On the use of micro-electrodes in fish pond
sediments
Leon E. Meijer, Yoram Avnimelech *
Faculty of Agricultural Engineering,Technion-Israel Institute of Technology,32000Haifa,Israel
Received 2 March 1999; accepted 17 July 1999
Abstract
The surface layer of fish pond sediments is characterized by very high microbial activity. Oxygen uptake by the microbes at the sediment surface leaves little oxygen to penetrate the sediment. In marine and freshwater lake sediments, oxygen only penetrated a few millime-ters. In this study, oxygen penetration into the sediment layer in fish ponds and simulated fish ponds was measured with an oxygen micro-electrode. Oxygen profiles determined with micro-electrodes showed that oxygen does not penetrate deeper than 1 mm under quiescent conditions in intensive and semi-intensive fish pond sediments. Sediment oxygen consump-tion (SOD) at approximately 25°C, calculated from measured oxygen profiles, ranged from 45 to 50 mg O2/m2 per h. Change in water temperature and the addition of formaldehyde
showed that most of the oxygen consumption in the sediment surface layer is related to biological activity. Redox potential profiles of fish pond sediments measured with a micro-platinum electrode, showed reducing conditions conducive to sulfide formation at 2 – 4 mm below the sediment surface. Nitrate addition to the overlying water poised redox potential above 300 mV to a sediment depth of about 15 mm. Nitrate thus buffered the sediment – wa-ter inwa-terface against reducing conditions in the sediment and the possible diffusion of toxic reduced end products to the water. © 1999 Elsevier Science B.V. All rights reserved.
Keywords:Fish pond sediment; Micro-electrodes; Oxygen; Redox; Nitrate
* Corresponding author. Tel.: +972-4-829-2480; fax:+972-4-822-1529.
E-mail address:[email protected] (Y. Avnimelech)
1. Introduction
The microbial population density in the surface layers of fish pond sediments is several orders of magnitude higher than the microbial population in the water (Ram et al., 1982). Although uneaten food, faeces, and dead algae sink to the bottom, the sediment is more than a sink. It plays an important role in the recycling of nutrients back into the water, thereby affecting the water quality. In aquaculture models, sediment is often described as a nutrient sink (Lorenzen et al., 1997) and its relevance to the pond as a whole often ignored (van Dam, 1990). Information on sediment – water interactions in fish ponds is limited partially due to the lack of suitable analytical tools to en-able measurement of concentration profiles in the upper sediment layer (Piedrahita, 1991). Sectioning of sediment cores of the soft and flocculent surface layer in segments smaller than 0.5 cm is extremely difficult. Similar methodol-ogical constraints were encountered in research of marine, lake and river sedi-ments (Revsbech et al., 1980a; De Beer and Sweerts, 1989; Christensen et al., 1989).
Revsbech et al. (1980a) introduced the use of oxygen-micro-electrodes with tip diameters between 10 and 100 mm in the study of marine sediments. Micro-elec-trodes enable fine-scale oxygen measurement of surface layers with little distur-bance of the physical environment during measurement. Using oxygen micro-electrodes, it was found that oxygen penetrated only a few millimeters into littoral lake and coastal sea sediments (Revsbech et al., 1980b; Sweerts, 1990). Oxygen micro-electrodes have not yet been applied in fish pond sediments stud-ies.
If available, oxygen is the dominant terminal electron acceptor during the decomposition of organic matter. With the depletion of oxygen, the micro-bial community reduces other electron acceptors in a characteristic thermo-dynamic sequence of NO3−, Mn4+, Fe3+, SO
4
2−, and HCO
3
−. Each reduction
takes place within a specific redox potential range (Reddy et al., 1986). A redox potential profile, which is a cross-section of the sediment redox poten-tial at different depths, thus provides an indication of the redox status of the sediment. In the presence of electron acceptors that yield higher energy, such as oxygen and nitrate, the sequential reduction of the electron accep-tors is inhibited. Nitrate added to anoxic lake sediment inhibited the reduction of iron (Ripl, 1976). Jenneman et al. (1986) added nitrate to sewage sludge and inhibited the formation of sulfide and methane. The redox-buffering effect of nitrate might protect the sediment – water interface against reducing con-ditions in the sediment and the possible diffusion of toxic end products to the water.
