5-Hydroxytryptamine (5-HT) has been proposed as a neurotransmitter in crustaceans on several grounds. Levels have been measured in the nervous system of various crustacean species (Livingstone et al. 1981; Elofsson et al. 1982; Laxmyr, 1984; Kulkarni and Fingerman, 1992). A number of immunopositive neurones have been identified in central ganglia using anti-5-HT antisera (Eloffson, 1983; Beltz and Kravitz, 1983; Sandeman et al. 1988; Bellon-Humbert and Van Herp, 1988). 5-HT has also been measured in the
haemolymph (Livingstone et al. 1980; Elofsson et al. 1982), thus suggesting a possible role as a neurohormone.
Various effects have been reported for 5-HT that implicate it as a neuroactive agent in crustaceans. Its injection into the circulation is known to induce a behavioural pattern of flexion in lobster and crayfish (Livingstone et al. 1980), and neuronal targets for this action have been identified in the thoracic and abdominal ganglia (Kravitz, 1988; Ma et al. 1992). A modulatory role on the crayfish escape response has also been JEB0864
The content and regional distribution of 5-hydroxytryptamine (5-HT) in the crayfish eyestalk was determined by high-performance liquid chromatography. Levels of the 5-HT precursors L-tryptophan (L-TRP) and
5-hydroxytryptophan (5-OH-TRP), and of three metabolites, 5-hydroxytryptophol (5-HTPH), N-acetylserotonin (NA-5-HT) and 5-hydroxy-indole-3-acetic acid (5-HIAA), were also determined. The total content of 5-HT in the eyestalk was 95.4±49.3 pg mg−1wet mass (mean ± S.D., N=55) while the
specific content was 9.6±4.9 fmolµg−1protein (mean ±S.D. N=5). 5-HT was present in all four ganglia of the eyestalk.
The highest proportion was found in the medulla terminalis (40.2 %) and the lowest in the retina lamina ganglionaris (9.9 %), which also had the lowest specific content. Conversely, the highest specific content of L-TRP was in the retina lamina ganglionaris.
5-HT biosynthesis and metabolism were explored in isolated eyestalks. The monoamine oxidase blocker pargyline, at concentrations between 0.8 and 10 mmol l−1, elicited a
dose-dependent increase in HT content. The biosynthesis of 5-HT in the crayfish eyestalk is suggested by the presence of its immediate precursor (5-OH-TRP) and by the suppression of 5-HT synthesis induced by m-hydroxybenzyl-hydrazine (m-HBH), a blocker of 5-OH-TRP decarboxylase.
The presence of immunopositive cell bodies and axons was demonstrated using an anti-5-HT antiserum. 5-HT-like immunopositivity was detected in various regions of the eyestalk. Efferent immunopositive axons were also identified in the optic nerve, and these may have originated in the protocerebral lobe of the supraoesophageal ganglion. The branchings of these axons were profusely distributed in the neuropile of the medulla terminalis.
A basal level release of 5-HT was detected in isolated eyestalks. The amount recovered was increased two-to threefold after blocking 5-HT uptake with fluoxetine (1µmol l−1). Incubation of eyestalks in solutions containing
a high K+ concentration (80 mmol l−1) released 5-HT.
Electrical stimulation of the optic nerve released 5-HT as a function of the intensity of stimulation. Both the basal and evoked release were suppressed by lowering the Ca2+
concentration in the medium.
These observations support a role for 5-HT as a neurotransmitter or neuromodulator in the crayfish eyestalk.
Key words: serotonin, neurosecretion, crayfish, Procambarus clarkii, crustacean.
Summary
Introduction
LOCALIZATION AND RELEASE OF 5-HYDROXYTRYPTAMINE IN THE CRAYFISH
EYESTALK
LEONARDO RODRÍGUEZ-SOSA1, ARTURO PICONES2,*, GABINA CALDERÓN ROSETE1,
SOCORRO ISLAS2 ANDHUGO ARÉCHIGA1,†
1División de Estudios de Posgrado e Investigación, Facultad de Medicina, Universidad Nacional Autónoma de
México, México DF and 2Departamento de Fisiología, Biofísica y Neurociencias, Centro de Investigación y de Estudios Avanzados, IPN, México DF
Accepted 14 August 1997
proposed (Glanzman and Krasne, 1986; Yeh et al. 1996). 5-HT is known to exert a facilitatory effect on lobster and crayfish neuromuscular transmission (Dudel, 1965; Glusman and Kravitz, 1982; Fischer and Florey, 1983; Dixon and Atwood, 1985) and to elicit changes in the discharge pattern of neurones in the lobster stomatogastric ganglion (Marder and Hooper, 1985; Harris-Warrick and Marder, 1991). 5-HT has also been shown to enhance sensitivity to light in crayfish retinal photoreceptors (Aréchiga et al. 1990).
It has been suggested that 5-HT induces the release of various neurohormones, such as the hyperglycaemic hormone in the crayfish Orconectes limosus (Keller and Beyer, 1968), the red and black pigment dispersing hormones in the fiddler crab Uca pugilator (Fingerman and Nagabhushanam, 1992) and the gonad-stimulating hormone in the crayfish
Procambarus clarkii (Sarojini et al. 1995a,b). Topical
application of 5-HT to the isolated eyestalk has been reported to modify electrical activity in neurosecretory cells (Nagano and Cooke, 1981). However, the possible role of 5-HT as a neurotransmitter or neuromodulator on neurosecretory cells of the crayfish eyestalk has not been confirmed (see Fingerman and Nagabhushanam, 1992; Lüschen et al. 1993). The presence of 5-HT, its biosynthesis and/or degradation have not been established in the isolated eyestalk in the specific regions where it would be expected to exert its physiological actions. In addition, the release of 5-HT in the eyestalk ganglia has not been explored.
It is the purpose of this study (a) to examine in the various regions of the crayfish eyestalk for the presence of 5-HT, its precursors, L-tryptophan (L-TRP) and 5-hydroxytryptophan OH-TRP) and its metabolites, 5-hydroxytryptophol (5-HTPH) and 5-hydroxy-indole-3-acetic acid (5-HIAA), produced after oxidation of 5-HT by the enzyme monoamine oxidase, and N-acetyl-5-HT (NA-5-HT), produced after acetylation by N-acetyltransferase, and to determine their regional distribution; (b) to determine the location of 5-HT-immunopositive cell bodies and axons in the eyestalk and optic nerve, particularly in the areas of efferent input to the neurosecretory cells in the X organ, and (c) to measure the release of 5-HT from the isolated eyestalk after stimulation.
