10
The Role of Public Chemical
Information in Antipredator
Behaviour
Brian D. Wisenden
1and Douglas P. Chivers
21Biosciences Department, Minnesota State University Moorhead, Moorhead, MN, 56563, USA.
e-mail: [email protected]
2Biology Department, University of Saskatchewan, Saskatoon, SK, S7N 5E2, Canada.
e-mail: [email protected]
ABSTRACT
Predators stalk, attack, capture and ingest prey. At each stage, chemical cues are released passively as a natural by-product of this behavioural interaction. These chemical cues are not signals in the strict sense because their production and release are not shaped by natural selection by benefits to the sender that accrue from receiver responses. However, predation exerts steep selection on receivers to detect and respond adaptively to these cues, because doing so reduces the receiver’s risk of predation. Here, we briefly discuss disturbance cues, predator odours, alarm cues and dietary cues released during predation. We then consider more complex behavioural ecological manifestations of public chemical information, including interactions between chemical and visual indicators of predation risk, and associations of risk with novel stimuli following exposure to chemical alarm cues.
Key Words: Chemical cues; Predator–prey interactions; Public information; Antipredator behaviour
1. INTRODUCTION
behaviourists, who, as humans, have sensory receptors sensitive to visual and acoustic modalities. However, a substantial literature now exists in the subfield of chemical communication, a world largely outside of everyday human experience. In particular, terrestrial arthropods have been well studied, and instead of studying light spectra and acoustic ranges, chemical ecologists discuss molecules, mixtures ratios, and the manner in which molecular side groups stimulate olfactory receptors.
Communication can be defined generally as an exchange of information between a signaller and a receiver. Signallers gain fitness benefits by manipulating receivers, and receivers gain fitness benefits from the information provided by the signaller. In this manner, the response of the receiver applies selection pressure on the signaller to increase the efficacy and efficiency of information transfer. Concomitantly, signallers create selection upon receivers to maximize detection of information in the signal. Communication theory was developed to explain the evolution of signals, the role of receptors in shaping signal properties such as information content, signal reliability and honesty (Bradbury and Vehrencamp, 1998).
In this chapter, we step back from this classic model of communication because most information in the environment is not an intentional signal issued by a signaller that benefits from the response of receivers (see Wisenden and Stacey, 2005, for discussion of the evolution of chemical signals). Fishes are surrounded by an ongoing chemical narrative describing biological interactions over space and time. The fitness rewards for the ability to detect and respond adaptively to chemical information steeply promote specialised receptors and behavioural templates triggered by these environmental chemical cues.
Public information benefits the receiver but unlike true communication, it does not necessarily benefit the sender. Consequently, we need not seek specialised signals in this type of system. Public information does, however, exert selection to promote the elaboration of specialised receptors in receivers for selective information gathering. Receptor proteins and sensory neurons trigger higher brain centres that result in behavioural responses to chemical cues. Chemical ecologists can test the selectivity, sensitivity thresholds and cognitive mapping of receptors by studying behavioural responses to various chemical stimuli.
Predator-prey interactions produce an abundance of chemical information. It is useful to parse predation events into component parts because different stages of predation release different types of chemical cues (Lima and Dill, 1990; Smith, 1992; Figure 10.1, Table 10.1) that indicate the imminence and magnitude of predation risk, which in turn, select different behavioural responses (Chivers and Smith, 1998).
Fig. 10.1: Overview of the predation sequence and types of cues released. Predators release a natural odour, called a kairomone, used by the prey to detects its presence. Chemical cues associated with the prey in the digestive tract of the predator, called dietary cues, are released through the anus. Any prey injured during capture releases chemical alarm cues that cause antipredator behaviour in conspecifics, heterospecifics in the same prey guild (cross-species reaction) and attract competing predators. Disturbed, but uninjured prey release disturbance cues.
then they can avoid an attack by avoiding detection. Several types of chemical cues give the prey advance warning of the presence of a predator.
The first type of chemical cue is the odour of the predator itself (review, Kats and Dill, 1998). Every species has a chemical signature that is passively released as a natural by-product of normal metabolic processes. These cues, known as kairomones, are defined as predator odours that alert prey to a predator’s presence. Moreover, kairomones also inform the prey of the species
Table 10.1: Classification of chemical cues released at each stage of the predation sequence.
Predation Predator cues Prey cues Receiver Threat Behavioural
stage level response
Detection Kairomones Disturbance Prey Low Inspection predator
Dietary cues cues Increase vigilance
Attack and Kairomones Alarm cues Prey High Flee
capture Dietary cues Disturbance Other Increase shoal
cues? predator cohesion
Reduce activity Seek shelter Increase vigilance Recognition learning Dietary
cues
Alarm cues
Cross-species response to alarm cues
Disturbance cues Kairomones
Predator attractant
o o
of predator and the degree of threat this predatory species represents (Kusch et al., 2004). The second type of chemical cue is the odour of disturbed prey. These cues are released when prey are startled or stressed, but not injured. The third type of chemical cue is damage-released alarm cues (review Chivers and Smith, 1998). These chemicals, released when a prey is injured by a predatory attack, indicate not only the presence of a predator, but one that is actively foraging, and one that is actively foraging on conspecific prey. Thus, injury-released cues usually evoke intense behavioural responses.
