RESEARCH ARTICLE
Configural learning: a higher form of learning in
Lymnaea
Cayley Swinton, Erin Swinton, Tamila Shymansky, Emily Hughes, Jack Zhang, Cailin Rothwell, Mili Kakadiya and Ken Lukowiak*
ABSTRACT
Events typically occur in a specific context and the ability to assign importance to this occurrence plays a significant role in memory formation and recall. When the scent of a crayfish predator (CE) is encountered in Lymnaea stagnalis strains known to be predator experienced (e.g. the W-strain), enhancement of memory formation and depression of feeding occur, which are part of a suite of anti-predator behaviours. We hypothesized thatLymnaeapossess a form of higher-order conditioning, namely configural learning. We tested this by simultaneously exposing W-strainLymnaeato a carrot food odour (CO) and predator scent (CE). Two hours later, we operantly conditioned these snails with a single 0.5 h training session in CO to determine whether training in CO results in long-term memory (LTM) formation. A series of control experiments followed and demonstrated that only the CO+CE snails trained in CO had acquired enhanced memory-forming ability. Additionally, following CE+CO pairing, CO no longer elicited an increased feeding response. Hence, snails have the ability to undergo configural learning. Following configural learning, CO becomes a risk signal and evokes behavioural responses phenotypically similar to those elicited by exposure to CE.
KEY WORDS: Higher-order learning,Lymnaea, Operant
conditioning, Anti-predator behaviour, Predator–prey interactions
INTRODUCTION
Learning, especially associative learning and the subsequent formation of memory, is a vital survival mechanism enabling organisms to generate internal representations of correlations between relevant environmental stimuli. The ability to assign importance to salient stimuli in the environment allows organisms to resolve ambiguity and respond purposefully to their environment (Maren, et al., 2013). The context in which stress is experienced becomes intrinsic to the representation of the experience and influences how and under what circumstances the memory is recalled (de Kloet et al., 1999; Joëls et al., 2006). Here, we tested the
hypothesis thatLymnaeaare capable of a form of‘higher-order’
learning; namely, they possess the cognitive capability for configural learning. We define cognitive capability hereafter Dyer (2012) as the capacity to learn and process information in a sophisticated manner not predicted by elemental responses to stimuli. Hence, the meaning of the stimulus depends on what other stimuli are encountered with it (i.e. the relationships between the sensory components). We define this relationship as configural
learning, using the term as defined by Sutherland and Rudy in 1989.
That is, the meaning of a stimulus depends on what other stimuli are experienced with it (see also Rudy and Sutherland, 1989; Stout et al., 2018). Configural learning results in the ability to treat stimuli experienced together as different from the simple sum of their elements (Giurfa, 2003, 2007; Mansur et al., 2018). We believe that configural learning is vitally important and essential for survival to
Lymnaea as it, along with instinct, occurs in the snails’ natural
environment, which is both changing and composed of multimodal
stimuli, and it is these‘stimuli compounds’that animals associate
events with rather than a single unimodal element (see Lorenz, 1951).
There are distinct populations or strains ofLymnaea(Dodd et al.,
2018) that can be categorized, for example, according to their cognitive or behaviour responses to specific predators (Rothwell et al., 2018). As regards their crayfish predator, there are both
predator-experienced and predator-naive strains ofLymnaea(Orr
et al., 2009a). Here, we define predator-experiencedLymnaeaas
those strains that innately respond with anti-predator behaviours when they detect chemical signals (kairomones) that convey that a crayfish predator is present. However, to be predator experienced need not mean that the snail has actually detected a crayfish at any time in its life. Thus, our W-strain snails, which have been in the laboratory since the 1950s, respond to crayfish kairomones even though they have never actually experienced a crayfish in almost 70 years. Hence, we define a strain as predator experienced if snails respond to the chemical cues with anti-predator behaviours (Burks and Lodge, 2002; Orr et al., 2007; Il-Han et al., 2010). For
example, when predator-experienced Lymnaea such as our
W-strain detect crayfish, a shell-crushing predator, via crayfish effluent (CE), escape behaviours including crawling above the water (Rigby and Jokela, 2000; Dalesman et al., 2006) and increased shadow-elicited withdrawal responses occur (Orr et al., 2007). Importantly, exposing these snails to CE enhances
long-term memory (LTM) formation inLymnaea(Orr and Lukowiak,
2008). Typically, snails will detect CE in a specific stimulus context. We hypothesized that predator-experienced snails will form a connection between an environmental stimulus (e.g. a food odour) and CE, such that subsequent food odour exposure will trigger behaviours typically evoked by CE on its own. These behaviours include suppression of feeding and enhancement of memory formation. A strain is considered to be predator naive if snails of that strain (e.g. TC1 snails) do not respond to CE or other cues transmitted by crayfish with anti-predator behaviours (Orr et al., 2009a).
Here, we used an elemental form of associative learning, operant conditioning of aerial respiratory behaviour. In the inbred
predator-experienced W-strain Lymnaea used, LTM formation requires
at least two 0.5 h training sessions separated by a 1 h interval (Lukowiak et al., 1996, 1998). In this strain, a single 0.5 h training session does not cause LTM formation (Sangha et al., 2003; Forest et al., 2016). However, if trained in CE, a single 0.5 h training
Received 8 August 2018; Accepted 3 January 2019
Hotchkiss Brain Institute, Cumming School of Medicine, University of Calgary, Calgary, AB, Canada T2N 4N1.
*Author for correspondence ([email protected])
K.L., 0000-0001-9028-1931
Journal
of
Experimental
session is sufficient to cause LTM to form (Orr and Lukowiak,
2008; Sunada et al., 2010; Il-Han et al., 2010; Forest et al., 2016).
Hence, training in CE promotes enhanced LTM formation. However, if snails are exposed to CE but not immediately trained (i.e. more than a 0.5 h wait), training in normal pond water (PW) does not result in enhanced LTM formation (Orr and Lukowiak, 2008, 2010). Therefore, CE enhancement of LTM formation is an activity-dependent (i.e. training) phenomenon. In the W-strain, specific stressors applied before or during training alter LTM
formation inLymnaea, either enhancing or obstructing it (Martens
et al., 2006; Dalesman et al., 2013; Lukowiak et al., 2014; Knezevic et al., 2016; Hughes et al., 2017).
Manipulating the context in which a snail undergoes associative
learning alters the snail’s ability to retrieve memory (Haney and
Lukowiak, 2001; Lukowiak et al., 2007). We define context as the surrounding environmental conditions in which a learning task occurs, including stimuli such as food odours. Haney and Lukowiak (2001) demonstrated that carrot odour (CO) can be used to create context-specific memories, and this technique has been implemented successfully in other publications from our laboratory (e.g. Lukowiak et al., 2007; Hughes et al., 2016, 2017). Context learning has been demonstrated in invertebrate model systems
including Aplysia, honeybees, Drosophila and ants, and in
mammalian preparations (Waddell and Quinn, 2001; Giurfa, 2003; Leonard and Edstrom, 2004; Vianna et al., 2004; Bos et al., 2010). Typically, animals trained in one context only showed memory if the animal was tested for memory in the same context. However, context-specific memory cannot be applied across all
conditions. For example, predator-experiencedLymnaeatrained in
CE demonstrate context generalization. To wit, following training in CE, memory is recalled in multiple contexts (e.g. PW or CO) despite the animal being capable of discriminating between the stimuli (Orr and Lukowiak, 2008). This stimulus generalization provides an animal with an adaptive advantage because it allows the animal to generalize the context of a predator encounter to different environmental conditions.