Materials and methods
The experiments were carried out using adult crayfishes
Procambarus clarkii (Girard) of either sex and in intermoult
at the time of the experiment. The crayfish were collected from Rio Conchos, Chihuahua, México, and adapted to laboratory conditions for 2 weeks, either under natural light:dark cycles or on a 12 h:12 h light:dark programme.
Eyestalks were excised and placed in chilled saline solution (van Harreveld, 1936). The exoskeleton, muscles and connective tissue sheath were removed, leaving intact the neural part of the optic peduncle, which was then divided by microdissection into four segments (see Fig. 1). However, we shall use the term eyestalk in this paper. Unless otherwise stated, all experiments were conducted at room temperature
(22–24 °C) and at night, when the 5-HT content has been reported to be at its highest (Fingerman and Fingerman, 1977; Fingerman et al. 1978).
Amine determinations
The amine determinations were made using high-performance liquid chromatography (HPLC) following techniques devised by Eloffson et al. (1982), Kilpatrick et al. (1986) and Leung and Tsao (1992). Eyestalks were dissected out, and groups of two eyestalks or eight eyestalk segments (see Fig. 1) were pooled and their mass recorded. The neural tissue was homogenized in 500µl of HClO4(100 mmol l−1)
containing variable amounts of 3,4-dihydroxybenzylamine (DHBA), and centrifuged at 1600 g for 20 min at 4 °C using a Sorvall centrifuge. The supernatant was filtered through a nylon sieve with a pore diameter of 0.22µm. The samples were stored at 0–5 °C until analysis, which was conducted on the same day. The protein content of the homogenized tissue was determined by a modification (Cerbón and Aréchiga, 1986) of the Lowry method using small samples of nervous tissue.
The samples were injected into a guard column of C-18, connected to a reverse-phase analytical column of C-18 (25 cm×4.6 mm) packed with a particle size of 5µm (LDC-Analytical). The amines were detected either by fluorescence or with an electrochemical detector. The HPLC system with fluorescence detector consisted of a programmable solvent delivery module (Waters, model 590), a universal LC injector (model U6K) and a scanning fluorescence detector (model 470). The signals were recorded on a data module (model 730). Fluorescence was detected at an excitation wavelength of 254 nm and an emission wavelength of 338 nm.
Electrochemical detection was carried out according to Kilpatrick et al. (1986). This HPLC system consisted of a model PM-60 pump (Bioanalytical System) and a Rheodyne 7125 injector. A 200µl sample loop was routinely employed. The volume eluted was analyzed by an amperometric detector LC4B (Bioanalytical System). A potential of 0.8 V, with respect to a Ag/AgCl reference electrode, was applied for oxidation of 5-HT. The current generated was recorded on an x,y chart recorder (Cole-Palmer).
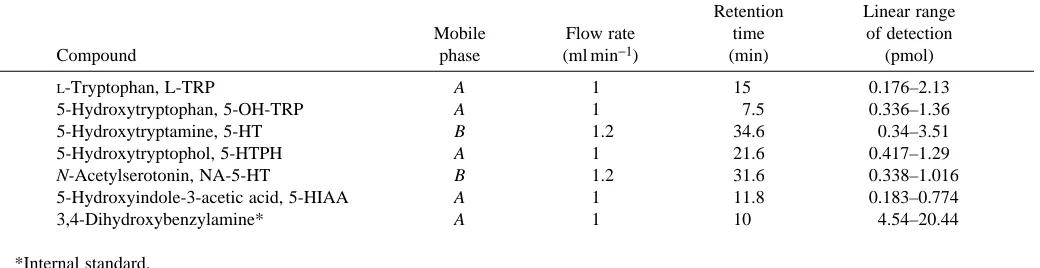
Three mobile phases were used. One of them (mobile phase A in Table 1) was described by Leung and Tsao (1992); its composition was as follows: sodium acetate (40 mmol l−1),
citric acid (10 mmol l−1), disodium EDTA (130µmol l−1),
octane-1-sulphonic acid (420µmol l−1), sodium chloride
(13 mmol l−1) and methanol 10 % (v/v), pH 4.68. Mobile phase
B only differed from phase A in the proportion of methanol, which was 20 % (v/v). It was used for the detection of 5-HT and NA-5-HT. A third mobile phase was used for electrochemical detection, with the following composition: sodium acetate (90 mmol l−1), citric acid (35 mmol l−1),
disodium EDTA (130µmol l−1), octane-1-sulphonic acid
(230µmol l−1) and methanol 10.5 % (v/v). The pH was adjusted
electrochemical detection, but was variable for fluorescence detection (see Table 1).
The amine solutions were freshly prepared for each experiment and dissolved in perchloric acid (100 mmol l−1).
With both procedures, the systems were calibrated to define the ranges of linearity for the following substances (all from Sigma Chemicals): L-tryptophan (L-TRP), 5-hydroxytryptophan OH-TRP), 5-hydroxytryptamine (5-HT), 5-hydroxytryptophol (5-HTPH), N-acetylserotonin (NA-5-HT), 5-hydroxy-indole-3-acetic acid (5-HIAA) and 3,4-dihydroxybenzylamine (DHBA), which was used as an internal standard. The amounts of the amines in the samples were calculated from the peak heights. For fluorescence detection, the linear range is indicated in Table 1; for electrochemical detection, the peak height of the current was linear from a minimum detection value of 0.15 pmol up to 1.5 pmol of 5-HT. The corresponding peaks in the chromatogram were confirmed by determining the increase in amplitude of the peak after adding pure substances to the column. The loss of substances during chromatography was estimated as 10–15 %, with DHBA as the internal standard.