2. SAMPLING BEHAVIOURS
The nose of most fishes is a pair of small invaginated pouches on the snout lined with olfactory receptors and connected to the external environment by one, or usually two, openings (see Hamdani and Døving, Chap. 9, this volume). Ciliary action passes water over the olfactory epithelium as a fish swims through the water. When predation risk is high, active movement is reduced because movement attracts the attention of predators. Thus, at a time when the adaptive value of chemical information is greatest, the risk of sampling from large volumes of the water column is also greatest. Fish have at least two behavioural mechanisms for chemically sampling the water around them that do not involve conspicuous movement. The first is fin-flicking (Keenleyside, 1955; Lawrence and Smith, 1989; Brown et al., 1999). Here, fish maintain a stationary position in the water column and rapidly open their medial and pectoral fins, which push the fish backward slightly, while giving an equal and opposite push forward with the caudal fin so as to maintain stationary status with minimal loss of crypsis. Water currents created by the pectoral and dorsal fins waft past the external nares and provide better access to chemical public information.
3. ALARM CUES
Chemical alarm cues are passively released into the public realm by damaged tissue when a predator attacks and captures a prey (Chivers and Smith, 1998). Animals able to detect and respond to these cues can avoid the most imminent class of predation threat. So ancient and steep is this selection pressure that antipredator responses to conspecific alarm cues is virtually universal among aquatic animals. Protozoans, flatworms, annelids, arthropods, molluscs, fishes and amphibians, all exhibit alarm reactions to conspecific chemical alarm cues (Wisenden, 2003).
Fig. 10.2: Chemical cues are distributed unequally throughout the water column for water in flow. In panel 1, a dye is released upstream of a stack of drinking straws lying on their side, producing stable laminar flow. In panels 2 and 3, resistance to flow from the surface, and substrate, retard advancement of the odour plume. For benthic fishes such as darters, elevated head posture during predator vigilance extends olfactory receptors into the leading edge of the plume. The cartoon fish on the downstream end of panel 3 indicate the advantage of head-up chemical sampling behaviour in providing earlier detection and continual updates of important chemical information. Panel 4 shows a johnny darter (Etheostoma nigrum) in normal head-down feeding posture. Panel 5 shows the same fish with its head partially raised. In full head-up posture, the snout points almost straight up and the pectoral fins are fully extended. Photographs taken by Oscar Noriega (Noriega and Wisenden, unpub. data).
1
2
3
o
o
4
5
Among fishes, members of the superorder Ostariophysi have received the most attention (Von Frisch, 1941; Pfeiffer, 1977; Smith, 1992; Chivers and Smith, 1998). The Ostariophysi is speciose and includes Gonorhynchiformes, carps and minnows (Cypriniformes), characins and tetras (Characiformes), catfishes (Siluriformes) and knife fishes (Gymnotiformes). Many of these are small schooling species that fall prey to a range of predators. Ostariophysans possess specialized epidermal cells that are easily ruptured upon the slightest damage to the skin. The cell contents are known as Schreckstoff (Von Frisch 1938) or ‘alarm substance’. While the alarm substance may have other functions—including an anti-parasite/anti-pathogen role (Chivers et al., unpub. data)—the alarm substance is highly effective in inducing antipredator behaviour. The active space generated by the release of alarm cues is quite large. Lawrence and Smith (1989) reported that one cm2 of minnow skin
produces 58,000 litres of active space, equivalent to a sphere with a radius of 3.2 m. Similar results have been found in other fishes (Mirza and Chivers, 2003a; Dupuch et al., 2004). Careful quantification of the area of skin damaged during an attack has never been carried out but would presumably depend on the size of the prey, the ratio of prey size to predator size and the hunting style of the predator. Predators only marginally larger than their prey require longer handling times and probably release a greater percentage of the prey’s stored alarm substance (Chivers et al., 1996). Be that as it may, for even a small prey species such a minnow, 1 cm2 of skin is a reasonably conservative estimate
of the amount of cue released and, thus, ecological volume affected by a typical predation event. For larger prey species, the area of skin damaged by a predator attack could be much larger. When prey detect alarm cues, they flee the area, increase shoal cohesion to enhance the selfish herd effect, and in lab aquaria at least, reduce activity and increase the time spent near the bottom (Lawrence and Smith, 1989). These behaviours reduce the probability of detection by the predator, and if detected, reduce the probability of capture (Mathis and Smith, 1993a).
Although ostariophysans are the group for which the behavioural consequences of alarm cue release have been best studied, many other fish groups behave similarly to conspecific skin extract and some groups possess analogous epidermal club cells (Poeciliidae: Garcia et al., 1992; Mirza et al., 2001; Eleotridae: Kristensen and Closs, 2004; Percidae: Smith, 1979, Commens and Mathis, 1999, Mirza et al., 2003; Centrarchidae: Brown and Brennan, 2002; Cottidae: Chivers et al., 2000; Bryer et al., 2003).
4. PREDATOR ODOUR
planktonic crustacea. An important predator in these systems is the dipteran Chaoborus, but several fishes including stickleback and perch have kairomones that affect plankton behaviour and morphology (see Wisenden, 2003 for review).
