Wtth 14 text-figures Printed tn Great Britain
FUNCTIONING OF THE INHIBITORY CONDITIONING
AXON INNERVATING INSECT MUSCLES*
BY GRAHAM HOYLE
Department of Biology, University of Oregon
{Received 19 July 1965)
The jumping muscle of locusts is supplied by three motor axons (Hoyle, 1955 a), one of which gives rise to large twitch contractions when stimulated with a single shock and is termed the fast (F) axon, whilst another gives rise to slow contractions only, when repetitively stimulated, and is termed the slow (S) axon (Hoyle, 1955 ft). The third axon was found not to give a contraction when stimulated alone but it did cause a hyperpolarization in some of the muscle fibres Thus, by comparison with peripheral inhibitory action at crustacean neuromusclar junctions (Fatt & Katz, 1953), amphibian heart (Hutter & Trautwein, 1956) and cat spinal cord (Eccles, 1953), its function could be expected to be inhibitory; but Hoyle (19556) found that stimulation of this axon at the same time as stimulation of either of the two excitatory ones did not lead to mechanical inhibition. Instead, the mechanical responses were enhanced, though the effect was slight so functional significance could not be ascribed to it with confidence. Since this phenomenon could not be fully understood it has remained in the background awaiting further investigation.
In the meantime, some pharmacological aspects of the third axon function have been studied in Schistocerca gregaria by Usherwood & Grundfest (1964, 1965). In conflict with the earlier finding, these authors found the axon partially to inhibit the contrac-tion caused by the S, though not the F, axon. The present work was undertaken to repeat the experiments on the third axon and to extend them to include other species and if possible other muscles. During the course of an investigation on learning of leg-raising, a good preparation for the study of the mode of action of an axon having similar properties to third axon of the jumping muscle was found (Hoyle, 1965 b), and thi3 has been extensively used in the present work.
This muscle is the anterior coxal adductor (a.c.a.), and it receives but a single excitatory axon. The results show that the conflicting data of Hoyle and of Usherwood & Grundfest can be resolved, because different preparations of the same nerve-muscle combination may show either of the two effects—reduction or enhancement of tension —and in two instances the mechanical contraction of the same preparation has at different times been facilitated or inhibited by the same axon. Anticipating this result, the axon which can function so diversely will be termed the ' inhibitory-conditioning axon' and will be given the abbreviation I-C. Its function has been studied both in isolated preparations and in intact and headless insects, where the objective has been to discover, by the long-term examination of the patterns of its discharges, the ways in which the axon is utilized in life, and the ways in which its discharges are associated with excitatory axon activity during spontaneous and reflexly evoked discharges.
• Supported by research grant NSF GB3160.
43°
GRAHAM HOYLEMATERIALS AND METHODS
The work has been done mainly on the metathoracic a.c.a. muscles of the African locust Schistocerca gregaria, and confirmed on the grasshopper Melanoplus differ entialis, the Florida lubber grasshopper Romalea wicroptera and the American locust Schisto-cerca vaga. In most of the present work the functioning of the axon was examined in
the whole insect, with a minimal amount of dissection. This will be referred to as the in situ preparation. In addition, various degrees of isolation from the body of the insect
Forceps tips
force transducer
Intracellular electrode
Anterior
Anterior coxal adductor
Posterior coxa! rotator
Posterior coxal adductor
Posterior
Fig. i. Drawing of coxal region of right metathoracic leg of SMttocerca gregaria after minimum dissection, clamping of cut apodeme of anterior coxal adductor by force transducer and place-ment of intracellular recording electrode.
In situ preparation
To examine the functioning under minimal dissection insects were secured, back down, in soft dental wax with the right metathoracic leg twisted forwards (cf. Hoyle, 1965 b). The loose connective between the rim of the coxa and the thorax was then cut away, exposing the underlying coxal and trochanteral muscles. The a.c.a. can be seen emerging from below the posterior rotator of the coxa, immediately posterior to a large tracheal trunk (Fig. 1). The muscle is attached to the coxal rim via a short apodeme. A semicircular cut was made through the rim close to the apodeme to free the muscle. The apodeme was then clamped by the tips of a micro-forceps mechano-electronic force transducer (Hoyle & Smyth, 1963).
Dental wax was shaped around the exposed coxa to make a cup which was filled with locust saline (Hoyle, 1953) replenished at frequent intervals. A ground lead was placed in the saline. A glass capillary microelectrode was next micro-manipulated into position and inserted into an a.c.a. muscle fibre. The lead was direct-coupled, and together with that of the force transducer, monitored on a Tektronix 502 cathode-ray oscilloscope and displayed continually on an Offner Dynograph pen oscillograph.