Release of 5-HT
Eyestalks were incubated in the test solutions for 10 min. The perfusate was collected and HClO4 (5 mmol l−1) was
added. Samples were kept at 4 °C until chromatographed. In some experiments, 5-HT uptake was prevented by adding fluoxetine (Ely Lilly) (1µmol l−1) and monoamine oxidase was
inhibited using pargyline (Sigma Chemicals) (0.8–10 mmol l−1). When basal or high-[K+]-induced 5-HT
release was measured, the eyestalks were incubated in a 400µl pool in a modified van Harreveld’s saline (1936) (solution A in Table 2). For release induced by high K+concentrations in
the medium, K+ was substituted for Na+ in the bathing
solutions at several concentrations. The results reported here correspond to saline solutions with the compositions indicated in Table 2.
For electrically evoked release, the eyestalk was mounted in a 200µl pool of saline solution. Trains of pulses were delivered through bipolar metal microelectrodes inserted into the optic nerve (0.6–2µA, 50 ms, 2 Hz, for 10 min). Pulses were
generated with a Grass S-48 stimulator through a PSIU 6 stimulus isolation unit and monitored on a Tektronix 2201 oscilloscope.
Immunocytochemistry
Cellular localization of 5-HT was made by immunocytochemistry, using a primary antibody against 5-HT (rabbit anti-5-HT, Immunonuclear Corp.). The crayfish were anaesthetised in iced water (4 °C) and the whole preparation was removed by microdissection. Eyestalks were excised and the neural tissue was cleansed as described above. In some experiments, the eyestalk was removed with the optic nerve and the supraoesophageal ganglion attached.
The preparations were left for 3 h at 4 °C in 4 % paraformaldehyde, in phosphate buffer (PBS 0.1 mol l−1), and
[image:3.609.50.570.88.222.2]then transferred for 30 min to a 30 % sucrose solution. The sample was then frozen and 16–18µm thick cryostat sections prepared. In other experiments, in which the eyestalks were whole-mounted, the dissection and fixation procedures were the same, but the tissue was not incubated in the sucrose solution. Either sections or whole mounts were incubated for 18 h at room temperature with the primary antiserum. Various dilutions of the primary antiserum were tried and the best results were obtained with a 1:400 dilution in PBS containing 0.3 % Triton X-100. After washing, preparations were exposed to fluorescein-isothiocyanate-conjugated goat anti-rabbit IgG, which was used as the secondary antibody (see Aréchiga et al. 1990). It was diluted (1:20) in PBS for 1 h at 4 °C. In some experiments, preparations were dehydrated by successive Table 1. Chromatographic conditions for the measurement of 5-HT, its precursors and three metabolites by fluorescence
Retention Linear range
Mobile Flow rate time of detection
Compound phase (ml min−1) (min) (pmol)
L-Tryptophan, L-TRP A 1 15 0.176–2.13
5-Hydroxytryptophan, 5-OH-TRP A 1 7.5 0.336–1.36
5-Hydroxytryptamine, 5-HT B 1.2 34.6 0.34–3.51
5-Hydroxytryptophol, 5-HTPH A 1 21.6 0.417–1.29
N-Acetylserotonin, NA-5-HT B 1.2 31.6 0.338–1.016
5-Hydroxyindole-3-acetic acid, 5-HIAA A 1 11.8 0.183–0.774
3,4-Dihydroxybenzylamine* A 1 10 4.54–20.44
*Internal standard.
Table 2. Composition of solutions used for eyestalk
superfusion and/or incubation
Solution Na+ K+ Ca2+ Mg2+ Cl− Hepes
A 205 5.4 13.5 2.6 242.6 2.5
B 205 5.4 0.1 16 242.6 2.5
C 130.4 80.0 13.5 2.6 242.6 2.5
D 130.4 80.0 0.1 16 242.6 2.5
Concentrations are given in mol l−1.
[image:3.609.316.564.644.711.2]incubations in ethanol at increasing concentrations from 10 % to 100 % and finally cleared in methyl salicylate. Preparations were observed under a fluorescence microscope (Zeiss).
Results
Regional distribution of 5-HT, its precursors and metabolites in the eyestalk
To determine the regional distribution of 5-HT, the eyestalks were divided into four segments, as shown in Fig. 1, which presents the proportion of 5-HT contained in each segment (hatched bars) as a percentage of the total content, which was 3.14±1.6 pmol per eyestalk (95.4 pg mg−1of wet tissue). The
specific 5-HT content (fmolµg−1 of protein) was also
determined (black bars). The most distal segment, comprising the retina and the lamina ganglionaris (R-LG), had a low total 5-HT content (9.9 %), as well as a low specific content (1.1 fmolµg−1protein), while the medulla terminalis (MT) and
optic nerve (ON) had the highest proportion in the eyestalk (40.2 %). However, the specific content of this region had only an intermediate value (13.5 fmolµg−1protein). The two middle
segments corresponded to the medulla externa (ME) and the medulla interna (MI); these had intermediate proportions of 5-HT (27.2 % in the ME and 22.8 % in the MI) and high specific
contents (17.3 fmolµg−1protein in the ME and
22.4 fmolµg−1protein in the MI) (see Table 3).
The regional distribution was also determined for the 5-HT precursors L-tryptophan (L-TRP) and 5-hydroxytryptophan (5-OH-TRP) and for three of its metabolites, 5-hydroxytryptophol (5-HTPH), 5-hydroxy-indole-3-acetic acid (5-HIAA) and N-acetylserotonin (NA-5-HT). As can be seen in Table 3, all six substances were present in the various regions of the eyestalk, and wide differences in regional content were apparent. The specific content of the precursors was higher than that of either 5-HT or its metabolites, except for 5-OH-TRP in MT, as seen in Table 3. The highest specific content of L-TRP was found in the retina lamina ganglionaris segment (see Table 3).
5-HT inactivation by monoamine oxidase in the eyestalk
The inactivation of 5-HT by monoamine oxidase (MAO) was tested by using pargyline, an inhibitor of MAO activity (see Neff and Yang, 1974). Groups of five eyestalks each were incubated for 2 h with various concentrations of pargyline. Experiments were carried around midnight, when MAO has been found in other nocturnal crustaceans to be most active (Fingerman et al. 1978). Within the range 0.8–10 mmol l−1, pargyline induces a dose-dependent
increase of 5-HT content and a simultaneous decrease in the
60
30
0
5-HT content (%)
[5-HT] (fmol
µ
g
−1protein)
4
2
0
5 5
60
30
0
40
20
0
5 5
60
30
0
40
20
0 4
4
60
30
0
40
20
0 6
6 R
LG
ME
MI
MT
[image:4.609.246.561.380.734.2]ON Fig. 1. Regional distribution of 5-hydroxytryptamine
content of 5-HTPH (Fig. 2). Control groups of eyestalks incubated in van Harreveld’s solution (Solution A in Table 2) did not show any significant changes in either HT or 5-HTPH within the 2 h period.
