J. exp. Biol. (198a), 99, 1-8 I With 3 figures
Mrinted in Great Britain
ION-DEPENDENT VISCOSITY OF
HOLOTHURIAN BODY WALL AND ITS IMPLICATIONS FOR
THE FUNCTIONAL MORPHOLOGY OF ECHINODERMS
BY JOHN P. EYLERS
Department of Zoology, Duke University, Durham, N.C. 27706*
(Received 28 September- 1981 - Accepted 14 January 1982)
SUMMARY
Dermis from the holothurian Thyone inermis was subjected to constant load and its rate of plastic deformation (creep) was used to calculate tissue viscosity. During these tests the material was bathed in distilled water, sea water, or solutions of sodium, potassium, calcium, and magnesium salts in various combinations. Distilled water caused a threefold increase in viscosity which was reversed by sea water. Sodium and potassium decreased viscosity, while calcium and magnesium in combination with sodium in-creased it. Ionic cross-links shielded by monovalent ions but facilitated by divalent cations are proposed to explain this behaviour, and the relation-ship between this and other anomalies of echinoderm connective tissue physi-ology is discussed.
INTRODUCTION
Connective tissues are vital to the function of every metazoan organism, but despite their wide range of mechanical properties they are remarkably similar in composition. Basically they are all aqueous proteoglycan gels of varying density in which may be embedded elastic and collagenous fibres, the latter sometimes serving as substrates for calcification. Closely packed fibres, as in tendon, and extensive calcification, as in bone, give these tissues an elastic nature which reflects the properties of their principal components. At the other extreme is vitreous humour, a viscous fluid which contains few inclusions. Between these extremes lie many tissues with complex viscoelastic behaviour not necessarily correlated with their fibre content, and it is clear that their characteristics must reflect properties of the gel. It follows that knowledge of the comparative mechanics and the molecular basis for the organization of connective tissue matrix is basic to our understanding of the functional morphology of metazoans and the selective pressures which shaped their evolution.
The purpose of this paper is to present mechanical experiments on the dermis of holothurian body wall and to interpret the results in the light of a model based on the physical chemistry of gels. Dermal tissue was chosen because it can be obtained in large quantities convenient for fashioning laboratory preparations. The ability of
2 J O H N P. EYLERS
holothurians to stiffen their body walls when subjected to physical stress has b e e j reported (Stott et al. 1974; Freinkel & Hepburn, 1975), and aclose correlation betweefP environmental ionic composition and rheological behaviour of connective tissues has been demonstrated in two other echinoderm classes (Takahashi, 19676 ; Wilkie, 1978 A). Together these observations suggest the possibility of active control over extra-cellular matrix properties. This investigation was designed to determine the effects of inorganic ions on the long term load response of the dermis.
MATERIALS AND METHODS
Holothurians of the species Thyone inermis Heller (Mortensen, 1927) were collected from the Firth of Clyde near Millport, Scotland, and shipped to the University of Leeds, England, where they were held in a sea water aquarium at 5 °C. Animals chosen for testing were first washed thoroughly in distilled water. Immediately there-after the epithelium was scraped off the body surface with a razor blade, the body wall was split open lengthwise, and the viscera and coelomic lining removed with a blunt probe. Finally, the carcase was washed once again with distilled water and left to soak for at least half an hour. A pair of razor blades mounted on a handle were used to cut 3 mm strips of tissue parallel to the long axis to the body. Two aluminium rings were tied 1 cm apart to each strip with cotton thread. The thickness of each preparation was measured with a caliper, after which it was placed in one of several test solutions.
