration) a* stimulus durations longer than 300 msec (Fig. 3). But in short stimulus durations tlH flexibility in the length of the 'off' latency is higher since its appearance is not tightly coupled with the end of the stimulus. The 'off' latency decreased - after stimulus
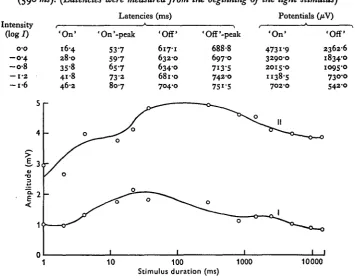
Table 2. Changes in the ERG as function of the light intensity at a given exposure time (590 ms). (Latencies were measured from the beginning of the light stimulus)
Intensity (log/)
o-o - o - 4 - o - 8
— 1-a - i - 6
' O n ' 16-4
2 8 0
358 41-8 46-2
Latencies
'On'-peak
5 3 7 5 9 7
6S-7
7 3 2 8 0 7
(ms) 'Off' 617-1 632-0 6340 6810 704-0 'Off'-peak 688-8 6970 7I3-S 742-0 75i-5 Potentials (/iv)
' O n ' 4731-9 32900 2015-0 1138-5 702-0 'Off' 2362-6 18340 1095-0 730-0 542-0 %
1 10 100 1000 10000 Stimulus duration (ms)
Fig. 6. The ERG amplitude as function of the stimulus duration (I = 'off', II = 'on'). extinction - when the stimulus duration increased up to values of 500 msec (Fig. 4, Table 1). In exposures longer than this the 'off' latencies are not more influenced by the addition of light energy to the stimulus. Thus it reacts differently than the 'on' under the same conditions. The difference between the latent period (70 msec, log / = o-o) and the peak latency (150 msec) of the 'off' (after extinction of the stimulus) is constant.
Visual mechanisms of Tenebrio molitor 743
The stimulus duration was found to determine the ERG magnitude to a given intensity as expected from Bunsen-Roscoe law. A similar trend is seen for both ' on' Ihd 'off' effects, although in the latter the response level is lower (Fig. 6). This and the similarity shown by the intensity functions presumably indicate a common origin in the receptor cells. When the set of the curves received for different stimulus durations under five intensity levels (from log / = o-o to l o g / = —1'6) were compared (for 'on' effect) equal amplitudes were received in combination of stimuli ranged from 10-40 msec duration and intensities between l o g / = -0-4 to l o g / = -1-2. At higher intensities the response amplitude was found to depend on the light intensity only. (Insufficient data were obtained to make it possible to calculate / x t [I = intensity, t = duration]) constants.
DISCUSSION
Autrum (1958) has classified compound eyes of beetles among the 'slow type' giving negative monophasic electrical response. In contrast, T. molitor which belongs taxonomically to this group has a diphasic ERG including also an 'off' component. The ERG of the dragonfly Aeshva cyanea consists of a retinal and a neural component. The latter ('off' effect) is contributed by the optic ganglia (Autrum, 1958). The positive 'off' response of Tenebrio molitor seems to originate in the receptor zone as well as the' on' response. By sequential and gradual increase of the stimulus duration it was possible to separate the ERG of T. molitor into 'on' and 'off' potentials (in 100 ms). Among previous studies reviewed it was found to be in 330 ms for the stemmata of Bombyx mori (Ishikawa & Hirao, i960).
An increase of amplitude as function of stimulus duration has been observed in some species. It was found in the ERG of the cricket Tachycines asynamorus by Autrum (1948a) and for the 'off' potential (the 'on' remained constant) of the fly
Calliphora erythrocephala (Autrum, 19486). A peak occurred for the euphausiid Euphausia pacifica at a stimulus duration of 100 ms (Boden & Kampa, 1965) (as in
Tenebrio molitor) and at 430 ms for the crayfish Procambarus clarkii (Naka &
Kuwabara, 1959). A constant response may be obtained with different intensities of illumination provided that the duration of the stimulus is varied in such a way that the product of intensity and duration is constant (Ixt = const., Bunsen-Roscoe Law of photochemistry). In insects this relation was found to hold for exposures of less than 80 ms (Harline, 1928) whereas in man it is between 2 and 100 ms (LeGrand, 1957). It was found to be between 10 and 40 ms in the present investigation. Exposures longer than this yield a maximum of the principal potential, which depends on the intensity alone. According to the present studies the amplitude is diminished at a given intensity level in durations above 300 ms for a reason which is still unknown.