The isolated preparation was described in a previous paper (Hoyle, 1966). During early stages of the investigation attempts were made partially to isolate the preparation, retaining only connexions with the tracheal system intact. Eventually it was realized that the preparation survives sufficiently well in complete isolation.
RESULTS
Tests on fully and semi-isolated preparations
The excitatory axon was usually stimulated by applying leads to the nerve and raising the stimulus strength to just above threshold. In some cases it was found con-venient to excite it by preganglionic stimulation applied to a nerve stump. The I-C axon was routinely stimulated by preganglionic stimulation whenever an effective site, usually the left connective with the mesothoric ganglion or sometimes part of the ganglion itself, could be found. Otherwise, the only way I-C action could be studied was by further raising the strength of the stimulus applied to nerve 3 c, thereby exciting the excitor and inhibitor axons together. This method is disadvantageous com-pared with preganglionic stimulation because it precludes the varying of the interval between excitatory and inhibitory impulses, so that the relative times of arrival at the junctions cannot be varied. The interval may be critical in relation to the inhibitory aspect of functioning.
In all, 72 preparations of either semi- or fully isolated kinds were studied. Of these 27 showed mechanical inhibition of excitatory axon contraction to greater or lesser degrees (Usherwood & Gmndfest phenomenon); 24 showed a slight enhancement of the excitatory response (Hoyle phenomenon); 2 preparations showed a slight inhibi-tory effect followed by a slight enhancement during continued interaction of the two axons; 19 preparations did not show any significant mechanical influences of I-C upon E contractions, although they did show electrical effects. It cannot be too strongly emphasized that these results related to the use of standard locust saline. Quite different results may be obtained in different media. For example, higher levels of
432 GRAHAM HOYLE
calcium ions increase the size of the excitatory junctional potentials and lead to greater" twitch contraction. Standard locust saline of Hoyle (1953) was not intended to be an optimal one as far as mechanical responses are concerned, but to give results similar to average results obtained with the muscles bathed in their own haemolymph. For descriptive purposes, the two different positive effects of I-C will be considered independently. First, inhibitory effects will be dealt with, then facilitating ones.
I I I I I I I I I I I I I I 1 1 i 1 1 1 1 1 1 1 1 1
I ) I I I I M I I I I I I M l I I I I I I I I I ) | | | ' I I I I I I M I I I I I I M I I I I I 1 I I I I I M | | 1
" 11111111111 i i i i n 111 m i | | 11 i n n i i n 1111 n i ii i n H i ii 1111
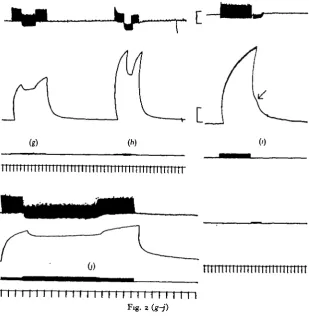
Fig. 2 (a-/). Legend on opposite page.
Mechanical inhibitory effects of I-C
three principal effects were seen. One was a relatively quick fall (complete in 1 sec.) in tension to a plateau (Fig. 26), which was maintained throughout inhibition. This effect reduced tetanus tension by not more than 20%. On cessation of stimulation tension returned to the tetanus level. A second kind was a very slow progressive fall in tension reaching a plateau after as long as 20 sec. (Fig. ze); in extreme cases tension fell to about 50% its initial value. The third was a relatively quick fall, followed by a
(g)
[image:5.451.78.388.155.474.2]I I I I I I I I I I I I H I I I I I 1 Fig. 2 (g-j)
Fig. 2. Responses of various preparations to stimulation of the excitatory and inhibitory-conditioning axons. Upper traces in each give intracellular recordings from average muscle fibres; second trace gives tension; third trace, or third and fourth traces, stimulus marks; fifth trace, time in seconds. Calibration: 10 mV; 2-5 g. (a), O'$ g. (b-j).
slow recovery of tension, in spite of continued inhibitory stimulation (Fig. 2/), which in some cases was complete; i.e. the inhibitory effect waned progressively until full tetanic tension was restored.
Stimulation of the I-C axon when it is acting as an inhibitor, at the same time as the excitor, leads to a slower rise in tetanic tension (Fig. 3 a, b). Stimulation of the axon at the end of a tetanus, during relaxation, gives rise to a quicker decay of tension (Fig. 2i).