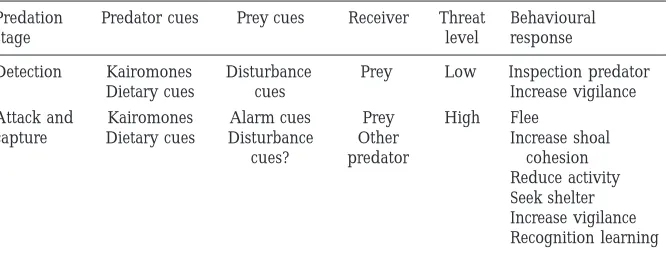
Prey fishes are well tuned to kairomones (Kats and Dill, 1998). One system that has received considerable scientific attention is the effect of kairomones of northern pike (Esox lucius) on the behaviour of small fishes upon which it preys. For example, Mathis et al. (1993) identified population differences in the responses of fathead minnows (Pimephales promelas) to pike odour. Minnows from populations sympatric with pike recognise and respond to pike kairomones, while minnows from populations allopatric from pike do not. Recognition learning can occur very quickly under natural conditions (Chivers and Smith, 1994a). For example, Chivers and Smith (1995) showed that a population of approximately 20,000 minnows in a 1 ha pond acquired recognition of pike odour within 2 weeks of the introduction of 10 juvenile pike to the pond. In a similar study, Brown et al., (1997) showed that a population of approximately 78,000 minnows in a 4 ha pond learned to recognise pike odour within 4 to 6 days following the introduction of 39 juvenile pike. In this case, the learned recognition of the predator odour occurred faster than recognition of the sight of the predator. Kusch et al., (2004) showed that minnows exhibit surprisingly sophisticated responses to pike odour. Minnows exhibit more intense anti-predator responses to increasing concentrations of pike odour (Fig. 10.3). Moreover, minnows are able to recognise the size of the pike that generated the cues. Juvenile pike represent a greater threat to minnows than larger pike and minnows show correspondingly higher intensity responses to cues of small pike than to cues of large pike. Such results indicate that we need to carefully consider what other information may be contained in a predators’ odour. Kairomones have been demonstrated to be important determinants of prey behaviour in many predator-prey systems (Kats and Dill, 1998).
5. DIETARY CUES
(2001) reviewed 79 papers published between 1956 and 2000 that examined the responses of fishes and amphibians to chemical cues from predators. While the majority of studies have not considered predator-diet as a variable, the ones that did had most often identified significant diet-related effects.
The use of diet cues allows prey animals to adjust the intensity of their responses to predators in order to reflect their risk of predation. For example, Mirza and Chivers (2001a) showed that juvenile perch (Perca flavescens) use the presence of conspecifics in the diet of adult conspecifics to determine appropriate response. The intensity of cannibalism is highly variable through time and space, hence juvenile perch should avoid wasting their time and energy responding to adults unless the adults are cannibalistic (Fig. 10.4).
Predator-diet cues have also been directly linked to adaptive changes in body morphology of some fishes. For example, Brönmark and Miner (1992) showed that crucian carp (Carassius carassius) exhibited an adaptive change in morphology (an increase in body depth) in response to pike fed carp but not to cues from pike fed chironomids. Similarly, Stabell and Lwin (1997) documented that carp exhibited an increase in body depth in response to cues of pike fed carp but not pike fed Arctic charr (Salvelinus alpinus). The increase
Fig. 10.3: The proportion of fathead minnows that responded to cues of small and large pike and to a control of distilled water. The proportion of minnows exhibiting a behavioural response to pike increases with increasing concentration of cues. Moreover, the proportion of minnows exhibiting a response to cues of small pike was greater than the proportion responding to cues of large pike. Figure modified from Kusch et al. (2004).
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
30 ml 60 ml 90 ml 120ml
Distilled water
30 ml 60 ml 90 ml 120ml
Small pike
30 ml 60 ml 90 ml 120ml
Large pike
Propor
in body depth results in the carp being less vulnerable to gape-limited predators.
6. DISTURBANCE CUES
Disturbance cues are released by startled or stressed prey. Early work in this area showed that decapod crustaceans responded with vigilant postures when exposed to water that contained disturbed conspecifics (Hazlett, 1985). Moreover, this same water induced predator vigilance in other taxa (e.g. darters), indicating that the disturbance cue was likely a generic metabolic by-product of stress common to many aquatic taxa, such as a pulse release of nitrogenous wastes (Hazlett, 1990; Kiesecker et al., 1999). Studies on fish are limited to darters (Wisenden et al., 1995), characins (Jordao and Volpato, 2000), and salmonids (Mirza and Chivers, 2001b, 2002).
Fig. 10.4: Mean (+ SE) change in shelter use of juvenile yellow perch (Perca flavescens) exposed to chemical cues from adult perch fed on either juvenile perch, spot tail shiners (Notropis hudsonius), swordtails (Xiphophorus helleri) or exposed to distilled water. Juvenile yellow perch exhibited antipredator responses dependent on the diet of the predator. Perch increase shelter use when exposed to cues of adult perch fed conspecifics and perch fed spot tail shiners but not to perch fed swordtails. The response to perch fed shiners represents cross-species responses to members of the same prey guild (see discussion on cross-species reactions for more explanation). Modified from Mirza and Chivers (2001a). Letters above bars denote significant differences at p < 0.05.