Elemental learning or‘lower-order’forms of associative learning
(i.e. classical and operant conditioning) have been shown in
Lymnaea(Ito et al., 2013). Additionally,Lymnaeaconsolidate these
different forms of associative learning into LTM (Benjamin et al., 2000; Lukowiak et al., 2003; Fulton et al., 2005; McComb et al., 2005a,b; Parvez et al., 2005, 2006; Sakakibara et al., 2005). Forms of higher-order conditioning are conditional learning, sensory pre-conditioning, configural learning and second-order conditioning (Giurfa, 2007). However, we have yet to perform experiments to determine whether forms of higher-order associative learning such as second-order conditioning or configural learning occur with
operant conditioning in Lymnaea. We have previously shown
second-order classical conditioning using our taste aversion
procedure in Lymnaea (Sugai et al., 2006); thus, we were
optimistic that configural learning was possible. Higher-order
conditioning is considered to be more‘cognitive’, as the animal’s
behaviour depends on its ability to make meaningful comparisons between current sensory stimuli and its representation of previous sensory experiences (Sahley et al., 1981; Hawkins et al., 1998; Giurfa, 2003, 2007; Devaud et al., 2015; Onuma and Sakai, 2016).
We hypothesized that configural learning inLymnaeawill occur
when predator-experiencedLymnaea simultaneously encounter a
food odour (e.g. CO) and CE. Here, we show that when
predator-experiencedLymnaeaare exposed to a stimulus that does not elicit
anti-predator behaviours, such as CO, at the same time as they experience CE (i.e. CO+CE), CO now elicits behaviours previously
elicited by the predator odour. These snails trained in CO show enhanced LTM formation and decreased rasping in response to CO.
MATERIALS AND METHODS Snails and animal maintenance
The animals used here are an inbred laboratory strain (W-strain) of
Lymnaea stagnalis(Linnaeus 1758) maintained at the University of
Calgary Biology Department. W-strain snails respond innately and appropriately to CE; thus, they are classified as predator experienced even though they have never experienced a crayfish predator. The
W-strain L. stagnalis used here originated from an inbred stock
maintained at the Vrije University of Amsterdam and were originally bred from animals collected in the 1950s in polders near Utrecht, The Netherlands. The W-strain snails are housed in
artificial PW (0.25 g l−1 of Instant Ocean in deionized water,
Spectrum Brands, Madison, WI, USA) supplemented with CaCO3
to ensure calcium concentrations remain above 50 mg (Dalesman
and Lukowiak, 2010). Snails were fed romaine lettucead libitum
and maintained at 20±1°C on a 16 h:8 h light:dark cycle. A total of 163 snails were used, and a snail was not used more than once in any experiment.
Crayfish effluent
Crayfish are a natural predator ofLymnaea. Predator-experienced
Lymnaeainnately detect and respond to the scent of a crayfish (CE)
with several anti-predator behaviours (Alexander and Covich, 1991;
Dalesman et al., 2006; Orr et al., 2007; Rigby and Jokela, 2000).
The crayfish used here were housed in a 70 l aquarium containing artificial pond water and were fed a diet of lettuce and snails. The water in the crayfish tank is termed crayfish effluent (CE; Orr et al., 2007). Previously, we have demonstrated that exposing
predator-experienced Lymnaea to CE during training enhances LTM
formation (Orr and Lukowiak, 2008, 2010; Lukowiak et al.,
2014). It should be noted here that some strains ofLymnaea(e.g.
TC1 snails collected from a pond in Alberta) are predator naive as regards crayfish (Orr et al., 2009a,b). That is, they do not respond with anti-predator behaviours to crayfish. Here, we examined whether simultaneously exposing snails to CE plus a food odorant (CO, see below; CO+CE) causes configural learning, such that the CO will elicit anti-predator behaviours in predator-experienced
Lymnaea.
Carrot odour
Lymnaeaexhibit context-specific learning, memory formation and
recall, where snails operantly conditioned in CO during training behave as if they are naive (i.e. no LTM) when tested in PW (i.e. no CO; Haney and Lukowiak, 2001; Lukowiak et al., 2007). The CO slurry was made by combining a single large commercially obtained carrot in a blender along with approximately 350 ml of PW. Following blending and straining of the mixture, we obtained
a carrot–PW slurry without any observable pieces of carrot (i.e.
CO). Thus, we used CO as a chemosensory cue in our experiments. We exposed snails to CO by setting up an apparatus that bubbles
either eumoxic air or hypoxic N2 gas through a sealed flask
containing the carrot slurry. The gas bubbled into the flask smelling of carrot was then diverted from the flask into a beaker containing PW. Snails were exposed to CO either under eumoxic conditions prior to training or under hypoxic conditions during conditioning of aerial respiration. It needs to be explicitly stated that multiple batches of fresh carrots purchased from different
grocery stores were used over a 6 week period for Figs 1–6 and a
4 month period for Fig. 7.
Journal
of
Experimental
Operant conditioning of aerial respiratory behaviour and test for memory
Operant conditioning training occurred in three hypoxic contexts: (1) CO; (2) CE; or (3) PW. Individually labelled snails were transferred from their home aquarium into a 1 l beaker filled with 500 ml of one of the three contexts. The specific training environment was made
hypoxic (≤0.1 ml O2l−1) by vigorously bubbling N2gas through the
water for 20 min prior to the operant conditioning session. This is the procedure we have used since our initial study in 1996 (Lukowiak et al., 1996). The hypoxic environment caused snails to attempt to open their pneumostome more frequently. Following this 20 min period, the intensity of bubbling was reduced and the snails to be trained were placed in the beaker for a 10 min acclimation period before training began. The reduced bubbling maintained the established hypoxia without disturbing the snails. The operant conditioning procedure consisted of applying a tactile stimulus (i.e. a poke) to the edge of the pneumostome with a wooden applicator every time a snail attempted to perform aerial respiration. The stimulus was strong enough to cause the pneumostome to close, but was gentle enough that the snails did not complete a full body withdrawal response. The total number of pokes per snail was recorded. Between sessions, snails were returned to their eumoxic home aquarium. In all cases, the memory test (MT) session was performed in hypoxic PW. As in the training session, snails received a tactile stimulus each time they attempted to open their pneumostome in the MT. LTM has been operantly defined as significantly fewer attempted pneumostome openings in the MT session than in the training session. In the W-strain, the standard operant conditioning procedure requires two 0.5 h training sessions (TS) separated by 1 h to cause formation of a 24 h LTM; however, enhanced LTM formation occurs with a single 0.5 h TS in this strain when trained in the presence of certain stressors (e.g. CE-predator detection), as well as when trained in food stuffs containing epicatechin (Lukowiak et al., 2000, 2014; Knezevic et al., 2016; Swinton et al., 2018).