434 GRAHAM HOYLE
though not at low, frequencies of I-C. This effect could be explained on the basis of a certain amount of facilitation of the inhibitory junctional potentials in these prepara-tions. However, facilitation of I-C junctional potentials was not common and in many muscle fibres the first I-C junctional potential was the largest, with a decline during
(b)
Fig. 3. Influence of I-C on rate of rise of tetanus and on twitch height at optimal phasing for inhibition, Schutocerca gregaria. (a) io/sec.; first response E alone; second E and I. Upper trace, intracellular, second trace, tension; third trace, time in seconds. (6) 20/sec. (smooth tetanus); details same as (a), (c) I-C axon was stimulated at io/sec, E at i/sec. First six responses at sub-optimal phasing, last three at optimal phasing: note reduction of both e.j.p. and twitch height. I-C j p.s depolarizing in this fibre. Upper trace, intracellular; second trace, tension; third trace, I-C stimulus; fourth trace, E stimulus; fifth trace, time in seconds. Calibration: io mV; i-og. in (a) and (A) 0^25 g. in (c).
(tii) Influence of phasing. Whenever the I-C axon could be excited independently the influence of varying the interval between I-C and E upon tension was examined. This alters the relative times of arrival, or phasing, of the two impulses at the neuro-muscular junctions. There were many examples in which there was no influence of phasing, the mechanical inhibitory effect, if any, remaining constant (Fig. 4). Nor could any preparations which did not show mechanical inhibition with coincident stimulation come to show it at some optimal phasing.
-I 1 1 1
\
1 1
Fig. 4. Lack of influence of phasing on both junctional potential and twitch height of whole muscle. This result was not uncommon. S. gregaria preparation. Upper trace, intracellular; second trace, tension; third trace, I-C stimulus; fourth trace, E stimulus; fifth trace, time in seconds.
Some preparations did, however, show variation in extent of attenuation of the twitch height in association with altered phasing. The same preparations did not, however, show any effect of phasing at tetanic frequencies.
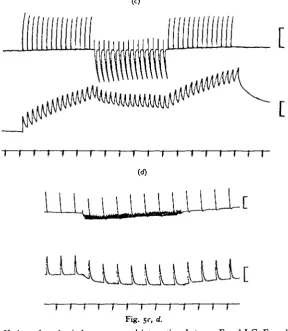
(iv) Relaxing action of I-C acting alone. It has already been noted that activity in the I-C axon can cause an increased rate of relaxation of the tetanus. In some preparations, both isolated and relatively intact, in which E was silent, activity in the I-C axon caused relaxation of passive tension (at mean body length) or tension obtained by stretch (Hoyle, 1966). A similar effect was sometimes seen in the present work (Fig. $d), but so also was contraction (Fig. 5a). The majority of muscles showed no mechanical effect of I-C acting alone.
Electrical effects of I-C
436 GRAHAM HOYLE
I I I I I I I I I I I I I I I I
U
' " ' ' I ' l l 11111111 ( I
J I
t t i ; t ) i i r
of the membrane was nevertheless apparent. This may be attributed to an undershoot of the repolarizing phase of the e.j.p.s during I-C j.p. action (Fig. 4a, b). Some showed the general polarization accompanied by enhancement of the e.j.p. (Fig. 2d). An enhancement of e.j.p.s is to be expected on biophysical grounds only if the inhibitory
(c)
1 1 1 ( 1 1 i 1 1 1 1 1 1 1 ( 1 r
1 i 1 1 1 1 1 i
Fig. 5<-. <*•
Fig. 5. Variety of mechanical responses and interactions between E and I-C. Four different
S. gregaria preparations, (a) I-C was stimulated first and gave rise to a slight contraction
(starts at arrow). E j.p.s markedly attenuated at optimal phasing but mechanical response was not affected significantly. Upper trace, intracellular; second trace, tension; third trace, I-C stimulus; fourth trace, E stimulus; fifth trace, time in seconds. (6) Mechanical response markedly reduced by I-C action. Other details as in (a), (c) Giant-sized I-C j.p.s and marked attenuation of e.j.p.s (70 % in this fibre). Partial relaxation of summated tension is caused in whole muscle. Upper trace, intracellular; second trace, tension; third trace, time in seconds. Calibration; 10 mV.; 0-5 g. (d) This preparation developed a weak contracture after prolonged stimulation at o-o/sec. Brief twitches were superimposed on the contracture. The contracture was markedly reduced by I-C stimulation at 15/sec. and the twitch height was slightly enhanced. Other details as in (c).
[image:9.451.81.370.141.473.2]by i.j.p. action. Very large attenuations in Crustacea are considered to be caused by presynaptic inhibition (cf. Dudel & Kuffler, 1961, for crayfish). Enhancements (Fig. 6) may require explanation by the converse process—presynaptic facilitation.