5-HT biosynthesis in the eyestalk
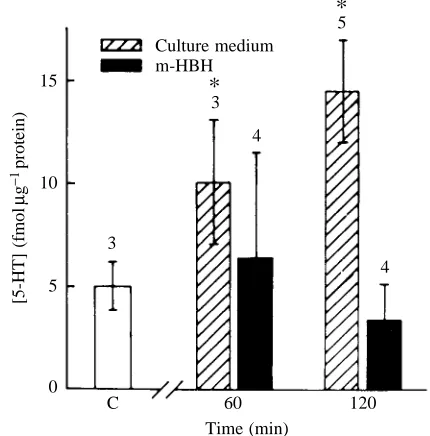
The biosynthesis of 5-HT from 5-OH-TRP was explored by using m-hydroxybenzyl-hydrazine (m-HBH), a selective blocker of 5-OH-TRP decarboxylase (Falck et al. 1966). Basal 5-HT content was determined, and groups of nine eyestalks were incubated in 80 mmol l−1K+solution for 30 min (Solution
C in Table 2). As shown below, this procedure releases 5-HT in the eyestalk. Samples of the depleted eyestalks (three each) were taken for 5-HT determination. A pool of six eyestalks was transferred to 20 % Leibovitz L-15 culture medium in normal saline solution, and samples consisting of three eyestalks were
taken after 60 and 120 min intervals to determine 5-HT content. Another pool of six eyestalks was incubated in the same culture-medium-enriched saline solution to which m-HBH (2 mg ml−1) had been added, and samples were also taken at
[image:5.609.45.569.84.197.2]intervals of 60 and 120 min for 5-HT determination. This protocol was repeated several times (Fig. 3). All experiments were conducted at room temperature (22–24 °C), near dusk. As seen in Fig. 3, the 5-HT content in the eyestalks incubated in the inhibitor remained unchanged during the 2 h period. This was in contrast to the increase in 5-HT content seen after a 2 h incubation in culture medium.
Table 3. Content and regional distribution of 5-HT and related substances in the eyestalk
ES R-LG ME MI MT
Protein content 325.1±101 178.7±103.1 26.5±9.2 13.5±12 54.8±19.6
(µg per structure)
L-TRP 353.3±65.6 177.4±79.4 38.2±23.5 40.2±14.6 91.2±60.1
5-OH-TRP 57.6±37.8 25.5±4.2 18.3±9.4 45.7±26.1 7.1±3.5
5-HT 9.6±4.9 1.10±0.22 17.3±13.9 22.4±10.3 13.5±5.03
5-HIAA 7.1±5.9 0.7±0.5 14.1±12.1 2.3±1.9 4.3±2.07
5-HTPH 3.1±0.7 2.0±0.5 17.6±7.8 24.8±10.3 6.7±0.93
NA-5-HT 6.1±3.8 1.5±0.4 17.6±11.1 12.2±2.4 9.1±6.8
Values are expressed as fmolµg−1protein and are presented as means ± standard deviation (N=5).
[image:5.609.332.547.417.635.2]ES, eyestalk; R-LG, retina and lamina ganglionaris; ME, medulla externa; MI, medulla interna; MT, medulla terminalis. For abbreviations of chemicals, see Table 1.
Fig. 2. Effect of pargyline on the content of 5-hydroxytryptamine (5-HT, filled circles) and its metabolite 5-hydroxytryptophol (5-HTPH; open circles). Higher pargyline concentrations result in a greater reduction of 5-HT degradation into 5-HTPH. All values correspond to mean and standard deviations of five determinations.
25
20
15
10
5
0
[5-HT] (fmol
µ
g
−1protein)
[5-HTPH] (fmol
µ
g
−1protein)
5-HTPH 5-HT
5
4
3
2
1
0
[image:5.609.63.291.475.677.2]0.8 1.6 4.0 10.0
[Pargyline] (mmol l−1)
Fig. 3. Blockage of 5-HT biosynthesis by m-hydroxybenzyl-hydrazine (m-HBH). Bars indicate the amount of 5-HT present in controls after 5-HT depletion (control, C) and after incubation in culture medium for 60 and 120 min, both in untreated medium (hatched bars) and after addition of m-HBH to the medium (black bars). Values represent mean ± S.D. The number of experiments is indicated above the bars; the asterisk indicates a significant difference (P<0.05, student’s t-test) compared with the content of the depleted eyestalk.
*
*
15
10
5
0
[5-HT] (fmol
µ
g
−1protein)
Culture medium m-HBH
C 60 120
3
3
4
5
4
Immunocytochemical localization of 5-HT-containing structures
The distribution of 5-HT-immunopositive cell bodies and fibres was assessed using an antibody against 5-HT. Whole mounts of eyestalks were used. Immunoreactive cells were found in all eyestalk regions, confirming results from other authors. The labelled somata and fibres were widely distributed. Various types of cell bodies were found in all four neuropile regions of the eyestalk (see Elofsson, 1983; Sandeman et al. 1988). The 5-HT-like immunopositivity found
in the lamina ganglionaris was described in detail in an earlier communication (Aréchiga et al. 1990). Both the medulla externa and the medulla interna contain immunopositive cell bodies and fibres. This is consistent with the HPLC assessment, which indicates the presence of 5-HT in all the eyestalk ganglia. Since the distribution of 5-HT-containing elements in the ME and MI was similar to that reported in detail for other crustacean species, no further analysis was made in our preparations.
