Human skeletal muscle disuse atrophy: effects on muscle protein synthesis, breakdown and insulin resistance a qualitative review
Full text
Figure
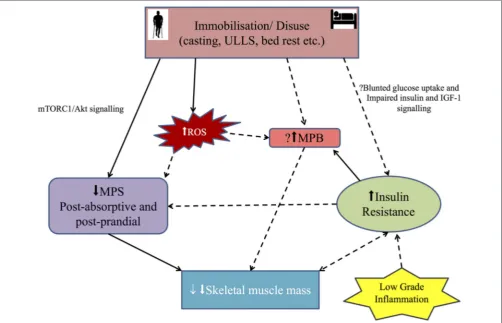
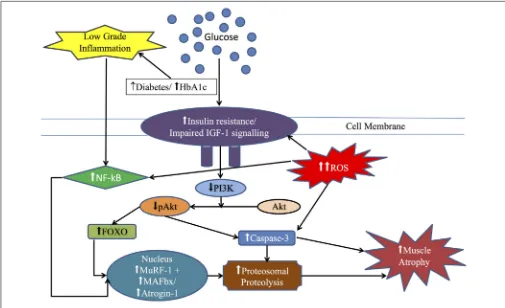
Related documents
Conclusion: Our data suggest that besides the high variety of different detected species, initial antibiotic treatment with a combination of systemic cefuroxime and
Among them are: political will to developing STI as shown by the government ’ s invest- ment in the MSI program and the Presidential Support to Scientists Fund; the presence of
However, for the medium and longer term, income-enhancing export diversification will require that at the center stage of the GoG’s diversification agenda are emerging champions
Yi people are good at singing and dancing in Shilin Yi Autonomous County, the Yi folk song and dancing are the essence of Yi culture, which convey the daily living conditions of
Key words: Technical Efficiency; Stochastic Frontier Production Function; Unorganised Manufacturing Sector; Kerala.. JEL Classification:
Early Childhood Education: Society and Culture provides a comprehensive overview of research from various socio-cultural perspectives enabling meaningful understandings of
In this paper, we have carried out an analysis of discrete-time queueing system with two heterogeneous servers subject to catastrophes for the early arrival system and the late
and Chouhan, S., (2011), Proving fixed point theorems using general principles in cone Banach spaces, Int.. and Rhoades, B.E., (2009), Fixed and periodic point results in cone