Distilled Water
Perch-fed Swordtail
Perch-fed Spot tail
Perch-fed Perch
150
100
50
0
–50
–100
Change in shelter use (s)
7. INTERACTIONS BETWEEN CHEMICAL AND VISUAL
CUES FOR RISK ASSESSMENT
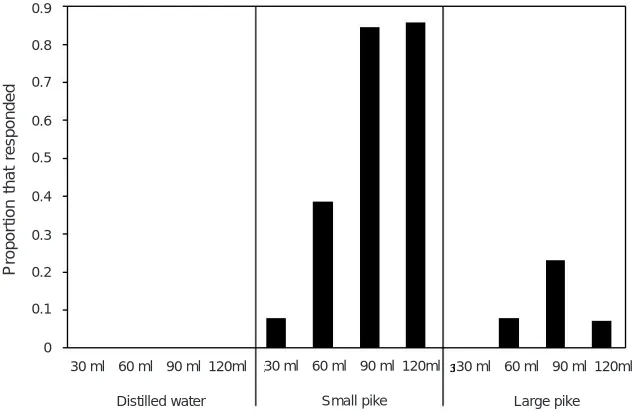
Fish rely on any and all sensory modalities to collect information about their world and adjust their behaviour accordingly. Anything less would be maladaptive. The value and influence of chemical information can be promoted or discounted by visual information, or vice versa. For example, many prey fishes engage in predator inspection behaviour upon visually detecting a predator. A great deal of literature exists on the hypothesized function(s) of inspection behaviour and the conditions under which it should occur (Dugatkin and Godin, 1992). To approach a predator would seem maladaptive, but prey accrue benefits by gaining information about the predator’s identity, the predator’s hunger state, and the predator’s diet. Much of this information is ascertained by olfactory sampling during inspection visits. For example, glowlight tetras (Hemigrammus erythrozonus) that inspect a cichlid predator avoid the area if the predator has eaten tetras recently but not if the predator has eaten swordtails (Xiphophorus helleri) (Brown and Godin, 1999; Fig. 10.5A). Finescale dace (Phoxinus neogaeus) avoid the ‘attack cone’ in front of the head
Fig. 10.5:(A) Glowlight tetras (Hemigrammus erythrozonus) inspect heterospecifics that are potential predators. If, upon inspection, the inspecting fish detect dietary alarm cues of conspecifics, they avoid the area (left pair of bars). Non-inspecting fish do not detect sufficient dietary cues to elicit avoidance behaviour. Asterisk denotes P < 0.001. Figure modified from Brown and Godin (1999).
Tetra Diet
Swordtail Diet
Inspectors Non-inspectors
40
20
0
–20
–40
–60
–80
–100
of a model predator if chemical stimuli of perch on a diet of dace are released. No avoidance of the attack cone occurred when odour of perch on a diet of swordtail or when water blank (control) accompanied the model (Brown et al., 2001; Fig. 10.5B).
Mosquitofish (Gambusia affinis) use chemical and visual cues in an additive manner when inspecting predatory green sunfish (Lepomis cyanellus) (Smith and Belk, 2003). Chemical cues from the sunfish diet and visual cues from changes in sunfish behaviour indicating sunfish hunger status, additively contribute to distance maintained from a sunfish during an inspection visit by mosquitofish. Although additive, chemical and visual cues differ qualitatively in the sense that chemical cues contribute more to distance from the predator, while visual cues contribute more to vertical distribution and inspection behaviour (Smith and Belk, 2003).
Fathead minnows (Pimephales promelas) and redbelly dace (Phoxinus eos) can combine chemical and visual sources of information in a synergistic fashion (Wisenden et al., 2003), meaning that the response to chemical and visual information in combination of was opposite to the response to at least one of the cues on its own. In this field experiment, traps were chemical labelled with minnow alarm cues or water (control) and such traps also differed visually
Fig. 10.5:(B) Wild finescale dace (Phoxinus neogaeus) were presented with a model predator and simultaneously the odour of perch (Perca flavescens) on a diet of finescale dace, swordtails (Xiphophorus helleri), or no diet (food deprived for 96 h). The presence of dietary alarm cues caused singleton dace to strongly orient their inspection behaviour toward the tail and away from the predator’s head, where risk of attack is greatest. Shared letters above bars denote non-significant differences at P < 0.05. Figure modified from tabular data in Brown et al. (2001).
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
Distilled Water Dace Food Deprived Swordtail Before
After
a
a
b
b
Propor
by the presence or absence of a small fish shoal contained within a jar inside the trap (an empty jar was used as a control). Alarm cue was predicted to be aversive and reduce catch numbers while the presence of a shoal was predicted to increase catch numbers because they are obligate shoalers. The lowest catch rate was for the combination of alarm cue + no shoal but the highest catch rate occurred for the combination of alarm cue + shoal. Fish shoal + water had the second highest catch rate and no shoal + water had the second lowest catch rate. Chemical alarm cues seemed to induce area avoidance in the absence of a shoal but caused a strong behavioural proclivity to increase shoal cohesion in the presence of a shoal. The presence of a shoal in the traps induced chemically alarmed fish in the general free-swimming population to shoal with the fish in the jar and thus, enter the traps. Chemical alarm cues caused avoidance when presented alone, but caused super-attraction when combined with the visual stimulus of a shoal. This occurred even though traps were the source of the alarm cue (Wisenden et al., 2003).