Mechanical facilitation
Enhancement of contraction can be obtained by stimulating the I-C axon at the same time as the E axon in about one-third of all preparations. It is seen as a progressive increase in height of the twitch during repetitive stimulation of I-C (Fig. 7). The extent of the enhancement is related to the frequency of firing of I-C, but only within narrow limits and not in a clear manner. The twitch height, in these examples, in-creases with increasing frequency only up to about 10 I-C/sec. and further increase in frequency has no additional effect; or instead a decrease, accompanied by relaxation of summated tension, may occur, as the inhibitory effect of I-C takes over following extensive depolarization of the fibres.
I I I I I I I I I I I M | I I I I I I I I I I I I I I I I I I I I I I [ I I I I I I I I I I I I I I I I ) I I I I IJ I I I I I I I I I I
Fig. 7. Facilitating action of I-C on twitch response. E axon was stimulated at i/gec. I-C axon fired spontaneously at progressively increasing frequency; note that twitch force doubles. Upper trace, intracellular; middle trace, tension; lower trace, time in seconds. Calibration: iomV.; 0-5 g.
Electrical events accompanying facilitation of tension development
Tension development by a muscle is considered to be directly linked to the mem-brane potential and starts when the memmem-brane is depolarized below a threshold (Hodgkin & Horowicz, i960; Orkand, 1962). Hence contractions could not be caused by the polarizing actions of the I-C axon. Muscle fibres which may be responsible for the mechanical facilitation observed have been located by micro-electrode probing in various parts of the muscle. They appear to be mixed randomly with fibres showing only polarizing responses, but themselves give depolarizing responses (Figs. 8, 9). If these depolarizing I-C j.p.s are responsible for tension enhancement, and if they are sufficiently large, they should give rise to small contractions; and such contractions have been observed. Contractions caused by I-C are of the slow, tonic kind, never even minutely twitch-like. The occurrence of such slow contractions only suggests the presence of specialized muscle fibres having slow contractile properties.
Specialized muscle fibres
44° GRAHAM HOYLE
plateaux of as much as 40 mV. (Figs. 9, 10). Recovery from such a peak may take as long as 3 min. This slow depolarization is not an artifact associated with the con-traction of the whole muscle, for its time-constant is much longer than that of relaxa-tion of the whole muscle.
I-C junctional potentials of both polarizing and depolarizing kinds occur in these fibres. When evoked at the same time as e.j.p.s they simply summate with the accumulating slow waves. Even quite large polarizing potentials fail to counteract the slow, late depolarization wave (Fig. 9 a). No explanation can be proposed at the present for this strange depolarization.
I I I I I I I I I I I I I I I I I I I [ I I I I I I I i I I I I I I I
—1—1—1—1—1—(—i—1—1—1—1—1—1—1—1—t—1—1—(—1—1—1—1—»—r—1—1—1 r
Fig. 8. Slow electrical after-responses occurring in certain muscle fibres only (ScbUtocerca
gregaria) I-C j.p.s were evoked throughout the record at i/sec. In this fibre they were small
and depolarizing. A single E j.p. was elicited. After completion of the response, in this case with a small undershoot, the slow response starts, reaching a peak at the point shown by the arrow. That the change is not an artifact is shown by the reversal of the I-C j.p. which occurs at the same time. Full recovery takes 9 sec. Upper trace, intracellular record; second trace, tension; third trace, I-C stimuli; fourth trace, E stimulus; fifth trace, time in seconds. Calibra-tion: iomV.; O'S g.
During repetitive stimulation of the excitatory axon at frequencies as low as 2—3/sec. the slow waves will summate to a level which exceeds the contraction-coupling threshold. Thus a very low frequency of discharge could give rise to a maintained, tonic contraction suitable for postural adjustment. It may be that these muscle fibres represent a specialized set concerned with muscle tone.
Reversal potential for effects of I-C axon
reversal of the sign of the junctional potential occurs are usually carried out by altering the membrane potential artificially through a second intracellular electrode.
Unfortunately, the more interesting depolarizing effects in this preparation occur in deeper fibres and it has so far proved difficult to study them satisfactorily by the double-electrode technique. However, the late, slow depolarizing wave acts as an effective means of altering the membrane potential. By stimulating the I-C axon during
60 mV.