The CO+CE simultaneous exposure
We placed a naive cohort of 10–12 W-strain snails in a beaker
containing 500 ml of CE, made eumoxic by bubbling CO atmospheric air into the beaker. Thus, snails simultaneously experienced CO+CE.
The snails were left in this beaker undisturbed for 45 min and subsequently transferred back to their home aquarium (i.e. neither CO nor CE present), where they remained for 2 h. The snails were then trained with a single 0.5 h training session in hypoxic CO using the operant conditioning of aerial respiration procedure as described above (Fig. 1). In all experimental and control groups (see below), MT was performed 24 h later in PW (i.e. neither CO nor CE was present in the MT session).
Control pairing procedure
We first exposed a naive cohort of snails to CO alone for a 45 min period. That is, they did not experience CE, only CO. These snails were then trained as described above in hypoxic CO (Fig. 2).
Second,we trained a naive cohort of snails that had been exposed to
only CE for 45 min. That is, they did not experience CO. These snails were then trained as described above in hypoxic CO (Fig. 3). Third, we paired CO with boiled CE and exposed a naive cohort of snails to this for 45 min. Boiling alters the composition of the CE (e.g. metabolites in the urine; see Poulin et al., 2018) such that snails no longer respond to it with anti-predator behaviours (Orr et al., 2007). These snails were then trained as described above in hypoxic CO (Fig. 4). Finally, we exposed a naive cohort of snails to CO+CE for 45 min (Fig. 5) or 10 min (Fig. 6). However, these snails were trained in hypoxic PW (i.e. neither CO nor CE present).
Rasping behaviour
Feeding behaviour or rasping in Lymnaea is a rhythmic motor
behaviour in which repeated movements of the radulae scrape the surface of a substrate, leading to the ingestion of food (Ito et al., 2013). In this experiment, snails were placed into a 14 cm diameter Petri dish with enough normal eumoxic PW or eumoxic CO-PW for the snails to be fully submerged. The snails were given a 10 min acclimation period in each session (i.e. PW and CO-PW) before their rasping behaviour was monitored. Each snail was monitored for 2 min and the number of rasps counted; the average number of rasps per minute was then calculated. In Fig. 7A, snails were exposed to CO only, on two occasions 24 h apart; during this period, snails were not fed. In Fig. 7B, snails were first monitored in PW, returned to their home aquarium for 3 h, then tested in CO-PW.
0 10 20 30
No. of attempted pneumostome openings
TS MT
CO+CE exposure period
(45 min)
[image:3.612.47.361.58.301.2]2 h Train snails 24 h Memory test
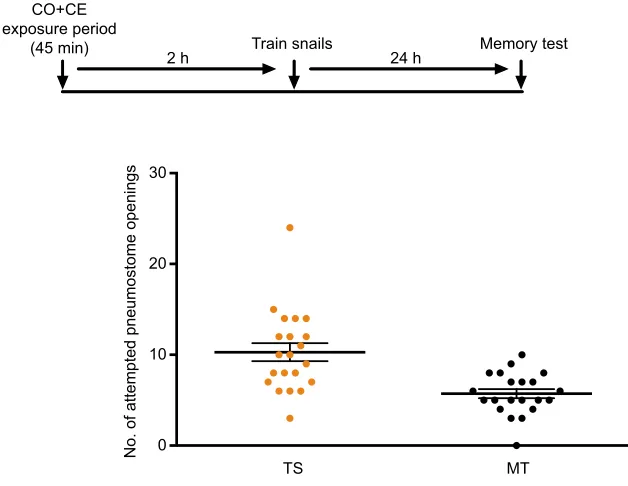
Fig. 1. Pairing of carrot odour and crayfish effluent (CO+CE) before training produces memory enhancement when snails are trained in CO alone.A timeline of the experiment is presented above the data. Untrained W-strainLymnaea(n=21) were simultaneously exposed to CO+CE for 45 min, 2 h before being trained (TS) in CO alone (orange circles) and were tested for memory (MT) 24 h later in pond water (PW; black circles). These snails exhibited long-term memory (LTM) as the number of attempted pneumostome openings in MT was significantly fewer than that in TS (P<0.01). The solid line is the mean and the error bars are the s.e.m.
Journal
of
Experimental
Snails were returned to their home aquarium for 24 h, and were not fed during this interval. Then, rasping behaviour was again determined in these snails in PW. Three hours later they were exposed to CO+CE for 45 min. Following this pairing they were returned to their home aquarium for 3 h before rasping behaviour was again determined in CO-PW.
Data analysis
Using Prism 7 for Mac, a two-way repeated-measures analysis of variance (RM-ANOVA) determined whether there was a statistical difference in the number of attempted pneumostome openings in
each training and memory test session for the data in Figs 1–5. A
post hoc test (Sidak’s multiple comparisons test) was then
performed to determine whether there was a statistical difference between TS and MT for each cohort of snails subjected to the
various conditions (i.e. paired CO+CE, etc.). We chose Sidak’s test
over the more popular Bonferroni test because Sidak’s test has more
power than the Bonferronipost hoctest, as recommended by Prism.
A one-way RM-ANOVA followed by Tukey’spost hoctests was
employed to compare whether there were significant differences in
rasping behaviour. In Fig. 7A, a pairedt-test was used. Differences
were considered significant ifP-values were smaller than 0.05.
RESULTS
We hypothesized that Lymnaeaare capable of a form of
higher-order learning known as configural learning. If our hypothesis is correct, W-strain snails will learn to associate the simultaneous of a
food smell (CO) with the ‘smell’ of a predator (i.e. CE). The
statistical analyses of data (a two-way ANOVA followed by Sidak’s
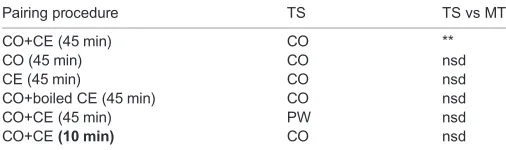
test) from the experiments performed to test this hypothesis are presented here and in Table 1. We report the following for the data in
Figs 1–6: (1) there was a significant interaction effect (F5,258=2.669;
0 10 20 30 40
No. of attempted pneumostome openings
TS MT
CO exposure period
(45 min)
[image:4.612.46.366.60.302.2]2 h Train snails 24 h Memory test
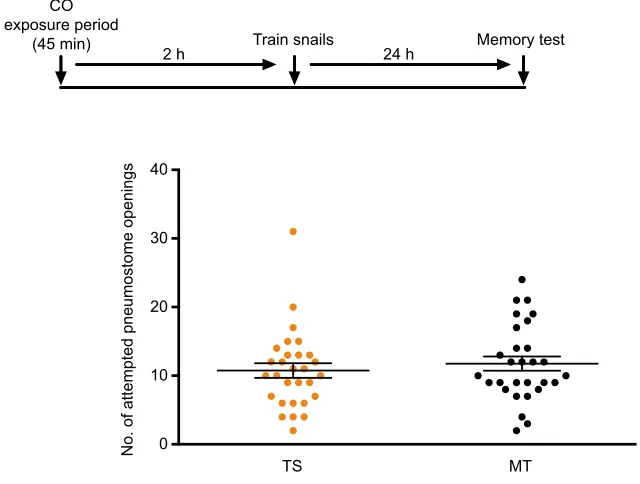
Fig. 2. Exposure to only CO for 45 min, 2 h before training in CO, does not result in memory
enhancement.A timeline of the experiment is presented above the data. Training in CO (orange circles) after exposure to CO in this W-strainLymnaeacohort (n=29) did not result in LTM formation in PW (black circles). The solid line is the mean and the error bars are the s.e.m.