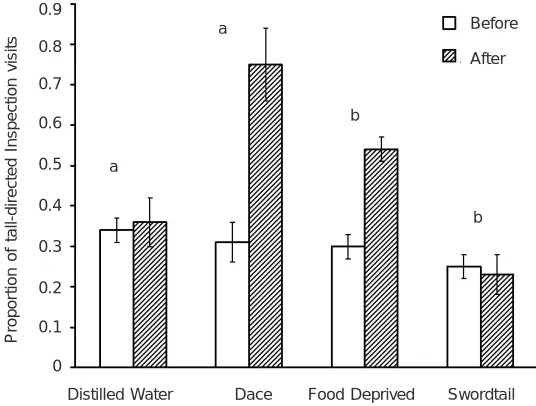
Behavioural responses to visual cues can be tempered by the presence or absence of confirming information from visual cues because the degree of threat, and therefore the best behavioural counter strategy for avoiding attack, depends on the predator species and its proximity to the prey. Thus, if the water is turbid, then confirmation of chemically mediated risk by visual information is reduced or not possible. Under turbid conditions, overt antipredator behaviour is the default response to chemical alarm cues (Hartman and Abrahams, 2000). Conversely, in clear water, chemical alarm cues unaccompanied by confirming visual indicators of risk produce a muted behavioural response to chemical alarm cues. However, if prey have been first exposed to chemical alarm cues, the magnitude of response to a subsequent visual indicators of risk is intensified (Garcia et al., 1992; Brown and Godin, 1999; Wisenden et al., 2004). The inhibitory effect of high water clarity can make it appear as if natural populations of free-swimming fish are unresponsive to chemical alarm cues (Fig. 10.6A), where closer examination shows unequivocal alarm responses (Figs 10.6B and 10.6C).
8. RELEASER-INDUCED RECOGNITION LEARNING
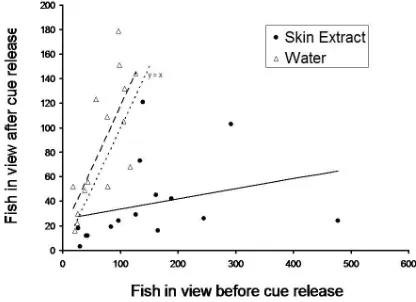
Fig. 10.6: (A) The number of fish in view of an underwater video camera 30 min. before and after the release of minnow (Phoxinus phoxinus) muscle extract (open circles, dotted line) or skin extract (solid circles, solid line). Although the trend was in the predicted direction, the two treatments did not differ statistically. The line labelled y = x indicates the point at which the number of fish, after cue introduction, is equal to the number before cue introduction. Based on tabular data in Magurran et al. (1996).
Fig. 10.6: (B) The number of fish in view of an underwater video camera 5 min. before and 2 min. after the release of skin extract (solid lines, solid circles) of black-nose shiners, or water control (dashed lines, open circles), followed immediately by the approach of a model predator (open and solid triangles for water and alarm cue trials, respectively). The effect of an approaching model predator (after water release) had the same effect as the release of chemical alarm cues. The effect of the model predator after alarm cue release was to further intensify the avoidance response. Based on a figure in Wisenden et al. (2004).
Skin Extract
Muscle Extract 14
12
10
8
6
4
2
0
2 3 4 5 6 7 8 9 10
Number of fish before test cue
Number of fish after test cue
y = x
160
140
120
100
80
60
40
20
0
0 20 40 60 80 100 120 140 160
Number of fish in view (time 1) Model predator (water cue)
Model predator (alarm cue) Alarm cue
with risk. This learning mechanism permits prey the flexibility to track spatial and temporal shifts in predation risk across geographic space and through ontogenetic and ecological time. Unlike many invertebrates and amphibians, most fishes do not have innate recognition of kairomones of their predators (but see Berejikian et al., 2003; Vilhunen and Hirvonen, 2003 for exceptions in some salmonids). Kairomones become associated with risk by experiencing the direct release of alarm cues during a predatory attack, or through dietary cues detected via passive (Mathis and Smith, 1993b) or even active (Brown and Godin, 1999) sampling by predator inspection behaviour.
In a recent study, Ferrari et al. (unpubl. data) showed that fathead minnows learn the identity of a predator associated with alarm cues and also the relative risk associated with that predator. Minnows exposed to cues of brook charr (Salvelinus fontinalis) combined with low concentrations of alarm cues learn to exhibit weak responses to charr cues. In contrast, minnows exposed to cues of brook charr combined with high concentrations of alarm cues learn to project strong responses to charr cues.
What happens if a fish experiences more than one novel stimulus after experiencing alarm cues? When presented with alarm cues and two novel objects—a black disk and a horizontal dark tube—and one object was made to rise up and sink 10 times, fathead minnows associated fear with the moving object, whether the moving object was the tube or the disk (Wisenden and Harter, 2001). The non-moving object, regardless of its shape, was ignored. Presumably, motion is a much more reliable indicator of predator identity than shape. What if the novel stimulus does not arrive simultaneously with
alarm cues? Do fish still associate danger with the novel stimulus? Zebra danios associated predation risk with pike odour when it was introduced 5 min. after the introduction of alarm cues (Korpi and Wisenden, 2001).
9. CROSS-SPECIES REACTIONS
A prey guild is an assemblage of species that share a common predator(s). Alarm cues indicate risk to any prey in the same guild as the species that release the alarm cues. Thus, just as natural selection steeply promotes antipredator behaviour in response to conspecific alarm cues, there is an almost equally steep selection promoting mechanisms for the recognition and response to alarm cues from heterospecific prey within the same prey guild. Cross-species reactions turn out to be quite common. In some cases, they occur by conservation of alarm cue chemistry among closely related species (Mirza and Chivers, 2001c; Brown et al., 2003) but in other instances, they occur by learned associations through a variety of pathways. These associations can form via secondary associations between a known kairomone and fresh release of a novel heterospecific alarm cue, or a known kairomone and dietary release of a novel heterospecific alarm cue, or a kairomone of a predator with a gut releasing a combination of dietary alarm cues of conspecifics and heterospecifics (Mirza and Chivers, 2001d, 2003b; Chivers et al., 2002). Pollock and Chivers (2003a,b) used artificial pond experiments to attempt to address how ecological variables influence the ability of fish to learn to recognize heterospecific alarm cues. They found that an increase in habitat complexity or a decrease in stickleback density reduced the learning of stickleback cues by minnows.