Mechanical testing apparatus
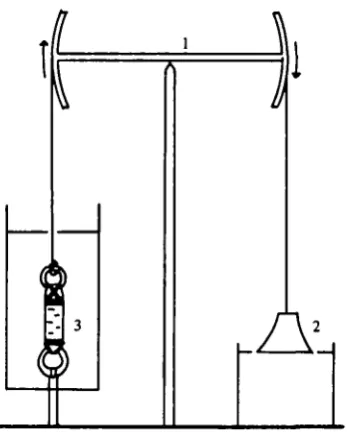
Creep tests (extension under constant load) were performed with a hyperbolic weight and balance beam device (Fig. 1) similar to mechanisms used in previous soft tissue studies (Alexander, 1962; Koehl, 1977). The ring at one end of the tissue strip was hooked to an immovable steel rod and the other ring was connected to the beam by an aluminium chain. Surrounding the specimen was an open-topped Plexiglass cylinder into which test solutions could be introduced with a syringe. The cylinder's volume was approximately 300 times that of the sample. A 10 g mass in the shape of a hyperbola of revolution was hung from the other end of the beam with its base just resting on the surface of a beaker of water. The curvature of the weight was so designed that as the tissue stretched and the weight fell into the water the negative buoyant force exactly compensated for the decreasing cross-sectional area of the specimen and the stress (<r = force/cross-sectional area) remained constant. While the tissue was being mounted the beam was stabilized by a thread which was burned to initiate the test. Tests ended after one to three hours when the tissue broke. A stylus attached to the beam recorded its movements on a smoke drum. All mechanical testing was conducted in a cold room at 9 °C.
Ion-dependent viscosity of hobtkurian body wall
t
1
f
i
is
i
'- 3II
1 \
[image:3.451.142.316.72.289.2]1
Fig. i. Balance beam and hyperbolic weight device for creep test*. The beam (i) transfers the force from the hanging weight (2) to the tissue preparation in a chamber filled with test solution (3). As the weight falls into the beaker of water buoyant force counteracts the effect of decreasing tissue cross-section and the stress remains constant.
Test solutions
The effect of high and low salinity on tissue creep was assessed by exposing the preparations to either distilled water (DW) or filtered sea water (SW). One group of matched specimens was soaked in SW for three hours before testing, another group was soaked in DW for three hours. In order to determine whether the effect of with-drawing ions could be reversed, a third group was first soaked in DW for three hours and then returned to SW for three more hours. Since the animals normally operate in a high salinity environment, a second protocol was designed to look for synergistic interactions among the common sea water cations. Sets of solutions were made up consisting of four different concentrations of sodium chloride each in pairwise combination with the chloride salts of potassium, calcium, or magnesium, also at four different molarities. Thus in each set there were sixteen different solutions in which sodium predominated and the other cation was present at or near its normal con-centration in sea water. The set of potassium-sodium solutions was tested three times, once on fresh tissue and twice on tissue which had been frozen in distilled water for one or two weeks. The other solutions were all tested once on frozen tissue.
RESULTS Strain vs. time: tissue viscosity
JOHN P. EYLERS
[image:4.451.68.353.49.234.2]10
Fig. 2. Strain v. time curves. Typical creep curves for tissue in distilled water (DW) and sea water (SW). Strain equals change in length (L-Lo) divided by starting length (Lo).
During the curvilinear phase a slight curl in the specimen was observed to straighten out, while during the linear phase the tissue necked down in the central region until it broke. The break never occurred near the threads where the rings were attached.
By applying a simple Maxwell model (Wainwright et al. 1976) to the linear region of the strain curve it is possible to calculate a viscosity (ij) for the tissue. Viscosity is defined as the ratio of the stress to the strain rate (tj = cr/i) and is reported in units of io3 Nm"8 s (GPa. sec in SI units). Used in this sense, viscosity is a normalized para-meter denning the resistance of the tissue to plastic flow under load. The higher the viscosity the greater the stress necessary to produce a given strain rate.