Of particular relevance to the role of 5-HT in the modulation of neurosecretory activity (see Sáenz et al. 1997) is the presence of abundant immunopositive fibres in the neuropile of the medulla terminalis, near the area where the X organ cells are known to branch. As seen in Fig. 4, a group of immunopositive cell bodies (10–15 of them) is located in the area immediately above the hemi-ellipsoid body. The
diameters of the cells range from 15 to 40µm. Profuse branchings of axons and abundant varicosities of these neurones in the neuropile of the medulla terminalis were observed in all preparations.
Interestingly, two separate bundles of 10–15 immunopositive axons each were identified in the optic nerve, one running along the medial side of the nerve and the other along the lateral side (Figs 5, 6). All fibres branched in the neuropile of the medulla terminalis (Fig. 5A). Their number and location were consistent in eight preparations. Their course and pattern of endings suggested that they might be efferent axons running from the supraoesophageal ganglion to the medulla terminalis. A group of cell bodies in the protocerebral lobe of the supraoesophageal ganglion, which were consistently stained, may be the origin of the efferent fibres to the medulla terminalis (Fig. 5B). However, this point requires further elucidation.
To test the efferent nature of these fibres, in five animals, one optic nerve was looped with a fine nylon thread, the ligature was removed after 72 h, and the two eyestalks, still attached by the optic nerves to the supraoesophageal ganglion, were excised and processed for immunocytochemistry. As seen in Fig. 6, the immunopositive fibres in the medial bundle are no longer present in the segment distal to the ligature, whereas those in the proximal segment appear dilated. Axons in the lateral bundle are still present in the distal stump.
A
B
hb
hb
[image:6.609.91.509.400.710.2]X-o
X-o
Release of 5-HT
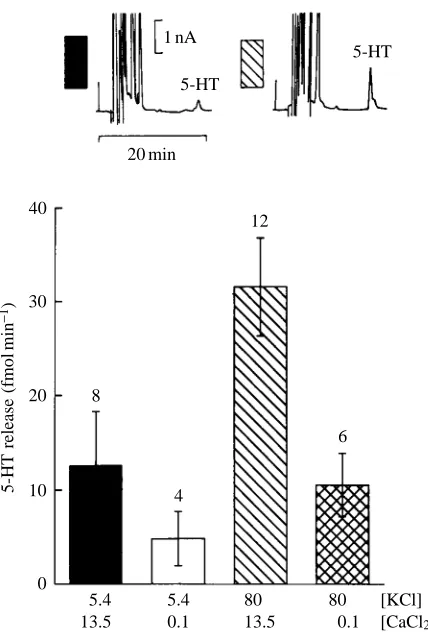
Both basal and evoked 5-HT release were examined. To enhance recovery, the active uptake of 5-HT in the eyestalk was inhibited by fluoxetine (Fuller et al. 1991; Chen and Reith, 1994). Four groups of four eyestalks each were incubated for 10 min in 400µl of solution A (see Table 2) to which fluoxetine (1µmol l−1) was added. As shown in Fig. 7, the basal release
of 5-HT in the eyestalk is increased over twofold after incubation in fluoxetine.
Two methods were used to test the evoked release of 5-HT from the eyestalk. From a group of 30 eyestalks, 12 were incubated in a solution in which a high concentration of K+
(80 mmol l−1) was substituted for the equivalent amount of Na+
(Solution C in Table 2). As shown in Fig. 8, the release of 5-HT, which under control conditions is very low, even after suppressing 5-HT uptake with fluoxetine, is greatly increased by incubation for 30 min in the high-[K+] solution.
To test the possible dependence of 5-HT release on [Ca2+] o,
Ca2+was substituted by Mg2+in the bathing fluid, leaving only
0.1 mmol l−1[Ca2+]
o(solutions B and D in Table 2). Groups of
six eyestalks were incubated in low-[Ca2+]
osolution, three of
them in normal [K+]
o and three in 80 mmol l−1 [K+]o. This
procedure was repeated several times, as indicated in Fig. 8. Lowering [Ca2+]
o greatly reduced both basal and the
high-[K+]-induced 5-HT release.
The other way of exploring 5-HT release was by selectively stimulating the optic nerve with brief electric pulses of varying intensities, as described in Materials and methods. These experiments were carried out on individual eyestalks, stimulated with pulses of 50 ms at 2 Hz for 10 min. As shown in Fig. 9, in 28 preparations, trains of pulses delivered to the optic nerve are capable of inducing a considerable release of 5-HT to the medium, in proportion to the intensity of stimulating current applied to the nerve. Again, 5-HT release is reduced by lowering [Ca2+]
oin the bathing fluid (not shown
in Fig 9). After trains of pulses, a lasting depletion of 5-HTwas commonly observed for as long as 30 min. However, in most preparations, 5-HT release almost completely recovered after 2–3 h.
As can be seen in the chromatograms shown in Figs 8 and 9, both the K+-induced and the electrically evoked stimulation
also caused the release of 5-HT-related substances. These compounds were not identified in the experiments reported here.
A
B
hb
on
sog
on
mt
on
mt
M
[image:7.609.103.503.71.405.2]L
Discussion
The total content of 5-HT in the eyestalk of Procambarus
clarkii (95.4 pg mg−1wet tissue) is close to the values reported
for other crayfish; for example, 100 pg mg−1wet tissue
determined by Eloffson et al. (1982) in Pacifastacus leniusculus and 102 pg mg−1wet tissue by Kulkarni and Fingerman (1992)
in Procambarus clarkii. It is also similar to values found in other crustacean species (Laxmyr, 1984). There is no other systematic survey of the regional distribution of 5-HT in crustaceans, but since 5-HT-like immunopositive cells have been found in the various eyestalk regions, it is not surprising to find 5-HT precursors and metabolites as well. The variability of the values may be related to differences in the manipulation of the tissue and the time between excision and measurement, which could result in a lowering of 5-HT content.
[image:8.609.56.541.69.374.2]The low content of 5-HT in the retina lamina ganglionaris segment was to be expected, since most of it is non-neural tissue, including the pigment cells and crystalline elements. A similar situation may explain the difference between the proportional and specific contents of 5-HT in the segment comprising the medulla terminalis and the distal end of the optic nerve; although the proportion is highest in this part of the eyestalk, the specific content is lower than that of the ME and MI. Whereas the mass of this segment (and its protein content) is more than twice that of ME and four times that of
Fig. 6. Reconstruction from serial photomicrographs of a crayfish optic nerve, showing the loss of medial immunopositive axons in the distal stump, 72 h after ligating the nerve. The arrow indicates the centripetal direction towards the supraoesophageal ganglion (see inset). Scale bar, 200µm. M, medial; L, lateral.