10. SOCIAL TRANSMISSION OF ALARM INFORMATION
11. FUTURE RESEARCH DIRECTIONS
In this chapter, we have outlined the major categories of chemical public information and the manner in which they are used by prey to ameliorate predation risk. Much basic work still needs to be carried out on the role of chemical cues in mediating predator-prey interactions. For example, releaser-induced recognition learning and social transmission of releaser stimuli should lead fishes to fear virtually everything in their environment before too long. This is not the case obviously, but the hierarchical criteria and their cognitive mechanisms for discerning irrelevant stimuli from salient stimuli are not well understood. Future work should be directed to filling this knowledge gap. Likewise, we should aim to determine what information prey acquire when they smell a predator. We know that prey can determine the diet of the predator (e.g. Mathis and Smith, 1993b) and the size of the predator (Kusch et al., 2004). Are there other important sources of information in the predator’s odour? For example, can prey determine the age of the odour? Information about the age of an odour combined with information about movement patterns of predators may allow prey greater flexibility and resolution in avoiding predation. We also need to consider whether different types of predators have the ability to counteract the ability of prey to detect them. Can predators mask their odours? Much of what is known about chemical ecology of fishes is derived from experiments conducted in laboratory aquaria. While this is convenient and productive, there is always the suspicion that lab studies may lack ecological realism (Magurran et al., 1996; Tremaine et al., in press). Chemical ecology is a field that lends itself well to experimental manipulation in the field because, unlike foraging and reproductive behaviours, antipredator behaviour occurs rapidly. Data can be collected within minutes of the test, and replicated many times in the course of a single afternoon.
Interactions between water pollutants on the transportation of chemical cues and the effect of pollutants on olfactory receptors are relatively open questions. Some recent experiments have demonstrated that relatively small changes in pH can have a major impact on responses to chemical alarm cues. For example, Leduc et al. (2004) showed that brook charr respond to alarm cues in streams with a pH of 6.88 but not in streams with a pH of 6.11. Sublethal concentrations of pollutants may have profound behavioural ecological effects for prey populations; thus, indirectly degrading ecosystem health. As global waterways continue to decline in quality, this area of chemical ecology is ever more deserving of close attention.
REFERENCES
Bradbury, J.W. and S.L. Vehrencamp. 1998. Principles of animal communication. Sinauer. Brönmark, C. and L.B. Miner. 1992. Predator-induced phenotypic change in body morphology
in crucian carp. Science 258: 1348–1350.
Brown, G.E. and S. Brennan. 2000. Chemical alarm signals in juvenile green sunfish (Lepomis
cyanellus, Centrarchidae). Copeia 2000: 1079–1082.
BrownG.E. and D.P. Chivers. In press. Learning as an adaptive response to predation. In: Ecology
and Evolution of Predator/Prey Interactions. P. Barbosa, and I. Castellanos (eds). Oxford
University Press, Oxford.
Brown, G.E. and J.-G.J. Godin. 1999. Who dares, learns: chemical inspection behaviour and acquired predator recognition in a characin fish. Anim. Behav. 57: 475–481.
Brown, G.E., D.P. Chivers, and R.J.F. Smith. 1997. Differential learning rates of chemical versus visual cues of a northern pike by fathead minnows in a natural habitat. Environ. Biol. Fishes 49: 89–96.
Brown, G.E., J.-G.J. Godin, and J. Pederson. 1999. Fin flicking: A visual antipredator alarm signal in a characin fish (Hemigrammus erythrozonus). Anim. Behav. 59: 469–476.
Brown, G.E., J.L. Golub, and D. Plata. 2001. Attack cone avoidance during predator inspection visits by wild finescale dace (Phoxinus neogaeus): The effects of predator diet. J. Chem. Ecol. 27: 1657–1666.
Brown, G.E., J.C. Jr. Adrian, N.T. Naderi, M.C. Harvey, and J.C. Kelly. 2003. Nitorgen oxides elicit antipredator responses in juvenile channel catfish, but not in convict cichlids or rainbow trout: conservation of the Ostariophysan alarm pheromone. J. Chem. Ecol. 29: 1781–1796. Bryer, P.J., R.S. Mirza, and D.P. Chivers. 2001. Chemosensory assessment of predation risk by
slimy sculpins (Cottus cognatus): responses to alarm, disturbance and predator cues. J. Chem.
Ecol. 27: 533–546.
Chivers, D.P., and R.S. Mirza. 2001. Predator diet cues and the assessment of predation risk by aquatic vertebrates: a review and prospectus. Chemical Signals in Vertebrates, Volume 9, pp. 277–284. A. Marchlewska-Koj, J.J. Lepri and D. Müller-Schwarze (eds). Plenum Press: New York.
Chivers, D.P., and R.J.F. Smith. 1994a. The role of experience and chemical alarm signalling in predator recognition by fathead minnows, Pimephales promelas. J. Fish Biol. 44: 273–285. Chivers, D.P., and R.J.F. Smith. 1994b. Fathead minnows, Pimephales promelas, acquire predator
recognition when alarm substance is associated with the sight of unfamiliar fish. Anim.
Behav. 48: 597–605.
Chivers, D.P., and R.J.F. Smith. 1995. Free-living fathead minnows rapidly learn to recognize pike as predators. J. Fish Biol. 46: 949–954.