2. Materials and methods
2.1.Fish ponds
Undisturbed sediment cores were collected from an intensive fish pond at kibbutz Ein Hamifratz and a similar pond at kibbutz Afikim, Israel. The ponds had an area of ca 3 ha each, a water depth of 1 – 1.4 m and a tilapia biomass of about 6 t/ha. Cores, 59-mm diameter plexiglass tubes, two from each pond, were transported to the laboratory, and the overlying water was aerated. Height of the water above the sediment in the cores was maintained between 20 and 25 cm and depth of the sediment was approximately 15 cm. Temperature of the overlying water was 21.5 and 26°C in cores from Afikim and Ein Hamifratz, respectively. Measurements of oxygen and redox potentials were conducted within 24 h of core retrieval. Oxygen profiles were measured with a Clark-style oxygen micro-electrode (Diamond Gen-eral, Ann Arbor, MI) with a tip diameter of 200mm (Revsbech and Ward, 1983). Smaller tip diameters are available, but the risk of breakage while inserting into the sediment increases with decreasing tip diameter. A coaxial shielded cable connected the oxygen micro-electrode to an electrometer (Keithly 614 including a picoamme-ter, polarization voltage – 0.75 V). Micro-electrode position was controlled with a micromanipulator (0.01 mm resolution) (Stoelting, Wood Dale, IL). Before mea-surement, the micro-electrode was inserted in the overlying water for at least 20 min for thermal equilibration. Current output in the upper 5 cm of the overlying water was taken as a measure of 100% saturation. Measurements were made at 0.5-mm vertical intervals from 5 to 1 mm above the sediment. Thereafter, measurements were made at 0.25-mm intervals down to a depth in the sediment where current output between additional steps did not change, indicating a zero oxygen concentration.
Oxidation-reduction (redox) potential profiles were measured in the same cores. Platinum electrodes (Delta Institute for Hydrobiological Research, Yerseke, The Netherlands) with a tip diameter of 0.5 mm were connected by an insulated wire to a pH meter (Radiometer, Copenhagen, Denmark). A calomel reference electrode (Orion, Boston, MA), placed in the overlying water of the core, completed the circuit. In order to standardize the redox potential, based on the standard hydrogen reference electrode, 244 mV was added to the measured potential (Faulkner et al., 1989). The platinum electrode was moved along the sediment core using the micromanipulator. Measurements were made in vertical intervals of 1 mm from 2 mm above the sediment surface to a sediment depth of 30 mm. Readings were made when drift was B1 mV/min, commonly after 20 – 30 min.
2.2.Role of biological oxygen consumption
the final profile was measured 11 days after formaldehyde addition. The influence of temperature on the oxygen profile was assessed by recording the oxygen profile in a core from an Ein Hamifratz fish pond in which temperature of the overlying water was 26°C. The same core was cooled in a refrigerator under continuous aeration until the temperature of the overlying water was 12°C, and the oxygen profile measured again. Equilibration of the electrode and measurement of the oxygen profile (approximately 30 min) was conducted at room temperature. Water temperature in the core did not rise during this time.
2.3.Simulated fish ponds
Cores collected in simulated fish ponds (Meijer et al., unpublished) were used to study the effects of feeding and nitrate additions on redox and oxygen profiles. Simulated ponds consisted of 100-cm diameter tanks, filled with a 25-cm layer of soil from a fish pond and with water to a depth of 65 cm above the soil layer. Aeration to the water was provided with fine air bubbles at mid-height of the water to prevent mixing and stirring of the sediment layer. Feed (28% protein) was added to duplicated tanks once daily (without fish) at a rate representing actual fish pond conditions (10 and 20 g/day for the low and high feed treatments, respectively). Oxygen and redox profiles of the aerated undisturbed sediment cores were mea-sured as described above.
2.4.Nitrate addition
The influence of nitrate on the redox profile of the Afikim fish pond sediment was examined by the addition of nitrate in the form of NaNO3 to the overlying water
of the core. Nitrate was maintained at 15 mg NO3– N/l and the redox potential
measured after 30 days of incubation. During the incubation, the overlying water in the core was aerated with an airstone.
Nitrate (as NaNO3) was added to one of duplicated tanks with a feed addition
rate of 22 g/m2per day. A core was collected from this pond; nitrate concentration
in the water at the time of coring was 34 mg NO3– N/l. Redox profiles in the aerated undisturbed sediment cores were measured as described previously.