442 GRAHAM HOYLE
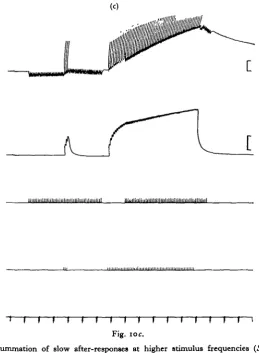
the slow recovery the reversal potential may be determined. At peak depolarization the i.j.p.s are usually polarizing potentials. They progressively decline in height as the membrane potential rises, until they disappear. As the recovery continues and the membrane potential rises further, they become depolarizing and gradually increase in size (Fig. 10). The point at which they vanish is the reversal potential and is
pre-niiiiiiiniiiiiiiiiiiihiiiiiiiiniiiiiiiniiil luuiiiiiiiiiiiiiiiiiiiiiiiiimiiiiiiiiiiiiiiiiiiini
r T 1 1 r 1
Fig. ioc.
I I F I
Fig. 10. Summation of slow after-responses at higher stimulus frequencies {ScHstocerca
gregaria). (a) I-C stimulated at i/sec, E at lo/sec. (6) Another fibre, same preparation as (a).
E stimulated at 3/sec. (c) Another preparation: small polarizing I-C j.p.s. Note that they are not increased during the summated wave but instead decrease, suggesting that I-C equilibrium potential has fallen. Other details as in Figure 96.
sumably equal to the equilibrium potential for the conductance increase caused by the I-C axon. According to Usherwood & Grundfest (1965) this increase is a selective
one to chloride ions.
[image:15.451.96.356.143.497.2]444
GRAHAM HOYLEPre-synaptic effects in fibres showing depolarizing I-C potentials
In these fibres there is usually very little effect of varying the timing of excitation of the two axons. In some, however, there is a slight attenuation. This is probably due to the equilibrium potential for the I-C junctional potential being below the equi-librium potential for the e.j.p.
Preliminary structural evidence for two kinds of muscle fibre
The muscle is being examined electron-microscopically and has already been found to have many interesting features which will be reported in detail elsewhere. The various fibres have similar, long (6/J) sarcomere lengths and have a similar basic
-T,
ry-i ry-i n ry-i n ry-i n ry-i l l I I I I I I I I I I I I I I I I I I I | | | I I I | i )
Fig. I I . Spontaneous burst activity of I-C axon occurring in almost intact preparation during silent period of E axon (Melanoplus differentiaUs). Note slight falls in resting tension during high-frequency firing of I-C alone. Upper trace, intracellular; second trace, tension; third trace, time in seconds. Calibration: 10 mV.; 0 1 g.
445
Inhibitory and conditioning effects in the life of the insect
The results described above do not leave one with even the possibility of making a definite proposal regarding the natural function of the I-C axon in the intact insect. Its effects depend so markedly on the state of the muscle at the time of the experiment
~wr^vv%^w#\^^
[image:17.451.45.404.349.562.2]ir ir
~r
Fig. 12. Spontaneous I-C axon activity combined with sporadic E axon activity. Three preparations; Melanoplus differentialu muscle bathed in haemolymph. Note in all: reduction of tension with high-frequency I-C bursts occurring alone; facditation of twitches associated with a high frequency of I-C activity. Upper trace, intracellular; second trace, tension; third trace, time in seconds. Calibration: io mV.; 0-2 g.
and on the proportions of the functionally opposed inhibitory and facilitatory effects in different fibres. It is, of course, possible that the state of affairs in the intact animal is quite different. The muscle is bathed in haemolymph which contains, in addition to mineral ions, organic materials including neurohumours which might affect the neuromuscular properties. Therefore there exists the possibility that in the intact
446 GRAHAM HOYLE
organism the function might be clear-cut and either wholly inhibitory or wholly facilita-tory, even though experimental preparations have given confusing and conflicting results.
The mode of action of the axon will depend on the ways in which it is utilized in natural reflex actions. For example if the I-C axon is active at the end of E axon bursts
~v v v v
r
~v
~y v v \>Fig. 12b. For legend see p. 445.
it might be aiding relaxation. If it is active during contraction of antagonistic muscles it is probably a peripheral inhibitor, and so on. If the normal actions in the intact animal can be determined it might be relatively easy to resolve the problem. Recordings of I-C action in relation to E activity made during spontaneous activity of the isolated preparation of ganglion-nerve-muscle (Hoyle, 1966) did not give any clues as to its role.
In preparations, it has not been possible, with the possible exception of the opener of the claw of crayfish (Wilson & Davis, 1965), to show that the axon is actually an inhibitory one.