Chivers, D.P., and R.J.F. Smith. 1998. Chemical alarm signalling in aquatic predator-prey systems: a review and prospectus. Ecoscience, 5: 338–352.
Chivers, D.P., G.E. Brown, and R.J.F. Smith. 1996. Evolution of chemical alarm signals: Attracting predators benefits alarm signal senders. Am. Nat. 148: 649–659.
Chivers, D.P., M.H. Puttlitz, and A.R. Blaustein. 2000. Chemical alarm signaling by reticulate sculpins (Cottus perplexus). Environ. Biol. Fishes. 57: 347–352.
Chivers, D.P., R.S. Mirza and J. Johnston. 2002. Learned recognition of heterospecific alarm cues enhances survival during encounters with predators. Behaviour 139: 929–938.
Commens, A. and A. Mathis. 1999. Alarm pheromones of rainbow darters (Etheostoma caeruleum): responses to skin extracts of conspecifics and congeners (Etheostoma juliae). J. Fish Biol. 55: 1359–1362.
Dugatkin, L.A. and J.-G.J. Godin. 1992. Prey approaching their predators: A cost-benefit perspective. Ann. Zool. Fenn. 29: 233–252.
Garcia, C., E. Rolan-Alvarez and L. Sanchez. 1992. Alarm reaction and alert state in Gambusia
affinis (Pisces, Poeciliidae) in response to chemical stimuli from injured conspecifics. J. Ethol.
10: 41–46.
Göz, H. 1941. Über den Art-und Individualgeruch bei Fischen. Z. Vergl. Physiol. 29: 1–45. Hartman, E.J. and M.V. Abrahams. 2000. Sensory compensation and the detection of predators:
the interaction between chemical and visual information. Proc. R. Soc. Lond. B. 267: 571– 575.
Hamdani, E.H. and K.B. Döving. Functional organization of the olfactory system in fish. In:
Communication in Fishes. F. Ladich, S.P. Collin, P. Moller and B.G. Kapoor (eds). Science
Publisher Inc., Enfield, NH. In press.
Hazlett, B.A. 1985. Disturbance pheromones in the crayfish, Orconectes virilis. J. Chem. Ecol. 11: 1695–1711.
Hazlett, B.A. 1990. Source and nature of disturbance—chemical system in crayfish. J. Chem. Ecol. 16: 2263–2275.
Jordao, L.C., and G.L. Volpato. 2000. Chemical transfer of warning information in non-injured fish. Behaviour 137: 681–690.
Keenleyside, M.H.A. 1955. Some Aspects of the Schooling Behaviour of Fish. Leiden, Holland: E.J. Brill.
Kats, L.B., and L.M. Dill. 1998. The scent of death: Chemosensory assessment of predation risk by prey animals. Ecoscience 5: 361–394.
Kiesecker, J.M., D.P. Chivers, A. Marco, C. Quilchano, M.T. Anderson and A.R. Blaustein. 1999. Identification of a disturbance signal in larval red-legged frogs (Rana aurora). Anim. Behav. 57: 1295–1300.
Korpi, N.L. and B.D. Wisenden. 2001. Learned recognition of novel predator odour by zebra danios, Danio rerio, following time-shifted presentation of alarm cue and predator odour.
Environ. Biol. Fishes. 61: 205–211.
Kristensen, E.A., and G.P. Closs. 2004. Anti-predator response of naïve and experienced common bully to chemical alarm cues. J. Fish Biol. 64: 643–652.
Kusch, R.C., R.S. Mirza, R.S., and D.P. Chivers. 2004. Making sense of predator scents: Investigating the sophistication of predator assessment abilities of fathead minnows. Behav.
Ecol. Sociobiol. 55: 551–555.
Lawrence, B.J., and R.J.F. Smith. 1989. Behavioral response of solitary fathead minnows, Pimephales
promelas, to alarm substance. J. Chem. Ecol. 15: 209–219.
Leduc, A.O.H.C., J.M. Kelly and G.E. Brown. 2004. The effects of weak acidic conditions on chemical alarm signalling in salmonids: Laboratory and field evidence. Oecologia 139: 318– 324.
Lima, S.L., and L.M. Dill. 1990. Behavioral decisions made under the risk of predation: A review and prospectus. Can J. Zool. 68: 619–640.
Magurran, A.E. 1989. Acquired recognition of predator odour in the European minnow (Phoxinus
phoxinus). Ethology 82: 216–223.
Magurran, A.E., P.W. Irving, and P.A. Henderson. 1996. Is there a fish alarm pheromone? A wild study and critique. Proc. R. Soc. Lond. 263: 1551–1556.
Mathis, A., and R.J.F. Smith. 1993a. Chemical alarm signals increase the survival time of fathead minnows (Pimephales promelas) during encounters with northern pike (Esox lucius). Behav.
Ecol. 4: 260–265.
Mathis, A., and R.J.F. Smith. 1993b. Fathead minnows, Pimephales promelas, learn to recognize northern pike, Esox lucius, as predators on the basis of chemical stimuli from minnows in the pike’s diet. Anim. Behav. 46: 645–656
Mathis, A., D.P. Chivers, and R.J.F. Smith. 1993. Population differences in responses of fathead minnows (Pimephales promelas) to visual and chemical stimuli from predators. Ethology, 93: 31–40.
Mirza, R.S., and D.P. Chivers. 2001a. Do juvenile yellow perch use diet cues to assess the level of threat posed by intraspecific predators? Behaviour 138: 1249–1258.