3. Results
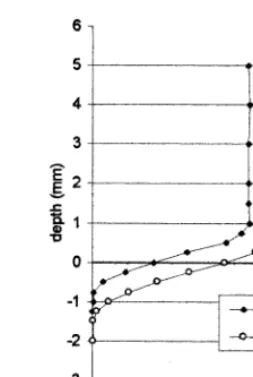
Oxygen concentrations in the water dropped linearly from saturation to less than 50% saturation across a 1 mm water layer immediately above the sediment surface. Oxygen was consumed completely at a depth of 1.00 and 1.25 mm in the Ein Hamifratz pond and the Afikim pond sediments, respectively (Fig. 1). Oxygen profiles in cores from the same pond were almost identical, therefore, only one profile for each pond is presented.
thin film of water to ‘stick’ to the sediment surface. Within this film, called the diffusive boundary layer (DBL), transport of solute occurs by molecular diffusion (Santschi et al., 1983). Thickness of the DBL can be measured with oxygen micro-electrodes by determining the depth of the water layer with a linear oxygen decrease just above the sediment (Jørgensen and Revsbech, 1985). Thickness of the DBL was 1.00 and 0.75 mm in the Ein Hamifratz pond and the Afikim pond sediments, respectively (Fig. 1). When oxygen penetrates only to a depth of 2 mm or less, almost the entire concentration gradient can be found in the DBL. Oxygen uptake can then be calculated from the slope of the linear concentration profile through the DBL, using Fick’s first law of diffusion:
J=DsfdC/dx (1)
where J is the oxygen flux, equal to the consumption of oxygen by the sediment (mg/m2per h), D
s represents the diffusion coefficient of oxygen in the sediment,f
is the porosity, and dC/dx is the oxygen concentration gradient. Since the DBL consists of water,Dsis equal toD0, the temperature-dependent diffusion coefficient in water (Broecker and Peng, 1974), the porosity is 1, and the concentration gradient dC/dx is the gradient in the DBL.
Using the measured oxygen concentration gradients and Eq. (1), calculated oxygen fluxes toward the sediment were 45 and 50 mg/m2per h for the Afikim and
Ein Hamifratz ponds, respectively.
Oxygen penetration in the simulated fish ponds sediments (Fig. 2) was 0.25 and 1.5 mm for the high and low food addition rate, respectively. Oxygen profiles from cores taken at the same feeding rate did not differ, therefore only one profile is presented (Fig. 2) The DBL was 1.5 mm thick in both treatments. Oxygen consumption of these sediments was 40 mg/m2 per h at the high feed addition rate
[image:5.612.188.308.384.555.2]and 16 mg/m2 per h at the low feed addition rate.
Fig. 2. Oxygen profiles in sediment cores from simulated fish ponds under two food addition rates (food containing 28% protein).
Addition of formaldehyde sterilizes the water and, although limited by the downward diffusion of the sterilizing agent, suppresses sediment microbial activity. Following addition of formaldehyde, increased penetration of oxygen into the sediment core was measured (Fig. 3). Oxygen penetrated to 13 mm 11 days following addition of the sterilizing agent. Before addition of formaldehyde the DBL was 1 mm thick and the SOD based on the oxygen decrease in the DBL was 35 mg/m2 per h. With the increased oxygen penetration after formaldehyde
[image:6.612.184.313.354.544.2]Fig. 4. Oxygen profiles in sediment core of a fish pond from Afikim at two different temperatures of the overlying water.
addition, most of the oxygen gradient moved out of the DBL into the sediment. Calculation of SOD based on the oxygen decrease in the DBL is thus not longer sound. In the sediment both porosity and the diffusion coefficient play a significant role in the oxygen flux (Eq. (1)). Porosity of the surface layer of simulated fish pond sediment was 0.90 (Meijer et al., submitted). Sweerts et al. (1991) found that in high porosity sedimentsD0/Dswas 1.5. This information allows calculation of the SOD
based on the oxygen gradient in the top 4 mm sediment layer. When formaldehyde was added, SOD decreased to 13 and 4 mg/m2 per h after 0.5 h and 11 days,
respectively.
A further indication of the biological nature of oxygen consumption by sediment was obtained when oxygen profiles in sediment cores were measured at 12 and 26°C (Fig. 4). With a temperature decrease from 26 to 12°C, the saturated concentration of oxygen in the overlying water increased from 8.09 to 10.76 mg O2/l and the oxygen penetration in the sediment increased from 1.25 to 2.00 mm. Based on the linear oxygen concentration gradient in the DBL, the oxygen consumption of the sediment at 26 and 12°C was 43 and 22 mg/m2 per h, respectively.