18
12
6
0
5-HT release (fmol
min
−
1)
C Fluoxetine
Fig. 7. Fluoxetine increases the release of 5-HT from the crayfish eyestalk. 5-HT release per eyestalk was measured before and after adding fluoxetine (1µmol l−1) to the bathing medium. Black bar, control (C). Hatched bar, after addition of fluoxetine. Values are means ±S.D., N=4.
M
[image:8.609.59.278.461.678.2]the MI (see Table 3), a large proportion of its mass corresponds to optic nerve axons, only a few of which are 5-HT-immunopositive, and to the neurosecretory system of the X organ, also devoid of HT. In fact, as seen in Figs 4 and 5, 5-HT immunopositivity in the MT is located in the neuropile branches and in a few somata, and it is remarkable that these elements have such a high content of 5-HT. The high content of L-TRP in the same region, compared with the low content in the haemolymph (van Marrewijk and Ravestein, 1974), suggests an active uptake mechanism for L-TRP, hitherto unexplored, and presumably active biosynthesis of 5-HT. Immunocytochemical evidence has identified a profuse network of 5-HT-like immunopositive cell bodies and fibres in the R-LG region (Aréchiga et al. 1990). These might be the sites of 5-HT biosynthesis. This would be in agreement with the role proposed for 5-HT as a local modulator of retinal activity (Aréchiga et al. 1990).
The biosynthesis of 5-HT in the crayfish eyestalk is clearly suggested by the presence of its precursor 5-OH-TRP, with a similar regional distribution to that of the amine. The
suppression induced by 5-HT synthesis inhibitors on the restitution of 5-HT levels after its depletion lends support to this notion. This is in agreement with similar findings in the crustacean Upogebia littoralis, by Marmaras and Fragoulis (1970) and Fingerman et al. (1978) in the crab Uca pugilator, where observations were made on intact animals. Our present evidence supports the hypothesis of a process of 5-HT biosynthesis intrinsic to the eyestalk.
The enhancement of 5-HT recovery from the perfusion fluid after incubation in fluoxetine as well as the rise of 5-HT content induced by pargyline suggest that two of the common mechanisms of 5-HT inactivation are present in the crayfish eyestalk: uptake of 5-HT, which is blocked by fluoxetine (Fuller et al. 1991; Chen and Reith, 1994), and 5-HT oxidation by MAO, which is inhibited by pargyline, as demonstrated earlier in Uca pugilator (Fingerman et al. 1974, 1978) but not detected by Barker et al. (1972) and Kennedy (1978) in the lobster Homarus americanus. From the evidence we present here, it is reasonable to conclude that MAO activity is present in the crayfish eyestalk.
The presence of NA-5-HT suggests a role for acetylation in the degradation pathway of 5-HT in the eyestalk. No particular emphasis was placed in this study on exploring the activity of the N-acetyltransferase system after finding the metabolite NA-5-HT, since the activity of this enzyme has already been
30 40
20
10
0
5-HT release (fmol
min
−1)
5.4 13.5
5.4 0.1
[KCl] [CaCl2] 80
13.5
80 0.1 8
4
12
6 20 min
1 nA
5-HT
[image:9.609.67.281.69.385.2]5-HT
Fig. 8. Release of 5-HT induced by incubation of the eyestalk in saline solutions containing a high K+concentration (80 mmol l−1) and its dependence on Ca2+concentration. The amount of 5-HT released per eyestalk was measured. Values are means ±S.D. and the number of experiments is indicated above each bar. Concentrations of KCl and CaCl2are indicated (in mmol l−1) under each bar. HPLC traces in the inset correspond to the control (black bar) and release of 5-HT after incubation in a high K+concentration (see text).
75
50
25
0
5-HT release (fmol
min
−1)
14
6
5
3 20 min
1 nA 5-HT
5-HT
B C
D A
0 1 2
[image:9.609.342.535.71.357.2]Stimulation intensity (µA)
established in the prawn Macrobrachium rosenbergii (Withyachumnarnkul et al. 1992, 1993). The interaction of this
system with that of MAO in the crayfish remains to be elucidated.
The location of immunopositive elements is similar to that described for other crustacean species (Elofsson, 1983; Sandeman et al. 1988). Of particular interest in this study is the large 5-HT content and the profuse ramifications of 5-HT-containing fibres present in the medulla terminalis neuropile, the probable site of arrival of efferent input to the X organ cells. The efferent nature of the immunopositive axons identified in the optic nerve is supported by their disappearance in the distal stump of the optic nerve after ligation and their continued presence in the proximal stump. These may be the same axons in which dense granules have been described, both in the optic nerve (Larimer and Smith, 1980) and in the neuropile (Andrew and Saleuddin, 1978). The possible site of origin of the immunopositive axons in the optic nerve may be the small immunopositive neurones present in the protocerebral lobe of the supraoesophageal ganglion; further studies are needed to confirm this, since other 5-HT-like immunopositive cell somata have been described in various central ganglia, although they are not known to send axons to the optic nerve (Sandeman et al. 1988).
The maximal release of 5-HT evoked by stimulation corresponds to approximately 2–3 % of the total 5-HT content. It is comparable to values found in other systems, such as the giant cerebral neurones in Aplysia californica (Gerschenfeld et
al. 1978) and the Retzius cell of the leech (Henderson, 1983).
The dependence of 5-HT release on [Ca2+]
osuggests that there
is an exocytotic mechanism, such as has been amply documented for other 5-HT-releasing preparations (Gerschenfeld et al. 1978; Livingstone et al. 1981; Henderson,
1983). The larger amount of 5-HT released by electrical stimulation compared with incubation in high-[K+] solutions
suggests that short, phasic stimuli are more efficient in eliciting release than a sustained, constant depolarization. The protocol of stimulation was chosen after testing various combinations of rate, voltage and train duration. The observation that other amines are co-released with 5-HT by high [K+] and electrical
stimulation is not surprising since the stimulation was not restricted to serotonergic neurones.