Mirza, R.S. and D.P. Chivers. 2001b. Chemical alarm signals enhance survival of brook charr (Salvelinus fontinalis) during encounters with chain pickerel (Esox niger). Ethology 107: 989– 1006.
Mirza, R.S. and D.P. Chivers. 2001c. Are chemical alarm signals conserved within salmonid fishes? J. Chem. Ecol. 27: 1641–1655.
Mirza, R.S., and D.P. Chivers. 2001d. Learned recognition of heterospecific alarm signals: The importance of a mixed predator diet. Ethology 107: 1007–1018.
Mirza, R.S., and D.P. Chivers. 2002. Behavioural responses to conspecific disturbance chemicals enhance survival of juvenile brook charr, Salvelinus fontinalis, during encounters with predators. Behaviour 139: 1099–1110.
Mirza, R.S., and D.P. Chivers. 2003a. Response of juvenile rainbow trout to varying concentrations of chemical alarm cues: Response thresholds and survival during encounters with predators
Can. J. Zool. 81: 88–95.
Mirza, R.S., and D.P. Chivers. 2003b. Fathead minnows learn to recognize heterospecific alarm cues they detect in the diet of a known predator. Behaviour 140: 1359–1370.
Mirza, R.S., S.A. Fisher, and D.P. Chivers. 2003. Assessment of predation risk by juvenile yellow perch (Perca flavescens): Responses to alarm cues from conspecifics and prey guild members.
Env. Biol. Fishes 66: 321–327
Mirza, R.S., J.J. Scott, and D.P. Chivers. 2001. Differential responses of male and female red swordtails to chemical alarm cues. J. Fish Biol. 59: 716–728.
Moore, P.A., M.J. Weissburg, J.M. Parrish, R.K. Zimmer-Faust, and G.A. Gerhardt. 1994. The spatial distribution of odours in simulated benthic boundary layer flows. J. Chem. Ecol. 20: 255–279.
Pfeiffer, W. 1977. The distribution of fright reaction and alarm substance cells in fishes. Copeia 1977: 653–665.
Pollock, M.S., and D.P. Chivers. 2003a. Habitat characteristics influence the ability of fathead minnows to learn to recognize heterospecific alarm cues. Can. J. Zool. 81: 923–927. Pollock, M.S., and D.P. Chivers. 2003b. The effects of density on learned recognition of
heterospecific alarm cues. Ethology 110: 341–349.
Suboski, M.D. 1990. Releaser-induced recognition learning. Psychol. Rev. 97: 271–284.
Smith, M.E., and M.C. Belk. 2001. Risk assessment in western mosquitofish (Gambusia affinis): Do multiple cues have additive effects? Behav. Ecol. Sociobiol. 51: 101–107.
Smith, R.J.F. 1992. Alarm signals in fishes. Rev. Fish Biol. Fish. 2: 33–63.
Smith, R.F. J. 1979. Alarm reaction of Iowa and Johnny darters (Etheostoma, Percidae, Pisces) to chemicals from injured conspecifics. Can. J. Zool. 57: 1278–1282.
Stabell, O.B., and M.S. Lwin. 1997. Predator-induced phenotypic changes in crucian carp are caused by chemical alarm signals from conspecifics. Environ. Biol. Fishes 49: 145–149. Tremaine, R.J., M.S. Pollock, R.G. Friesen, R.C. Kusch, and D.P. Chivers. In press. The response
of prey fishes to chemical alarm cues: What recent field experiments reveal about the old testing paradigm. Chemical Signals in Vertebrates, Volume 10. R.T. Mason, M. LeMaster and D. Müller-Schwarze (eds). Plenum Press: New York.
Vilhunen, S., and H. Hirvonen. 2003. Innate antipredator responses of Arctic charr (Salvelinus
alpinus) depend on predator species and their diet. Behav. Ecol. Sociobiol. 55: 1–10.
Von Frisch, K. 1938. Zur Psychologie des Fisch-Schwarmes. Naturwissenschaften 26: 601–606. Von Frisch, K. 1941. Über einen Schreckstoff der Fischhaut und seine biologische Bedeutung. Z.
Vergl. Physiol. 29: 46–149.
Wisenden, B.D. 2003. Chemically-mediated strategies to counter predation. In: Sensory Processing
in the Aquatic Environment. S.P. Collin and N.J. Marshall (eds). Springer-Verlag, New York,
Wisenden, B.D. and K. Barbour. 2005. Antipredator responses to skin extract of redbelly dace Phoxinecseos, by free-ranging populations of redbelly dace and fathead minnows. Pimephaleo
promelas. Environ. Biol. Fishes 72: 227–233.
Wisenden, B.D. and K.R. Harter. 2001. Motion, not shape, facilitates association of predation risk with novel objects by fathead minnows (Pimephales promelas). Ethology 107: 357–364. Wisenden, B.D., and N.E. Stacey. 2005. Fish semiochemicals and the network concept. In: Animal
Communication Networks. P.K. McGregor (ed.). Cambridge University Press. In press.
Wisenden, B.D., D.P. Chivers, and R.J.F. Smith. 1995. Early warning of risk in the predation sequence: a disturbance pheromone in Iowa darters (Etheostoma exile). J. Chem. Ecol. 21: 1469–1480.
Wisenden, B.D., M.S. Pollock, R.J. Tremaine, J.M. Webb, M.E. Wismer, and D.P. Chivers. 2003. Synergistic interactions between chemical alarm cues and the presence of conspecific and heterospecific fish shoals. Behav. Ecol. Sociobiol. 54: 485–490.