0 10 20 30
No. of attempted pneumostome openings
TS MT
CE exposure period
(45 min)
[image:4.612.47.357.503.738.2]2 h Train snails 24 h Memory test
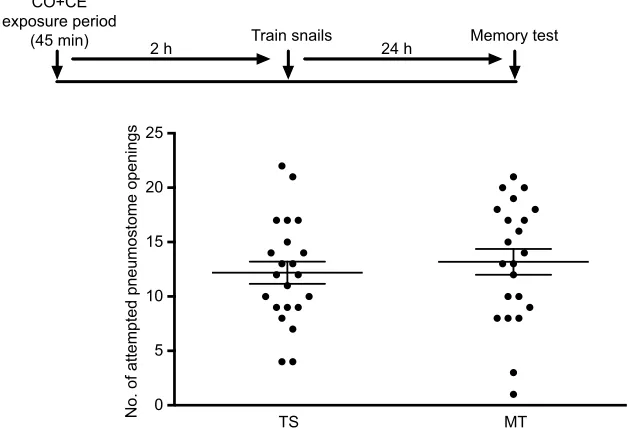
Fig. 3. Exposure to only CE for 45 min, 2 h before training in CO, does not result in memory
enhancement.A timeline of the experiment is presented above the data. This cohort of W-strainLymnaea(n=21) also did not exhibit LTM following exposure to CE and training in CO (orange circles) when tested 24 h later in PW (black circles). The solid line is the mean and the error bars are the s.e.m.
Journal
of
Experimental
P=0.0226); (2) there was also a significant ‘pairing effect’ (i.e. comparing the data from CO+CE with those from CO only, etc.:
F5,258=10.70;P<0.0001); and (3) there was no significant TS and
MT effect (i.e. TS in CO+CE versus CO only, etc.:F1,258=0.8450;
P=0.3588). The post hocSidak’s multiple comparisons tests are
presented in Table 1 and show that the only significant difference between TS and MT was in the CO+CE pairing group when snails were then trained in CO. This was the only cohort to exhibit LTM.
We first placed a cohort of untrained W-strain snails (Fig. 1,n=21)
into a beaker containing eumoxic CE-PW (CE) and CO. Snails experienced this procedure for 45 min (i.e. paired CO+CE) and were then returned to their home eumoxic aquaria (important to note, neither CE nor CO was present there). Two hours later, this cohort of snails was trained with a single 0.5 h TS in hypoxic CO and then tested 24 h later (MT) in hypoxic PW (CO not present). There were significantly fewer attempted pneumostome openings in the MT than in the TS
(P<0.01, determined by Sidak’s post hoc test). Thus, after the
simultaneous exposure to CO+CE for 45 min, when trained in CO,
snails exhibited an enhanced ability to form LTM in PW. Importantly, note that the TS occurred in CO 2 h after the simultaneous exposure to CO+CE, whilst the MT session 24 h later occurred in PW.
We next performed a number of experiments to determine whether it was both necessary and sufficient to experience the CO+CE pairing for CO to cause enhancement of LTM formation. First, we exposed an
untrained cohort of snails (n=29) to CO alone for 45 min and then,
following a 2 h rest, trained them in hypoxic CO (Fig. 2). These snails did not form LTM in PW. That is, there was no significant difference
(determined by Sidak’spost hoctest) between the number of attempted
pneumostome openings in the MT and TS. We then exposed a cohort
of untrained snails (n=21) to CE only, for 45 min, and then trained
them 2 h later in hypoxic CO (Fig. 3). As can be seen, LTM was not formed. That is, the number of attempted pneumostome openings in the MT was not significantly lower than the number in the TS
(determined by a Sidak’spost hoc test). Thus, in these two control
experiments, training in hypoxic CO was not sufficient to lead to LTM formation when the CO+CE pairing did not occur.
0 5 10 15 20 25
No. of attempted pneumostome openings TS MT
CO+boiled CE exposure period
[image:5.612.47.361.59.288.2](45 min) 2 h Train snails 24 h Memory test
Fig. 4. Simultaneous exposure to CO+boiled CE for 45 min, 2 h before training in CO, does not result in memory enhancement.A timeline of the experiment is presented above the data. Pairing of CO+boiled CE before training in CO in this cohort of W-strainLymnaea(n=22) did not result in LTM formation (MT; black circles) when snails were trained in CO alone (orange circles). The solid line is the mean and the error bars are the s.e.m.
0 5 10 15 20 25
No. of attempted pneumostome openings TS MT
CO+CE exposure period
(45 min) 2 h Train snails 24 h Memory test
Fig. 5. Simultaneous exposure to CO+CE for 45 min, 2 h before training in PW, does not result in memory enhancement.A timeline of the experiment is presented above the data. This cohort of W-strainLymnaea(n=22) when exposed to CO+CE then trained in PW (black circles), rather than in CO as in Fig. 1, did not exhibit LTM formation (MT; black circles). The solid line is the mean and the error bars are the s.e.m.
Journal
of
Experimental
[image:5.612.47.362.521.735.2]It has previously been shown (Orr et al., 2007, 2009b) that the crayfish kairomones that signal predator presence and which elicit
anti-predator behaviours in W-strainLymnaea are inactivated by
boiling the CE. Therefore, we first boiled the CE, let it cool to room temperature, and then added the CO mixture. We then exposed a
cohort of untrained snails (n=22) to a pairing of CO+boiled CE for
0 5 10 15 20
No. of attempted pneumostome openings TS MT
CO+CE exposure period
[image:6.612.50.355.59.284.2]2 h Train snails 24 h Memory test
Fig. 6. Simultaneous exposure to CO+CE for 10 min, 2 h before training in CO, does not result in memory enhancement.A timeline of the experiment is presented above the data. When these W-strainLymnaea(n=22) were trained in CO (orange circles) for 10 min, rather than 45 min, 2 h after exposure to CO+CE, they did not exhibit LTM when tested in PW (black circles) 24 h later. The solid line is the mean and the error bars are the s.e.m.