The widespread distribution of 5-HT in the various eyestalk neuropiles suggests that it may have a multiplicity of functions. So far, only the effects on the retina (Aréchiga et al. 1990) and on neurosecretory activity (Fingerman and Nagabhushanam, 1992; Sáenz et al. 1997) have been described. A more detailed search for other possible functions is necessary.
The results reported here support the concept that 5-HT is a neurotransmitter or a modulator in the crayfish eyestalk. Besides its modulatory role in retinal activity, it may act at other levels of the eyestalk, as suggested by the wide distribution of neurones immunopositive to the anti-5-HT antiserum. The presence of a mesh of immunopositive axons in the neuropile of the medulla terminalis points to a role in the modulation of neurosecretory activity at that level. This
view is also supported by the demonstration of a direct action of 5-HT on X organ neurones (see Sáenz et al. 1997).
The authors are indebted to Elizabeth Becerra and Victor Anaya for valuable assistance in various aspects of the experimental work. This project was partly supported by CONACyT grant no. 0804-N9110.
References
ANDREW, R. D. ANDSALEUDDIN, A. S. (1978). Structure and innervation
of a crustacean neurosecretory cell. Can. J. Zool. 56, 423–430.
ARÉCHIGA, H., BAÑUELOS, E., FRIXIONE, E., PICONES, A. AND
RODRIGUEZ-SOSA, L. (1990). Modulation of crayfish retinal
sensitivity by 5-hydroxytryptamine. J. exp. Biol. 150, 123–143.
BARKER, D. L., MOLINOFF, P. B. AND KRAVITZ, E. A. (1972).
Octopamine in the lobster nervous system. Nature New Biol. 263, 61–62.
BELLON-HUMBERT, C. AND VAN HERP, F. (1988). Localization of
serotonin-like immunoreactivity in the eyestalk of the prawn Palaemon serratus (Crustacea, Decapoda, Natantia) J. Morph. 196, 307–320.
BELTZ, B. S. ANDKRAVITZ, E. A. (1983). Mapping of serotonin-like immunoreactivity in the lobster nervous system. J. Neurosci. 3, 585–602.
CERBÓN, J. ANDARÉCHIGA, H. (1986). Characterization of the Na+, K+
ATPase in a microsomal preparation from the abdominal ganglia of crayfish. Effect of neurodepressing hormone (NDH). Comp. Biochem. Physiol. 85B, 561–566.
CHEN, N. H. ANDREITH, M. E. A. (1994). Effects of locally applied cocaine, lidocaine and various uptake blockers on monoamine transmission in the ventral tegmental area of freely moving rats: a microdialysis study on monoamine interrelationships. J. Neurochem. 63, 1701–1713.
DIXON, D. AND ATWOOD, H. L. (1985). Crayfish motor nerve terminal’s response to serotonin examined by intracellular microelectrode. J. Neurobiol. 16, 409–424.
DUDEL, J. (1965). Facilitatory effects of 5-hydroxytryptamine on the crayfish neuromuscular junction. Naunyn-Schmiedebergs Arch. Pharmak. 249, 515–528.
ELOFSSON, R. (1983). 5-HT-like immunoreactivity in the central
nervous system of the crayfish Pacifastacus leniusculus. Cell Tissue Res. 232, 221–236.
ELOFSSON, R., LAXMYR, L., ROSENGREN, E. ANDHANSSON, C. (1982).
Identification and quantitative measurements of biogenic amines and DOPA in the central nervous system and haemolymph of the crayfish Pacifastacus leniusculus (Crustacea). Comp. Biochem. Physiol. 71C, 195–201.
FALCK, B., OWMAN, C. H. ANDROSENGREN, E. (1966). Changes in rat pineal stores of 5-hydroxytryptamine after inhibition of its synthesis or break-down. Acta physiol. scand. 67, 300–305.
FINGERMAN, M., JULIAN, W. E., SPIRTES, M. A. ANDKOSTRSEWA, R.
M. (1974). The presence of 5-hydroxytryptamine in the eyestalks and brain of the fiddler crab Uca pugilator, its quantitative modification by pharmacological agents and possible role as a neurotransmitter in controlling the release of red pigment-dispersing hormone. Comp. gen. Pharmac. 5, 299–303.
FINGERMAN, M. AND NAGABHUSHANAM, R. (1992). Control of the
FINGERMAN, M., SCHULTZ, R. E., BORDLEE, B. P. ANDDALTON, D. P. (1978). Twenty-four hour variation of 5-hydroxytryptophan-decarboxylase and monoamine oxidase activities in the eyestalks of the fiddler crab, Uca pugilator. Comp. Biochem. Physiol. 61C, 171–175.
FINGERMAN, S. W. ANDFINGERMAN, M. (1977). Circadian variation in
the levels of red pigment-dispersing hormone and 5-hydroxytryptamine in the eyestalks of the fiddler crab, Uca pugilator. Comp. Biochem. Physiol. 56C, 5–8.
FISCHER, L. AND FLOREY, E. (1983). Modulation of synaptic
transmission and excitation–contraction coupling in the opener muscle of the crayfish, Astacus leptodactylus, by 5-hydroxytryptamine and octopamine. J. exp. Biol. 102, 187–198.
FULLER, R. W., WONG, D. T. AND ROBERTSON, D. W. (1991).
Fluoxetine, a selective inhibitor of serotonin uptake. Med. Res. Rev. 11, 17–34.
GERSCHENFELD, H. M., HAMON, H. AND PAUPARDIN-TRITSCH, D.
(1978). Release of endogenous serotonin from two identified serotonin containing neurones and the physiological role of serotonin reuptake. J. Physiol., Lond. 274, 265–278.
GLANZMAN, D. L. AND KRASNE, F. B. (1986).
5,7-Dihydroxytryptamine lesions of crayfish serotonin-containing neurones: Effect on the lateral giant escape reaction. J. Neurosci. 6, 1560–1569.
GLUSMAN, S. ANDKRAVITZ, E. A. (1982). The action of serotonin on
excitatory nerve terminals in lobster nerve–muscle preparations. J. Physiol., Lond. 325, 223–241.
HARRIS-WARRICK, R. M. AND MARDER, E. (1991). Modulation of
neural networks for behavior. A. Rev. Neurosci. 14, 39–57.