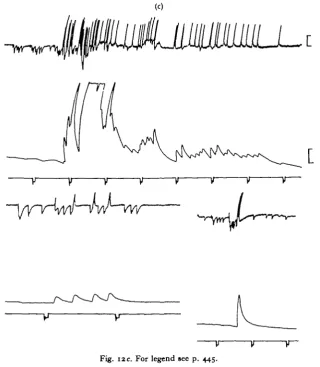
[image:19.451.66.387.137.505.2]The problem was examined in the present work in the insects Schxstocerca gregaria and Melanophis differentiatis by recording from the metathoric a.c.a. muscles in situ,
Fig. I2c. For legend see p. 445.
with haemolymph bathing the muscle, and the minimum of dissection—just sufficient to expose the muscle. These experiments have been carried out both with and without preparation of the muscle for tension recording, and both with and without the head. Preparations have been watched continually for several hours at a time, and several thousand active bursts have been examined. Removing the head has surprisingly little effect on the detailed patterns of activity occurring in the muscle. The spon-taneously occurring activity is full of variety and, although certain kinds of activity recur in a roughly similar way it may be stated that no exactly similar pattern of E and I-C impulse bursts ever occurs twice.
448 GRAHAM HOYLE
Spontaneous activity
During the long periods of fairly regular discharge of the E axon, the I-C axon seldom fires, though sometimes it fires also for long periods, its frequency changing approximately in parallel with changes in the E. From time to time, especially in intact insects, less frequently in beheaded preparations, the continuous background dis-charge is replaced by periods of silence alternating with bursts of firing. These are
f
i 11 i i i i i i i M i i 1 1 i M i i i i i i i i i i i ) 111 i i i i i i i i i I I i i i i i 1111 1I i i i i i i i i i i i i i i i 11 r
111 11111111111
1111111111 I III111111 111 11 I 1111111II 11111111 11) I) 111111 1111
wjaii^taiiuiL
I I I I I I I I I | | I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I M I I I I I I I I I I I I
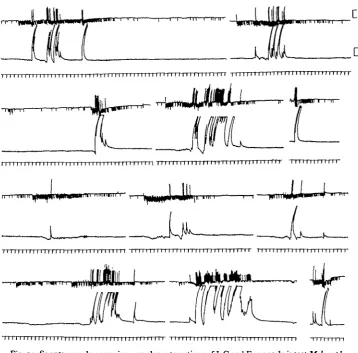
probably associated with attempts to turn right-way-up, and to escape or kick away the wax, etc. It is during these periods that one may expect to see the I-C axon put to good use. What invariably happens is that the I-C axon fires during the rare silent periods of the E axon, often in little bursts (Fig. 11). Return to firing of the E axon is
almost invariably preceded by a burst of firing of I-C which continues into the E dis-charge period for a short time, then stops (Fig. 12).
UJM,
I I I H I I I I I I I I I I I I I I I I I I I I I 1 I I I I I I I I I 1 I I I I I I I I I I I I I I I 1 I I I I I I t I I I I I I I I I 1 r I I I I I I 11 n I I I r i I
T i l l I I 1 IT Ti
rr
'\mmrrr-n-111111 [ '\mmrrr-n-111111 '\mmrrr-n-111111'\mmrrr-n-111111 '\mmrrr-n-111111'\mmrrr-n-111111111 ri 1 n i n n 11111 n r '\mmrrr-n-11111111
[image:21.451.46.405.149.502.2]-J-TTTTTTTTTTTTTTTTTTTT1
Fig. 14. Spontaneously occurring complex interactions of I-C and E in nearly intact Melanoplus
differentialis preparations. In these results a quick alternation of E and I-C occurred which
resulted in rapid contractions altering with rapid fluctuations in tension. Upper trace, intra-cellular records; second trace, tension; third trace, time in seconds. Calibration: iomV.;o-2g.
Another fairly frequent occurrence is discharge of I-C during a particularly vigorous burst of E activity.
450 GRAHAM HOYLE
rapid repolarization than occurs passively. Bursts of I-C impulses may occur during silent periods in the E discharge. In many instances the resting tension falls during these, especially when there is extensive summation of the I-C impulses (Figs. 11, 12). Reflexly evoked activity
Tactile stimulation applied to the animal anywhere on its body calls forth some change in the output to the a.c.a. Usually, this consists of a brief burst of increased frequency in E, followed by a silent period, and perhaps a few additional bursts. Sometimes there is a simple, abrupt cessation. This is most readily elicited by applying air puffs to the anal cerci. Cereal stimulation evokes the familiar leg extension-jump reflex, and the role of the a.c.a. in this is to relax.