0 5 10 15 20 25
PW CO PW CO
3 h 24 h 3 h
Rasps in PW 3 h
Rasps in CO Rasps in PW Rasps in CO CO+CE
exposure period (45 min)
0 min 3 h 24 h
0 5 10 15 20
Rasping behaviour (rasps min
–1
)
CO CO
24 h
Rasps in CO Rasps in CO 24 h
A
B
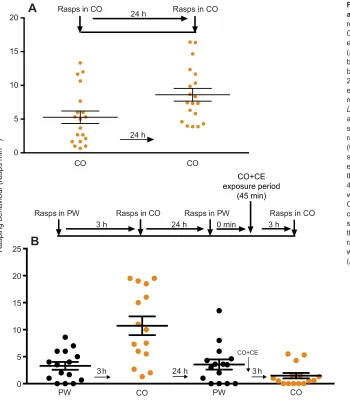
CO+CE
Fig. 7. Rasping behaviour and CO before and after configural learning.(A) The rasping response in W-strainLymnaeawas calculated for CO. Twenty-four hours later, the snails were again exposed to CO. There was a statistically significant (P<0.001) increase in the number of rasps elicited by the second exposure to CO, most likely because the snails had not been fed during the 24 h interval between the first and second exposure to CO. (B) The spontaneous feeding response in an untrained cohort of W-strain Lymnaeawas calculated for PW (black circles) and 3 h later for CO (orange circles). There was a significant increase (P<0.001) in the number of rasps per minute in CO. Twenty-four hours later (the snails were not fed during this period), the spontaneous feeding response was again examined in PW (black circles). Immediately after this, the snails were exposed to CO+CE for 45 min. Three hours later, their feeding response was again tested in CO. Following the exposure to CO+CE, CO did not increase rasping. The number of rasps in the second presentation of CO was not significantly different from that in PW. However, there was a significant decrease in the number of rasps in the second exposure to CO compared with the response to the first exposure to CO (P<0.001).
Journal
of
Experimental
[image:6.612.55.405.339.740.2]45 min. When these snails were trained 2 h later in hypoxic CO, they did not exhibit LTM (Fig. 4), as there was no significant difference between the number of attempted openings in the MT and
TS (determined by Sidak’spost-hoctest).
We next determined whether training snails in CO subsequent to the CO+CE pairing is necessary to cause enhanced LTM formation.
We simultaneously exposed a cohort of untrained snails (n=22) to
CO+CE for 45 min and then 2 h later we trained these snails (Fig. 5) in PW (not CO as we did in Fig. 1). We tested memory 24 h later in
PW and found that LTM was not present (determined by Sidak’s
post hoctest).
The final experiment examining LTM formation that we performed was to simultaneously expose a cohort of untrained
snails (n=22) to CO+CE for 10 min rather than 45 min (Fig. 6). We
then trained these snails in CO as in Fig. 1 and tested memory 24 h later in PW. We found that LTM was not present in the MT, as there
was no significant reduction (determined by Sidak’spost hoctest) in
the number of attempted pneumostome openings from the TS to the MT. That is, a 10 min simultaneous exposure to CO+CE was not sufficient for CO to cause enhancement of LTM.
Next, we asked whether configural learning (i.e. the simultaneous exposure for 45 min to CO+CE) also alters the feeding response of the snails. As part of the suite of anti-predator behaviours elicited in
predator-experienced Lymnaea by predator detection (e.g. CE),
feeding behaviour (rasping) is significantly reduced (Dalesman et al., 2006; Orr et al., 2007). We first determined that the feeding response elicited by CO was not significantly reduced with two repeated
presentations of CO. Thus, an untrained cohort of snails (n=19) was
exposed to CO and then 24 h later exposed to CO a second time. These data are presented in Fig. 7A. It is important to note here that in the 24 h interval between the first and second exposure to CO, snails were not fed. As can be seen, rasping behaviour was not significantly decreased on the second exposure to CO; in fact, it significantly
increased ( paired t-test; t=6.801, d.f.=18; P<0.0001). We then
performed a similar experiment using another cohort of untrained
snails (n=26). These data are presented in Fig. 7B. The number of
rasps in these snails was observed in PW and CO before and after CO
+CE pairing. An ANOVA (F3,100=22.80; P<0.0001) followed by
Tukey’spost hoctest indicated that there were significant differences
in the feeding responses in this experiment. The rate of rasping seen in
PW is typical for Lymnaea (Sugai et al., 2006; Ito et al., 2013).
Following the initial observation of rasping in PW, we returned the snails to their home aquarium for 3 h. These snails were subsequently exposed to CO and their rasping behaviour was observed. As can be seen, the number of rasps increased significantly compared with the
response in PW (P<0.001). Following this, the snails were returned to
their home aquarium for 24 h and, as in Fig. 7A, were not fed. They
were again observed in PW. The number of rasps in their second exposure to PW was not different from the number of rasps observed in their first exposure, 24 h previously. Immediately after their second exposure to PW, the snails experienced the CO+CE pairing for 45 min and were then returned to their home aquarium. We then tested their response to CO 3 h later. The number of rasps elicited by
this exposure to CO decreased significantly (P<0.001) compared with
their initial response to CO (i.e. the response before the CO+CE pairing). The number of rasps elicited by CO was now not significantly different from the rasping behaviour seen in PW. Thus, following the CO+CE pairing, CO no longer elicited a significant increase in the feeding response.
DISCUSSION
We show here that predator-experienced W-strainLymnaeapossess
features of‘higher-order’learning as they have the ability to associate
a food odour (CO), which inherently elicits a feeding response, with the scent of a predator (i.e. CE) when they simultaneously experience CO+CE. During this simultaneous exposure, the snails freely behave and no other external stimuli are applied. The experiencing of the two stimuli together leads to CO, which previously did not elicit anti-predator behaviours, to acquire the properties of the anti-predator kairomone (i.e. CE) in eliciting anti-predator behaviours (i.e. enhanced memory-forming ability and suppression of feeding behaviour). We classify this new association between CO and CE as configural learning as defined by Sutherland and Rudy (1989).
That is, snails have the ability to assign a new‘meaning’to the odour
stimulus (CO) as a result of experiencing CO simultaneously in the presence of kairomones from crayfish (CE) that evoke anti-predator responses. We believe that configural learning is of great importance
to the snail as it may be an essential form of learning in the snails’
natural environment. This new learning (i.e. CO signalling anti-predator behaviour) persists for at least 2 h. We hypothesize snails
come to associate the CO with a ‘motivational state’ that is
instinctively elicited by CE in W-strain snails. Consequently, CO
after pairing now elicits a‘fear state’as a result of the association
made by the snails between CO and CE. A component of this fear state, the suppression of the feeding response, becomes elicited by
CO (Fig. 7B). CO now signals‘fear’rather than eliciting an appetitive
feeding response. CO, therefore, has acquired a new motivational state (i.e. fear) as opposed to its intrinsic motivational state (i.e. enhanced rasping). This change in stimulus signalling did not occur when CO was paired with boiled CE, as boiling CE inactivates its predator-signalling properties (Orr et al., 2007). Configural learning also did not occur with a 10 min simultaneous exposure to CO+CE. Finally, enhanced LTM formation did not happen if, after the simultaneous 45 min pairing of CO+CE, the snails were trained in PW and not CO.