In a previous study a way was found to calculate the spectral response of T. molitor according to the latency ('on') function which is approximately a reciprocal of the amplitude (Yinon, 1969). In the present studies a relation was not found between the amplitude and the latency in relation to the stimulus duration. Thus the 'on' latency is no longer a function of the stimulus duration, even in the small range in which the amplitude is.
SUMMARY
1. The ERG complex of the compound eye of the yellow mealworm bettle TenebrH
molitor (L.) separates into two components with stimulus durations of longer than
ioo ms. These are the 'on' and the 'off' effects. Above stimulus durations of i s the positive potential following the 'on' is faster and the positive 'off' is followed by a small negative one.
2. The latent period of the ' on' is independent of the stimulus duration while for stimulus durations of longer than 300 ms the 'off' latency is coupled with the end of the stimulus. If measured after the extinction of the stimulus the 'off' latency is affected by the stimulus duration.
3. The 'on' and 'off' amplitudes behave similarly for various stimulus durations. Both originate in the receptor cells.
4. The Bunsen-Roscoe Law of photochemistry holds for stimulus durations of 10-40 ms. For a given intensity, the amplitudes diminished for durations longer than 300 ms.
I am indebted to Professor E. Auerbach, Vision Research Laboratory, Hadassah University Hospital, Jerusalem, Israel, in whose laboratory this work was conducted, and also to Professor A. Shulov, Department of Entomology, the Hebrew University, who provided financial support.
REFERENCES
AUTRUM, H. (1948a). Zur Analyse des zeitlichen Auflosungsvermogens des Insektenauges. Nachr. Ges.
Wiss. Gottingen, Math.-physik kl. 13-18.
AUTRUM, H. (19486). Uber das zeitliche Auflosungsvermogen des Insektenauges. Nachr. Ges. Wiss.
Gottingen, Math.-physik. kl. Biol.-Phys. Abt. 8-13.
AUTRUM, H. (1958). Electrophysiological analysis of the visual systems in insects. Exp. Cell. Res.
(Suppl.) 5, 426-39.
BODEN, B. P. & KAMPA, E. M. (1965). An aspect of euphasiid ecology revealed by echo-sounding in a fjord. Crustaceana 9, 155-73.
DAVSON, H. (1962). The Eye, vol. 2. New York and London: Academic Press.
GOLDSMITH, T. H. (1963). The course of light and dark adaptation in the compound eye of the honey-bee. Camp. Biochem. Physiol. 10, 227-37.
HARTLINE, H. K. (1928). A quantitative and descriptive study of the electric response to illumination of the arthropod eye. Am. J. Physiol. 83, 466-83.
ISHIKAWA, S. & HIRAO, T. (i960). Electrophysiological studies on vision of the silkworm Bombyx mori I: Electroretinogram of stemmata. J. Seric. Set. Japan 29, 8-14.
LEGRAND, Y. (1957). Light, Colour and Vision. London: Chapman and Hall Ltd. MAZOKHIN-PORSHNYAKOV, G. A. (1969). Insect Vision. New York: Plenum Press.
NAKA, K. & KUWABARA, M. (1959). Two components of the compound eye of the crayfish. J. Exp. Biol. 36, 51-61.
YINON, U. (1969). Spectral efficiency as function of latency in the visual mechanism of insect {Tenebrio
molitor L.). Experientia 25, 711.
YINON, U. & AUERBACH, E. (1969). The visual mechanisms of Tenebrio molitor: Variations taking place in the ERG of pupa and adult during development. J. Exp. Biol. 51, 635-41.
YINON, U. (1970a). The visual mechanisms of Tenebrio molitor: Some aspects of the spectral response.
J. Exp. Biol. 53, 221-9.
YINON, U. (19706). Similarity of the electroretinogram in insects. J. Insect Physiol. 16, 221-5. YINON, U. (1970c). The electrophysiological and behavioral response of a stored product beetle to
light. J. Stored Prod. Res. 6, 195-8.
YINON, U. (i97O<f). Electrophysiological evidence of relationships in Acridid insects. J. System. Zool. 19. 397-8.