The existence of this simple, marked central inhibition (cf. Hoyle, 1966) shows that the E motor neuron can be fully and satisfactorily inhibited centrally, therefore peri-pheral inhibition is not essential for normal functioning—at least in this reflex. In many instances the central inhibition was found to be accompanied by firing of I-C, as if to accelerate relaxation and, as it were, to assist the central action. However, this action of I-C builds up to a peak immediately before E starts to fire gain, and, as is the case during spontaneous fluctuations, usually overlaps the E burst (Figs. 13, 14). The reflexly evoked activity is, unfortunately, extremely variable in spite of careful control of the stimulation—a common experience with insects.
DISCUSSION
Many questions of physiological and biophysical interest are raised by the fore-going, but our present task is to consider the light shed by these results on the probable natural function of the I-C axon.
No evidence has been obtained from the intact animal which would support the view that the I-C axon is utilized as a simple peripheral inhibitor. Instances of its firing during contraction of antagonistic muscles have been observed, but they do not occur consistently. Firing during vigorous excitor activity in what was considered to be a purposeful response was commonly encountered. The data obtained from semi-isolated preparations were equivocal, since not all the a.c.a. muscles tested showed inhibition of contraction by I-C and many showed an opposite effect at low frequencies of firing of E.
One possibility which must be considered is that the I-C axon functions to aid relaxation. The natural relaxation rate is slow, possibly because of the presence in the muscle of 'slow' muscle fibres. A peripheral aid to their relaxation might be function-ally valuable and I-C undoubtedly increases the relaxation rate. The effect is, however, quite small in many instances and it is difficult to believe that it could be of vital importance. Such marginal effects may, of course, be of significance in the more stressful conditions obtaining in the wild.
that here the I-C axon is utilized primarily in the rare periods when the E axon is silent and especially just before it resumes firing. If it were to be functioning in this way in order to ensure a large contraction, then the mechanical response immediately following the I-C burst should be distinctly larger than one caused by a similar E burst which is not preceded by I-C activity. These contractions are indeed larger; a com-parison of twitch heights in Fig. 13 will show that those which are preceded by I-C activity are larger than ones occurring alone. Once again, however, the effect is prob-ably not great enough to warrant the development of a special axon.
It may be, of course, that the I-C axon is an evolutionary relic (whose primeval function was peripheral inhibition) from a time before central inhibition had been evolved by arthropods. If so, it may have acquired a new function. However, a full consideration of its functions can probably not be made without taking into account the influence on contraction—and behaviour—of fluctuating haemolymph composition (cf. Hoyle, 1965 a).
In searching for all possible roles it should be recalled that the Russian school of comparative physiology (e.g. Voskresenskaya, 1959; Onianie, 1964) has long been drawing attention to the probable existence of nerves which have a 'conditioning' effect on muscular contraction. A function of this kind might be considered for the locust and grasshopper I-C axons.
One finding of note is that the percentage of muscle fibres which give large electrical potential changes when I-C is stimulated is high in the anterior coxal adductor, being at least 60%. In the extensor tibiae it is much smaller, perhaps less than 30%. The bundle of muscle fibres attached to the most proximal margin of the extensor apodeme contains a much higher proportion of fibres responding to I-C than the muscle as a whole. This bundle also has a high proportion of dually excitatory-innervated muscle fibres, including some which receive the S, but not the F, axon (Cerf, Gmndfest, Hoyle & McCann, 1959). The muscle fibres which have an S axon supply (not more than about 30% in the jumping muscle—Hoyle, 19556) do all the work of postural maintenance and ordinary locomotion. They are therefore under almost constant neuromuscular barrage. Indeed, activity which continued non-stop at frequencies from 9/sec. to 130/sec. over an 8-day period has been observed in a locust nymph with trailing recording leads implanted in its metathoracic extensor tibiae (see Hoyle, 1964, for technique).
The a.c.a. is a continuously active postural muscle discharging at rates from 8/sec. to over 100/sec. in the intact animal. Its muscle fibres are all functioning in a manner comparable to the few dually excitatory-innervated ones of the jumping muscle. Since these muscle fibres do not get an opportunity to rest they may tend to become filled with an undesirably high concentration of some ion species. It is these same muscle fibres which show large I-C axon electrical activity. Perhaps the I-C axon may be functioning to counteract or relieve ionic accumulations (or losses).
452 GRAHAM HOYLE
SUMMARY
1. An axon which, in addition to the single excitatory axon, innervates the anterior coxal adductor muscles of locusts and grasshoppers causes peripheral effects, some of which are comparable to those of crustacean peripheral inhibitory axons. Tetanus tension is reduced by the axon.