To our knowledge in an invertebrate model system, configural learning has only previously been shown in bees (Giurfa, 2003, 2007, Dyer, 2012; Devaud et al., 2015). For example, bees possess
the ability to acquire olfactory patterning discrimination.
Interestingly, this ability is blocked when neural activity in the mushroom bodies is blocked, even though elemental learning still
occurs. Other ‘higher forms’ of learning such as second-order
conditioning and contingency learning have been shown in a
number of molluscs includingLimax(Sahley et al., 1981, 1990),
Hermissenda(Farley, 1987), Aplysia (Hawkins et al., 1998) and
Lymnaea(Sugai et al., 2006). The second-order conditioning seen
in Sugai et al.’s (2006)Lymnaeastudy utilized a conditioned taste
[image:7.612.47.300.100.175.2]aversion (CTA) procedure (i.e. classical conditioning), which is different from the operant conditioning procedure used here. We
Table 1. Results of a two-way ANOVA (F5,258=2.669;P=0.0226) followed
by Sidak’s multiple comparisonspost hoctest with‘pairing’type as a
between-subjects factor, and‘training/memory tests’as the
within-subjects factor
Pairing procedure TS TS vs MT
CO+CE (45 min) CO **
CO (45 min) CO nsd
CE (45 min) CO nsd
CO+boiled CE (45 min) CO nsd
CO+CE (45 min) PW nsd
CO+CE(10 min) CO nsd
CO, carrot; PW, pond water; CE, crayfish effluent; TS, training session; MT, memory test. All MT sessions were in PW. Five of the six TS were in CO, while the 6th was in PW. **P<0.01; nsd, not significantly different.
Journal
of
Experimental
direct interested readers to the paper by Pearce (2002) on configural
versus more ‘elemental’ forms of learning. Thus, our finding of
higher-order learning here is not too surprising. What is different in
our study is that we showed thatLymnaeaare capable of configural
learning using an operant conditioning procedure. That is, snails acquired a new association by experiencing a carrot odour, which typically only elicits feeding behaviour (Sugai et al., 2006), simultaneously with a crayfish kairomone (CE), which elicits a suite of anti-predator behaviours, including enhanced memory formation and suppression of feeding (Orr et al., 2009a,b). The simultaneous encountering of CO+CE leads to CO subsequently acquiring an ability on its own to elicit anti-predator behaviours rather than eliciting an increase in feeding. We believe this is the initial instance of configural learning in a mollusc.
One of the stress factors that animals experience in their natural environment leading to alterations in behaviour, physiology and even anatomy (e.g. shell thickness or shape) is predator detection (Rundle and Brönmark, 2001; Orr et al., 2007; Herberholz et al., 2004; Stokes et al., 2004). When animals are confronted with predation risk, they often exhibit purposive behavioural responses that may help them survive (Dalesman et al., 2006; Orr et al., 2007; Rigby and Jokela, 2000). Learning about predators and under what circumstances a predator threat may occur is an important adaptation and will produce alterations in behaviour during times of predation risk (Coolen et al., 2005). Additionally, the ability to make associations to the circumstances under which predation risks or actual events are experienced plays an important role in eliciting appropriate responses to similar circumstances in the future.
Stress, in the biological context, is defined as any circumstance or
condition that disrupts the physiological or psychological
homeostasis of an organism (Kim and Diamond, 2002). Stress itself is a physiological mechanism that helps organisms take on challenges. Animals are exposed to many stressful survival challenges in their natural environments. Our results show that simultaneously experiencing a predator threat with an odour changes the behavioural response and probably the neuronal response to that odour (Funk and Amir, 2000). Our results also allow us to hypothesize that the neurons responsible for LTM formation (e.g. RPeD1; Scheibenstock et al., 2002) will become responsive to an odour that has now gained emotional or ecological significance as a result of being experienced with a kairomone that signals danger. Experiments will be performed in the future to test this hypothesis.
Our results are consistent with the hypothesis that stimulus–stimulus
learning is an important adaptive learning mechanism that helps animals decipher the meaning of important stimuli in their environment. From previous experiments, we know that exposure to CE results in short-lasting changes in the electrophysiological properties of RPeD1, and that combining CE exposure with training is necessary for long-lasting changes to occur (Orr and Lukowiak, 2008; Orr et al., 2009b). The perception of CE primes the molecular mechanisms required for LTM formation (Parvez, et al., 2006) and we believe that after configural learning used here, CO does the same.
We believe that the context in which predator detection, or for that matter any significant stressful event, is processed in the nervous system will enable the organism to predictively respond to the context
as if it were the stressor. Whether this is adaptive in all cases–for
example, it may be maladaptive in post-traumatic stress disorder
(PTSD)–remains to be determined. However, in the example we
tested here, experiencing predator threat with a non-threatening odour (i.e. CO+CE) shows that the context in which a stressor is presented will signal a response that is phenotypically similar to that following presentation of the stressor alone. Further investigation is required to
determine whether the causal neuronal mechanisms underlying enhanced memory formation are similar to those demonstrated when the animal is trained with the CE stressor alone.
In summary, predator-experienced W-strainLymnaeaassociate
the chemosensory CO with the scent of a predator (CE) by just being simultaneously exposed to the two stimuli. This simultaneous exposure results in CO acquiring a new ability to elicit anti-predator behaviours. Our results show that configural learning and memory
formation occur in Lymnaea, indicating that the snails not only
possess the ability to consider the individual components of stimuli (i.e. CO and CE) but also can form a relationship between the two stimuli. Additionally, the two stimuli are treated by the snail as different from the simple sum of the stimuli alone.
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: C.S., K.L.; Methodology: C.S., E.S., J.Z., K.L.; Validation: J.Z.; Formal analysis: K.L.; Investigation: C.S., E.S., T.S., E.H., J.Z., C.R., M.K.; Resources: K.L.; Data curation: K.L.; Writing - original draft: K.L.; Writing - review & editing: E.S., K.L.; Supervision: K.L.; Project administration: K.L.; Funding acquisition: K.L.
Funding
Funding was provided by the National Science and Engineering Research Council of Canada (NSERC) to K.L. C.S. was supported by a Summer Studentship from Alberta Innovates - Health Solutions; C.R. was supported by a post-doctoral grant from the Alberta Innovates - Health Solutions.
References
Alexander, J. E., Jr and Covich, A. P.(1991). Predation risk and avoidance behavior in two freshwater snails.Biol. Bull.180, 387-393.
Benjamin, P. R., Staras, K. and Kemenes, G.(2000). A systems approach to the cellular analysis of associative learning in the pond snailLymnaea.Learn. Mem.7, 124-131.