Effects of salinity and paired cations on viscosity
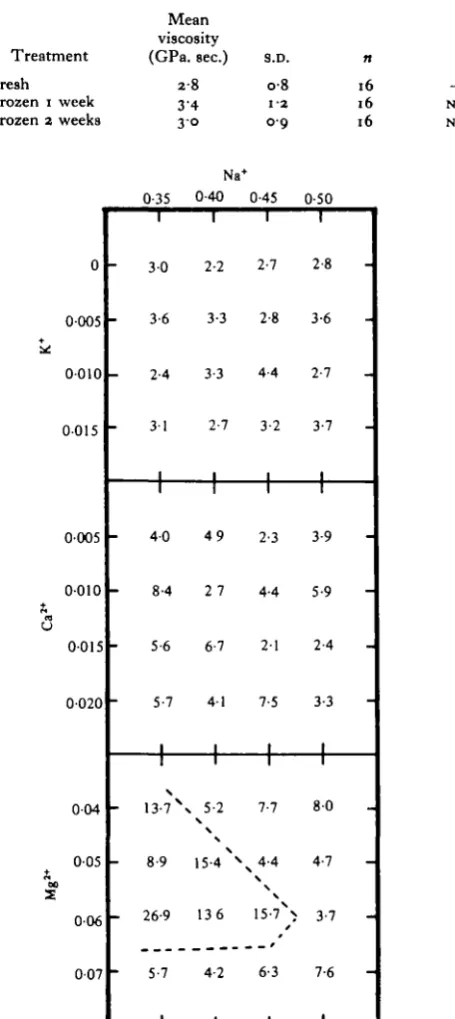
[image:4.451.100.356.545.628.2]The results of the sea water-distilled water experiments are presented in Table 1. Exposing the tissue to distilled water caused a threefold increase in the resistance to flow. However, the increase in viscosity is completely reversible, as demonstrated by the tissue which had first been soaked in distilled water and then in sea water. The results from the paired cation experiments convey considerably more information. In Table 2 the means and standard deviations for all sixteen solutions in the potassium-sodium set are reported for fresh and frozen tissue. Storage appears to have no effect on viscosity as there is no significant difference between fresh tissue and tissue frozen for one or two weeks. In Fig. 3, response surface maps summarize the results for each
Table 1. Effects of salinity on tissue viscosity
Treatment SW (3 h) DW (3 h)
DW (3 h) SW (3 h)
Mean viscosity (GPa. sec)
Si
15-8 5'4
S.D.
i-5 3 1 2-3
< OOOI
N.8.
SW: filtered sea water; D W : distilled water; S.D.: standard deviation; n: number of observations:
Ion-dependent viscosity of holotkurian body wall
Table 2. Effect of frozen storage on tissue viscosity in
potassium-sodium solutions
Treatment Fresh Frozen i week Frozen 2 weeks
Mean viscosity (GPa. sec.) 2-8 34 3° S.D. o-8 i-a oo 16 16 16 N.S. N.S. Na+ 0 0005 0010 0015 0005 0010 0015 0020 004 005 006 007 0-35 3 0 3-6 2-4 31 1
1
4 0 8-4 5-6 5-7 11
- 13-7\ 8-9- 26-9
5-7 0-40 2-2 3-3 3-3 2-7 1
1
4 9 2 7 6-7 41 1 1 5-21 5 - 4 ^
136 4-2 0-45 2-7 2-8 4-4 3-2 1
1
2-3 4-4 2-1 7-5 11
7-7x4 - 4 \\ 15-7% 6-3 0-50 2-8 3-6 2-7 3-7 I
1
3-9 5-9 2-4 3-3 1 1 8 0 4-7 3-7 7-6 I6 JOHN P. EYLERS
Table 3. x* analysis of cation-cation interactions in Figure 3
Mean viscosity
Treatment (GPa. sec) s.D. pi n x* D.F. P2
K x N a 3-1* o-6 — 16 097 9 > 0-99 C a x N a 4-6 1-9 < o-oi 16 8-05 9 — M g x N a 9-5 6-2 < o-ooi 16 2i'45 9 < 0-05
p 1: Student (-test of means; D.F. : degrees of freedom for x*; P2: probability that variations in response are random; • : pooled from Table 2.
of the cation pairs. These maps display the concentrations of the cations as co-ordinates on the x and y axes while the effect of each pair is plotted as an elevation on the z axis. A flat surface indicates that altering either variable has no effect, whereas sloped or folded surfaces imply synergistic interactions between the cations. An objective assessment of flatness may be derived from x2 analysis of the surface, the null hypothesis being that observed variations are randomly distributed. The results of x* tests along with the means and standard deviations for each response surface are given in Table 3. The presence of calcium and magnesium both cause a significant increase in tissue viscosity compared to the potassium-sodium set. There is a high probability that the variations in the potassium-sodium set are random, whereas it is nearly as likely that the magnesium-sodium results are grouped, indicating a synergistic interaction. An obvious collection of high values is outlined in the figure. It is not possible to make a definitive statement either way concerning the calcium-sodium set.