2. The axon was termed the inhibitory-conditioning axon (I-C). Resting tension was reduced in a few preparations by the action of the axon, and the relaxation rate was markedly increased.
3. In some preparations, however, the axon caused a slight contraction of the muscle, and paradoxically enhanced twitch tension.
4. It was found by intracellular recording that in some muscle fibres the electrical response to I-C is polarizing, but in others it is depolarizing.
5. A few (but variable number) of the muscle fibres in each preparation showed slow, summating, after-depolarizations following termination of the excitatory junctional potentials. This is a new phenomenon, and suggests that these fibres, or their neuromuscular junctions, have special properties.
6. A study was made of the reflex and spontaneous discharges of the excitatory and I-C axons in the nearly intact animal, and in preparations, in an attempt to determine the function of I-C.
7. No evidence was found that I-C is actually used as a peripheral inhibitor and its natural function remains enigmatical.
REFERENCES
ATWOOD, H. L. & DORAI-RAJ, B. S. (1964). Tension development and membrane responses in phasic and tonic muscle fibers of a crab. J. Cell. Comp. Phytiol. 64, 55—72.
ATWOOD, H. L., HOYLE, G. & SMYTH, T. J. (1965). Mechanical and electrical responses of single innervated crab muscle fibres. J. Phytiol. (in the Press).
CERF, J. A., GRUNDFEST, H., HOYLE, G. & MCCANN, F. V. (1959). The mechanism of dual
responsive-ness in muscle fibers of the grasshopper Romalea nucroptera. j . Gen. Phytiol. 43, 377-95. DORAI-RAJ, B. S. (1964). Diversity of crab fibers innervated by a single motor axon. J. Cell. Comp.
Phytiol. 64, 41-54.
DUDEL, J. & KUFFLER, S. W. (1961). Presynaptic inhibition at the crayfish neuromuscular junction.
J. Physiol. 155, 543-62.
ECCLES, J. C. (1953). The Neurophytiological Basis of Mind. Oxford University Press. 314 pp. FATT, P. & KATZ, B. (1953). The effect of inhibitory nerve impulses on a crustacean muscle fibre.
J. Phytiol. 131, 374-89.
HODGKIN, A. L. & HOROWICZ, P. (i960). The influence of potassium and chloride ions on the membrane potential of single muscle fibres. J. Phytiol. 148, 127—60.
HOYLE, G. (1953). Potassium ions and insect nerve muscle. J. Exp. Biol. 30, 121-35.
HOYLE, G. (1955a). The anatomy and innervation of locust skeletal muscle. Proc. Roy. Soc. B, 143,
281-92.
HOYLE, G. (19556). Neuromuscular mechanisms of a locust skeletal muscle. Proc. Roy. Soc. B, 143,
343-67-HOYLE, G. (1964). Exploration of neuronal mechanisms underlying behaviour in insects. In Neural
Theory and Modeling, pp. 346—76. Ed. R. Reiss. Stanford University Press.
HOYLE, G. (1965a). Neural control of skeletal muscle. In Phytiology of Insecta, vol. 2, pp. 203-232, 407-49. Ed. M. Rockstein. New York: Academic Press.
HOYLE, G. (19656). Neurophysiological studies on 'learning' in headless insects. In Physiology of
Insect Central Nervous System, ed. J. E. Treheme. Academic Press.
HOYLE, G. (1966). An isolated insect ganghon-nerve-muscle preparation. J. Exp. Biol. 44, 413-27. HOYLE, G. & SMYTH, T. (1963). Neuromuscular physiology of giant muscle fibers of a barnacle, Balanus
HOTTER, O. F. & TRAUTWEIN, W. (1956). Vagal and sympathetic effects on the pacemaker fibers in the sinus venosus of the heart. J. Gen. Physiol. 39, 715-33.
ONIANIE, T. H. (1964). Comparative Physiology of Neuromuscular Apparatus. Tbilisi. (In Russian.) ORKAND, R. K. (1962). The relation between membrane potential and contraction in single crayfish
muscle fibres. J. Physiol. 161, 143-59.
USHERWOOD, P. N. R. & GRUNDFEST, H. (1964). Inhibitory postsynaptic potentials in grasshopper muscle. Science, 143, 817-18.
USHERWOOD, P. N. R. & GRUNDFEST, H. (1965). Peripheral inhibition in skeletal muscles of insects.
J. Neuropkysiol. 38, 497-518.
VOSKRESENSKAYA, A. K. (1959). Functional Characteristics of the Nerve—Muscular System of Insects. Academy of Science of the USSR (Akademia Nauk SSR), Moscow. (In Russian)