Bos, N., Guerrieri, F. J. and d’Ettorre, P. (2010). Significance of chemical recognition cues is context dependent in ants.Anim. Behav.80, 839-844.
Burks, R. L. and Lodge, D. M.(2002). Cued in: advances and opportunities in freshwater chemical ecology.J. Chem. Ecol.28, 1901-1917.
Coolen, I., Dangles, O. and Casas, J.(2005). Social learning in noncolonial insects?Curr. Biol.15, 1931-1935.
Dalesman, S. and Lukowiak, K. (2010). Effect of acute exposure to low environmental calcium on respiration and locomotion inLymnaea stagnalis(L.). J. Exp. Biol.213, 1471-1476.
Dalesman, S., Rundle, S. D., Coleman, R. A. and Cotton, P. A.(2006). Cue association and antipredator behaviour in a pulmonate snail,Lymnaea stagnalis. Anim. Behav.71, 789-797.
Dalesman, S., Sunada, H., Teskey, M. L. and Lukowiak, K.(2013). Combining stressors that individually impede long-term memory blocks all memory processes.PLoS ONE8, e79561.
de Kloet, E. R., Oitzl, M. S. and Joëls, M.(1999). Stress and cognition: are corticosteroids good or bad guys?Trends Neurosci.22, 422-426.
Devaud, J.-M., Papouin, T., Carcaud, J., Sandoz, J.-C., Grunewald, B. and Giurfa, M. (2015). Neural substrate for higher-order learning in an insect: mushroom bodies are necessary for configural discriminations.Proc. Natl Acad. Sci. USA112, E5854-E5862.
Dodd, S., Rothwell, C. and Lukowiak, . (2018). Otherness, crowding, and long-term memory formation inLymnaea.Comp. Biochem. Physiol. A222, 43-51.
Dyer, A. G.(2012). The mysterious cognitive abilities of bees: why models of visual processing need to consider experience and individual differences in animal performance.J. Exp. Biol.215, 387-395.
Farley, J.(1987). Contingency learning and causal detection inHermissenda: I. Behavior.Behav. Neurosci.101, 13-27.
Forest, J., Sunada, H., Dodd, S. and Lukowiak, K.(2016). TrainingLymnaeain the presence of a predator scent results in a lasting ability to form enhanced long-term memory.J. Comp. Physiol A202, 399-409.
Fulton, D., Kemenes, I., Andrew, R. J. and Benjamin, P. R.(2005). A single time-window for protein synthesis-dependent long-term memory formation after one-trial appetitive conditioning.Eur. J. Neurosci.21, 1347-1358.
Funk, D. and Amir, S.(2000). EnhancedFosexpression within the primary olfactory and limbic pathways induced by an aversive conditioned odor stimulus. Neuroscience98, 403-406.
Journal
of
Experimental
Giurfa, M.(2003). Cognitive neuroethology: dissecting non-elemental learning in a honeybee brain.Curr. Opin. Neurobiol.13, 726-735.
Giurfa, M.(2007). Behavioral and neural analysis of associative learning in the honeybee: a taste from the magic well.J. Comp. Physiol. A193, 801-824.
Haney, J. and Lukowiak, K.(2001). Context learning and the effect of context on memory retrieval inLymnaea.Learn. Mem.8, 35-43.
Hawkins, R. D., Greene, W. and Kandel, E. R.(1998). Classical conditioning, differential conditioning, and second-order conditioning of the Aplysia gill-withdrawal reflex in a simplified mantle organ preparation. Behav. Neurosci.
112, 636-645.
Herberholz, J., Sen, M. M. and Edwards, D. H.(2004). Escape behavior and escape circuit activation in juvenile crayfish during prey-predator interactions. J. Exp. Biol.207, 1855-1863.
Hughes, E., Shymansky, T., Sunada, H. and Lukowiak, K.(2016). Qualitatively different memory states in Lymnaea as shown by differential responses to propranolol.Neuro Learn Mem.136, 63-73.
Hughes, E., Shymansky, T., Swinton, E., Lukowiak, K. S., Swinton, C., Sunada, H., Protheroe, A., Phillips, I. and Lukowiak, K. (2017). Strain-specific differences of the effects of stress on memory in Lymnaea.J. Exp. Biol.220, 891-899.
Il-Han, J.-I., Janes, T. and Lukowiak, K.(2010). The role of serotonin in the enhancement of Long-term memory resulting from predator detection in Lymnaea.J. Exp. Biol.213, 3603-3614.
Ito, E., Kojima, S., Lukowiak, K. and Sakakibara, M.(2013). From likes to dislikes: conditioned taste aversion in the great pond snail Lymnaea stagnalis. Can. J. Zool.91, 405-412.
Joëls, M., Pu, Z., Wiegert, O., Oitzl, M. S. and Krugers, H. J.(2006). Learning under stress: how does it work?Trends. Cogn. Sci.10, 152-158.
Kim, J. J. and Diamond, D. M.(2002). The stressed hippocampus, synaptic plasticity and lost memories.Nat. Rev. Neurosci.3, 453-462.
Knezevic, B., de Freitas, E., Komatsuzaki, Y. and Lukowiak, K. (2016). A flavanoid component of chocolate quickly reverses an imposed memory deficit. J. Exp. Biol219, 816-823.
Leonard, J. L. and Edstrom, J. P.(2004). Parallel processing in an identified neural circuit: theAplysia californicagill-withdrawal response model system.Biol. Rev. Camb. Philos. Soc.79, 1-59.
Lorenz, K.(1951). The role of Gestalt perception in animal and human behavior. In Aspects of Form(ed. L. L. Whyte), pp. 157-178. London: Lund Humphries.
Lukowiak, K., Ringseis, E., Spencer, G., Wildering, W. and Syed, N.(1996). Operant conditioning of aerial respiratory behaviour inLymnaea stagnalis.J. Exp. Biol.199, 683-691.
Lukowiak, K., Cotter, R., Westly, J., Ringseis, E. and Spencer, G.(1998). Long-term memory of an operantly conditioned respiratory behaviour pattern in Lymnaea stagnalis.J. Exp. Biol.201, 877-882.
Lukowiak, K., Adatia, N., Krygier, D. and Syed, N.(2000). Operant conditioning in Lymnaea: evidence for intermediate- and long-term memory. Learn. Mem.7, 140-150.
Lukowiak, K., Sangha, S., Scheibenstock, A., Parvez, K., McComb, C., Rosenegger, D., Varshney, N. and Sadamoto, H.(2003). A molluscan model system in the search for the engram.J. Physiol.97, 69-76.
Lukowiak, K., Fras, M., Smyth, K., Wong, C. and Hittel, K. (2007). Reconsolidation and memory infidelity inLymnaea.Neurobiol. Learn. Mem.87, 547-560.
Lukowiak, K., Sunada, H., Teskey, M., Lukowiak, K. and Dalesman, S.(2014). Environmentally relevant stressors alter memory formation in the pond snail Lymnaea.J. Exp. Biol.217, 76-83.