DISCUSSION
Ion-dependent viscosity of holotkurian body wall 7
K
iternal bulk fluid movement. At any instant the resistance to load is a combination f the time-independent entropy elasticity of the network (Wainwright et al. 1976) and the time-dependent penetration of solvent through it (Mow & Mansour, 1977). However, the equilibrium shape change is solely due to network properties (Mow & Lai, 1979).Creep occurs when cross-links are subjected to forces which exceed their strength. Adjacent polymers pull apart and move past one another to establish bonds at new positions in the network resulting in permanent plastic deformation. The viscosity of the material is inversely proportional to the flow rate and directly proportional to the strength of the network bonds (Mark & Tobolsky, 1950).
The similarity of the potassium-sodium results for fresh and frozen tissue indicates that the network is fairly stable and does not depend upon the immediate activity of cells to maintain structural integrity. The use of frozen tissue in the other tests further precludes the mediation of cellular processes. The sizeable increase in viscosity result-ing from the complete withdrawal of ions by distilled water, in combination with the slight decrease relative to sea water produced by the potassium-sodium solutions, indicates the existence of ionic cross-links which can be shielded by monovalent cations in the aqueous environment. The increase in viscosity effected by calcium and mag-nesium argues for cross-link facilitation by divalent cations, and the synergism of some magnesium-sodium pairs indicates that the ratio of divalent to monovalent species may play an important role in any mechanism based on this effect.
The results of these experiments on holothurians fit a pattern observed in other classes of echinoderms. The inter-vertebral ligaments of ophiuroids lose structural stability in the absence of cations (Wilkie, 1978a, b). Juxtaligamental cells which in-vest and penetrate the tissue may be able to control the mechanical state by regulating the composition of interstitial water (Wilkie, 1979). Paradoxically the catch apparatus of the sea urchin spine (Takahashi, 1967 a, b)' locks' in the presence of high concentra-tions of potassium, but this may be the result of indirect action mediated by the cells. A substance which causes both body wall and catch apparatus to stiffen has recently been discovered in the coelomic fluid of sea cucumbers (Motokawa, 1981).
In the light of the above evidence, it is proposed that echinoderms generally possess the ability to alter the mechanical properties of connective tissues by active adjustment of interstitial water chemistry. These tissues exist in closed compartments bounded by living cells which can serve as active or passive barriers to the surrounding medium. In principle, regulation is not confined to rheological properties as the elasticity of the network can be affected by intra chain chemical reactions as well. Because of the close relationship between elastic and osmotic forces it is possible that volume changes can also occur allowing the system to do work, a possibility already demonstrated in several synthetic polymer systems (Kuhn, Ramel, & Walters, i960; Mandelkern, 1964). The existence of such mechanisms would unravel the mystery of still other anomalies in echinoderm dynamics, such as the contraction of collagenous ligaments in crinoids (Meyer, 1971) and the flexible rigidity of starfish (Eylers, 1976a).
The fact that echinoderms are exclusively marine has long intrigued investigators (Binyon, 1972). It is possible that the reduction of external salinity may set up a gradient which the overlying cells are powerless to pump against and the structural
8 JOHN P. EYLERS
integrity of the animals is severely compromised. Thus the physiological cause for thd fresh water barrier may reside in the mechanism of matrix control (Eylers, 1976 ft).
This work was made possible by grants from Cocos Foundation Inc., Indianapolis, Indiana, and Sigma Xi, The Scientific Research Society, New Haven, Connecticut.
REFERENCES
ALEXANDER, R. M C N . (1962). Visco-elastic properties of the body wall of sea anemones. J. exp. Biol. 39. 373-386.
BINYON, J. (197a). Phytiology of EMnoderms. Oxford: Pergamon
BUECHE, F. (1962). Physical Properties of Polymers. New York: John Wiley.
CASSARO, C. M. F. & DIETRICH, C. P. (1977). Distribution of sulphated mucopolysaccharides in invertebrates. J. biol. Chem. 252, 2254-2261.
ELDER, H. Y. (1973). Distribution and function of elastic fibers in the invertebrates. Biol. Bull. mar.
biol. Lab., Woods Hole 144, 43-63.