HENDERSON, L. P. (1983). The role of 5-hydroxytryptamine as a
transmitter between identified leech neurones in culture. J. Physiol., Lond. 339, 309–324.
KELLER, R. ANDBEYER, J. (1968). Zur hyperglykämischen Wirkung
von Serotonin und Augenstielextrakt beim Flusskrebs Orconectes limosus. Z. vergl. Physiol. 59, 78–85.
KENNEDY, M. B. (1978). Products of biogenic amine metabolism in
the lobster: sulfate conjugates. J. Neurochem. 30, 315–320.
KILPATRICK, I. C., JONES, M. W. ANDPHILLIPSON, O. T. (1986). A
semiautomated analysis method for catecholamines, indoleamines and some prominent metabolites in microdissected regions of the nervous system: An isocratic HPLC technique employing coulometric detection and minimal sample preparation. J. Neurochem. 46, 1865–1876.
KRAVITZ, E. A. (1988). Hormonal control of behavior: amines and
the biasing of behavioral output in lobsters. Science 241, 1775–1781.
KULKARNI, G. K. ANDFINGERMAN, M. (1992). Quantitative analysis
by reverse phase high performance liquid chromatography of 5-hydroxytryptamine in the central nervous system of the red swamp crayfish, Procambarus clarkii. Biol. Bull. mar. biol. Lab., Woods Hole 182, 341–347.
LARIMER, J. L. ANDSMITH, J. T. F. (1980). Circadian rhythm of retinal
sensitivity in crayfish: modulation by the cerebral and optic ganglia. J. comp. Physiol. A 136, 313–326.
LAXMYR, L. (1984). Biogenic amines and DOPA in the central
nervous system of decapod crustaceans. Comp. Biochem. Physiol. 77C, 139–143.
LEUNG, P. Y. ANDTSAO, C. S. (1992). Preparation of an optimum mobile phase for the simultaneous determination of neurochemicals in mouse brain tissues by high- performance liquid chromatography with electrochemical detection. J. Chromatogr. 576, 245–254.
LIVINGSTONE, M. S., HARRIS-WARRICK, R. M. ANDKRAVITZ, E. A.
(1980). Serotonin and octopamine produce opposite postures in lobsters. Science 208, 76–79.
LIVINGSTONE, M. S., SCHAEFFER, S. F. ANDKRAVITZ, E. A. (1981).
Biochemistry and ultrastructure of serotonergic nerve endings in the lobster: Serotonin and octopamine are contained in different nerve endings. J. Neurobiol. 12, 27–54.
LÜSCHEN, W., WILLING, A. AND JAROS, P. P. (1993). The role of
biogenic amines in the control of blood glucose level in the decapod crustacean Carcinus maenas L. Comp. Biochem. Physiol. 105C, 291–296.
MA, P. M., BELTZ, B. S. AND KRAVITZ, E. A. (1992). Serotonin-containing neurones in lobsters: Their role as gain-setters in postural control mechanisms. J. Neurophysiol. 68, 36–54.
MARDER, E. ANDHOOPER, S. L. (1985). Neurotransmitter modulation
of stomatogastric ganglion of decapod crustaceans. In Model Neural Circuits and Behavior (ed. A. I. Selverston), pp. 319–337. New York: Plenum Press.
MARMARAS, V. J. AND FRAGOULIS, E. G. (1970).
5-Hydroxytryptophan-decarboxylase activity in the decapod crustacean Upogebia littoralis. Experientia 26, 359–360.
NAGANO, M. ANDCOOKE, I. M. (1981). Electrical activity in the crab
X organ sinus gland system. Site of initiation, ionic bases and pharmacology. In Neurosecretion: Molecules, Cells, Systems (ed. D. S. Farner and K. Lederis), pp. 504–505. New York: Plenum Press.
NEFF, N. H. AND YANG, H. Y. T. (1974). Another look at the monoamine oxidase and the monoamine oxidase inhibitor drugs. Life Sci. 14, 2061–2074.
SÁENZ, F., GARCIA, U., AND ARÉCHIGA, H. (1997). Modulation of electrical activity by 5-hydroxytryptamine in crayfish neurosecretory cells. J. exp. Biol. 200, 3079–3090.
SANDEMAN, D. C., SANDEMAN, R. E. ANDAITKEN, A. R. (1988). Atlas
of serotonin containing neurones in the optic lobes and brain of the crayfish, Cherax destructor. J. comp. Neurol. 269, 465–478.
SAROJINI, R., NAGABHUSHANAM, R., DEVI, M. AND FINGERMAN, M.
(1995a). Dopaminergic inhibition of 5-hydroxytryptamine-stimulated testicular maturation in the fiddler crab, Uca pugilator. Comp. Biochem. Physiol. 111C, 287–292.
SAROJINI, R., NAGABHUSHANAM, R. ANDFINGERMAN, M. (1995b). In
vivo inhibition by dopamine of 5-hydroxydopamine-stimulated ovarian maturation in the red swamp crayfish, Procambarus clarkii. Experientia 51, 156–158.
VANHARREVELD, A. (1936). A physiological solution for freshwater
crustaceans. Proc. Soc. exp. Biol. Med. 34, 428–432.
VANMARREWIKJ, W. J. A. ANDRAVESTEIN, H. J. L. (1974). Amino
acid metabolism of Astacus leptodactylus Esch. I. Composition of the free and protein-bound aminoacids in different organs of the crayfish. Comp. Biochem. Physiol. 47B, 531–542.
WITHYACHUMNARNKUL, B., BUPPANIROJ, K. AND PONGSA
-ASAWAPAIBOON, A. (1992). N-Acetyltransferase and melatonin
levels in the optic lobe of giant freshwater prawns, Macrobrachium rosenbergii de Man. Comp. Biochem. Physiol. 102A, 703–707.
WITHYACHUMNARNKUL, B., PONGSA-ASAWAPAIBOON, A. AND
POOLSANGUAN, B. (1993). Characteristics of the enzyme
N-acetyltransferase in the optic lobe of the giant freshwater prawn, Macrobrachium rosenbergii de Man. Comp. Biochem. Physiol. 104B, 449–454.