Mansur, B. E., Rodrigues, J. R. V. and Mota, T.(2018). Bimodal patterning Discrimination in harnessed honey bees.Front. Psychol.9, 1529.
Maren, S., Phan, K. L. and Liberzon, I.(2013). The contextual brain: implications for fear conditioning, extinction and psychopathology.Nat. Rev. Neurosci.14, 417-428.
Martens, K. R., de Caigny, P., Parvez, K., Amarell, M., Wong, C. and Lukowiak, K.(2006). Stressful stimuli modulate memory formation inLymnaea stagnalis. Neurobiol. Learn Mem.87, 391-403.
McComb, C., Rosenegger, D., Varshney, N., Kwok, H. Y. and Lukowiak, K.
(2005a). Operant conditioning of an in vitro CNS-pneumostome preparation of Lymnaea.Neurobiol. Learn. Mem.84, 9-24.
McComb, C., Varshney, N. and Lukowiak, K.(2005b). JuvenileLymnaeaventilate, learn and remember differently than do adult Lymnaea. J. Exp. Biol. 208, 1459-1146.
Onuma, T. and Sakai, N.(2016). Higher-order conditioning of taste-odor learning in rats: evidence for the association between emotional aspects of gustatory information and olfactory information.Physiol. Behav.164, 407-416.
Orr, M. V. and Lukowiak, K.(2008). Electrophysiological and behavioral evidence demonstrating that predator detection alters adaptive behaviors in the snail Lymnaea.J. Neurosci.28, 2726-2734.
Orr, M. and Lukowiak, K.(2010). Sympatric predator detection alters cutaneous respiration in Lymnaea.Commun. Integr. Biol.3, 42-45.
Orr, M. V., El-Bekai, M., Lui, M., Watson, K. and Lukowiak, K.(2007). Predator detection inLymnaea stagnalis.J. Exp. Biol.210, 4150-4158.
Orr, M., Hittel, K., Lukowiak, K. S. Han, J. and Lukowiak, K.(2009a). Differences in LTM-forming capability between geographically different strains of Alberta Lymnaea stagnalisare maintained whether they are trained in the lab or in the wild.J. Exp. Biol.212, 3911-3918.
Orr, M. V., Hittel, K. and Lukowiak, K.(2009b).‘Different strokes for different folks’: geographically isolated strains ofLymnaea stagnalisonly respond to sympatric predators and have different memory forming capabilities.J. Exp. Biol. 212, 2237-2247.
Parvez, K., Stewart, O., Sangha, S. and Lukowiak, K. (2005). Boosting intermediate-term into long-term memory.J. Exp. Biol.208, 1525-1536.
Parvez, K., Rosenegger, D., Martens, K., Orr, M. and Lukowiak, K.(2006). Learning at a snail’s pace.Can. J. Neurol. Sci.33, 347-356.
Pearce, J. M.(2002). Evaluation and development of a connectionist theory of configural learning.Anim. Learn Behav.30, 73-95.
Poulin, R. X., Lavoie, S., Siegel, K., Gaul, D. A., Weissburg, M. J. and Kubanek, J.(2018). Chemical encoding of risk perception and predator detection among estuarine invertebrates.Proc. Natl. Acad. Sci. USA115, 662-667.
Rigby, M. C. and Jokela, J.(2000). Predator avoidance and immune defence: costs and trade-offs in snails.Proc. Biol. Sci.267, 171-176.
Rothwell, C. M., Spencer, G. E. and Lukowiak, K.(2018). The effect of rearing environment on memory formation.J. Exp. Biol.221, 408-413.
Rudy, J. W. and Sutherland, R. J.(1989). The hippocampal formation is necessary for rats to learn and remember configural discriminations.Behav. Brain Res.34, 97-109.
Rundle, S. D. and Brönmark, C.(2001). Inter- and intraspecific trait compensation of defence mechanisms in freshwater snails.Proc. Biol. Sci.268, 1463-1468.
Sahley, C., Rudy, J. W. and Gelperin, A.(1981). An analysis of associative learning in a terrestrial mollusc: I. Higher-order conditioning, blocking, and a transient US pre-exposure effect.J. Comp. Physiol.144, 1-8.
Sahley, C., Martin, K. and Gelperin, A.(1990). Analysis of associative learning in the terrestrial molluscLimax maximus. II. Appetitive learning.J. Comp. Physiol.
167, 339-345.
Sakakibara, M., Aritaka, T., Iizuka, A., Suzuki, H., Horikoshi, T. and Lukowiak, K.
(2005). Electrophysiological responses to light of neurons in the eye and statocyst ofLymnaea stagnalis.J. Neurophysiol.93, 493-507.
Sangha, S., Morrow, R., Smyth, K., Cooke, R. and Lukowiak, K.(2003). Cooling blocks ITM and LTM formation and preserves memory.Neurobiol. Learn Mem.80, 130-139.
Scheibenstock, A., Krygier, D., Haque, Z., Syed, N. and Lukowiak, K.(2002). The Soma of RPeD1 must be present for long-term memory formation of associative learning inLymnaea.J. Neurophysiol.88, 1584-1591.
Stokes, V. L., Pech, R. P., Banks, P. B. and Arthur, A. D.(2004). Foraging behaviour and habitat use by Antechinus flavipesand Sminthopsis murina (Marsupialia: Dasyuridae) in response to predation risk in eucalypt woodland. Biol. Conserv.117, 331-342.
Stout, D. M., Glenn, D. E., Acheson, D. T., Spadoni, A. D., Risbrough, V. B. and Simmons, A. N.(2018). Neural measures associated with configural threat acquisition.Neurobiol. Learn. Mem.150, 99-106.
Sugai, R., Shiga, H., Azami, S., Watanabe, T., Sadamoto, H., Fujito, Y., Lukowiak, K. and Ito, E.(2006). Taste discrimination in conditioned taste aversion of the pond snail Lymnaea stagnalis.J. Exp. Biol.209, 826-833.
Sunada, H., Horikoshi, T., Lukowiak, K. and Sakakibara, M.(2010). Increase in excitability of RPeD11 results in memory enhancement of juvenile and adult Lymnaea stagnalis by predator-induced stress. Neurobiol. Learn Mem. 94, 269-277.
Sutherland, R. and Rudy, J.(1989). Configural association theory: the contribution of the hippocampus co learning, memory, and amnesia.Psychobiology 17, 129-144.
Swinton, E., de Freitas, E., Swinton, C., Shymansky, T., Hiles, E., Zhang, J., Rothwell, C. and Lukowiak, K.(2018). Green tea and cocoa enhance cognition inLymnaea.Commun. Integr. Biol.11, e1434390.
Vianna, M. R., Coitinho, A. S. and Izquierdo, I.(2004). Role of the hippocampus and amygdala in the extinction of fear-motivated learning.Curr. Neurovasc. Res.
1, 55-60.
Waddell, S. and Quinn, W. G.(2001). What can we teachDrosophila? What can they teach us?Trends. Genet.17, 719-726.