EYLERS, J. P. (1976 a). Aspects of skeletal mechanics of the starfish Asterias forbesii. J. Morph. 149, 353-368.
EYLERS, J. P. (19766). Mechanical properties of holothurian body wall. Thalassia Jugoslav. 12, m - 1 1 4 . FREINKEL, W. D. & HEPBURN, H. R. (1975). Dermal stiffness and collagen cross-linking in a sea
cucumber (Holothuroidea). S. Afr. J. Sci. 71, 280-281.
HERMANS, P. H. (1949). Gels. In Colloid Science (ed. H. R. Kruyt). New York: Elsevier. JACOBS, M. H. (1967). Diffusion Processes. New York: Springer-Verlag.
KOEHL, M. A. R. (1977). Mechanical diversity of connective tissue of the body wall of sea anemones.
J. exp. Biol. 69, 107-125.
KOIZUMI, T. (1935). Studies on the exchange and equilibrium of water and electrolytes in a holo-thurian, Caudina chilensis (J. Mliller). Ill (a). On the velocity of permeation of K, Na, Ca, and Mg through the isolated body wall of Caudina. Sci. Rep. Tdhoku Imp. Univ. ser. 4, 10, 269-275.
KUHN, W., RAMEL, A. & WALTERS, D. H. (i960). Conversion of chemical into mechanical energy by homogeneous and cross-striated polymeric systems. In Size and Shape Changes of Contractile
Polymers, (ed. A. Wasserman). Oxford: Pergamon.
MANDELKERN, L. (1964). Contractile processes in fibrous macromolecules. A. Rev. Phys. Chem. 15, 421-448.
MARK, H. & TOBOLSKY, A. V. (1950). Physical Chemistry of High Polymer Systems, vol. II. New York: Interecience.
MEYER, D. L. (1971). The collagenous nature of problematical ligaments in crinoids (Echinodermata).
Mar. Biol. 9, 235-241.
MORTENSEN, T. (1927). Handbook of EMnoderms of the British Isles. London: Oxford University Press. MOTOHIRO, T. (i960). Studies of mucoprotein in marine products. I I : isolation of poly-fucose sulphate
from the mucoprotein in Cucumaria japonica. Bull. Jap. Soc. scient. Fish. a6, 1175-1178.
MOTOKAWA, T. (1981). The stiffness change of holothurian dermis caused by chemical and electrical stimulation. Comp. Biochem. Physiol. 70 C, 41-48.
Mow, V. C. & MANSOUR, J. M. (1977). The nonlinear interaction between cartilage deformation and interstitial fluid flow. Biomechanics io, 31-39.
Mow, V. C. & LAI, W. M. (1979). Mechanics of animal joints. A. Rev. Fluid Mech. 11, 247-288.
STOTT, R. S. H., HEPBURN, H. R., JOFFB, I. & HEFFRON, J. J. A. (1974). The mechanical defensive
mechanism of sea cucumbers. S. Afr. J. Sci. 70, 46-48.
TAKAHASHI, K. (1967 a). The catch apparatus of the sea urchin spine. I. Gross histology. J. Fac. Sci.
Tokyo Univ. 11, 109-120.
TAKAHASHI, K. (19676). The catch apparatus of the sea urchin spine. II. Responses to stimuli. J. Fac.
Sci. Tokyo Univ. 11, 121-130.
TANFORD, C. (1961). Physical Chemistry of Macromolecules. New York: John Wiley.
WAINWRIGHT, S. A., BIGGS, W. D., CURREY, J. D. & GOSLINE, J. M. (1976). Mechanical Design in Organisms. New York: John Wiley & Sons.
WILKIE, I. C. (1978a). Arm autotomy in brittlestars (Echinodermata: Ophiuroidea). J. Zool.i Land.
186, 311-330.
WILKIE, I. C. (19786). Nervously mediated changes in the mechanical properties of a brittlestar ligament. Mar. Behav. Physiol. 5, 289-306.
WILKIE, I. C. (1979). The juxtaligamental cells of Ophiocomina nigra (Abildgaard) (Echinodermata: Ophiuroidea) and their possible role in mechano-effector function of collagenous tissue. Cell Tissue