The redox potential in the overlying water and at the topmost sediment layer is around 400 mV, as expected for oxygenated systems (Revsbech et al., 1980a,b). However, it dropped sharply down core and, at a depth of 12 mm, the redox potential was below −100 mV.
Fig. 5. Redox potential profiles in a sediment core of a fish pond from Afikim without nitrate in the overlying water of the core and after an incubation of 30 days with 15 ppm N – NO3in the overlying water.
A high feed addition rate resulted in a similar redox profile (Fig. 6) as the profile from the Afikim pond (Fig. 5) with reducing conditions 1 mm below the sediment surface. A lower organic load resulted in less reducing conditions (Fig. 6). The redox potential declined sharply with depth, reaching a redox potential of 80 mV, 3 mm below the sediment surface. The addition of nitrate to a simulated pond with a low feeding regime affected redox potential in the top 20 mm of the sediment
[image:8.612.186.313.350.523.2](Fig. 6). At deeper sediment depths, redox potentials were identical to those of the core without nitrate addition. Oxygen concentrations in the overlying water of cores were not affected by nitrate.
4. Discussion
Oxygen consumption and redox reactions are essential features of fish pond sediments. Fish production is affected by the presence of anaerobic conditions in the sediment (Avnimelech and Zohar, 1986). It was estimated that oxygen pene-trates several cm into the sediment (Boyd, 1990; Hussenot and Martin, 1995). The penetration of oxygen into the sediment is limited due to the high sediment oxygen demand (SOD) and to the slow oxygen diffusion in the sediment. This limitation is expected to generate very steep gradients of oxygen and redox potential in sediment surface layers. Methods enabling measurements of oxygen concentration and redox potential at very fine intervals are needed to study sediment oxygen and redox stratification.
Micro-electrode technology has been developed, adapted and utilized to study microbial mats (Revsbech et al., 1983), marine sediments (Revsbech et al., 1980a,b), lake sediments (Sweerts, 1990), and river sediments (Christensen et al., 1989). This technique enabled to measure fine concentration gradients. Oxygen was found to penetrate only a few millimeters into littoral lake and coastal sea sediments. Oxygen profiles in fish pond sediments were expected to be steeper due to high organic matter loading of such ponds (Blackburn, 1987).
Oxygen penetration into fish pond sediments was found in this work to be about 1 mm, similar to oxygen penetration of 1 – 2 mm for sediments of shallow water bodies (Revsbech et al., 1980b; Sweerts, 1990; Lindeboom and Sandee, 1984). The domain in which strict aerobic benthic activity can take place is accordingly, very limited.
SOD is a reflection of benthic respiration, although chemical oxidations of reduced species diffusing up from deeper sediment layers also takes place. The deeper penetration of oxygen following formaldehyde addition and the effect of temperature on oxygen penetration are both indicators of the biological nature of this process. Reduced water temperature has two effects: (1) increased saturation concentration in overlying water; and (2) reduced metabolic activity of the sedi-ment. The concomitant deeper oxygen penetration, however, increases the sediment layer in which aerobic bacteria can be active. The resulting biological sediment oxygen consumption might therefore be larger than expected. Hargreaves and Tucker (1996) found that nitrification in channel catfish ponds was important in spring and fall. Greater dissolved oxygen concentration and reduced microbial activity associated with lower water temperatures in those seasons led to an increased oxygen penetration in the sediment relative to summer conditions, thereby increasing the volume of aerobic sediment capable of nitrification.
mm thick, between oxidized and reduced regions. Highly reducing conditions (B−100 mV) were measured a few millimeters below the sediment surface. Those reducing conditions result in sulfide formation as sulfate is reduced below a redox potential of −100 mV (Engler and Patrick, 1973). Lindeboom and Sandee (1984) found similar redox profiles in a salt water lake sediment rich in organic carbon. The suggestion by Hussenot and Martin (1995) to assess the reducing condition of fish pond sediments by measuring redox potential every 2 cm overlooks the fine stratification of redox profiles.
Nitrate in the overlying water served as a potent redox buffer. It diffused into the sediment, served as an electron acceptor once oxygen was consumed and poised the redox potential. Addition of nitrate to water led to the relocation of reducing conditions away from the sediment surface to deeper layers, about 15 mm below the sediment surface. The exposure of benthic crustaceans or fish to anaerobic metabolites is thus greatly reduced.
The change in color of the sediment, from black to brown, indicates the transition from a reduced system to the dominance of high redox potentials. This may be induced by oxygen (Boyd, 1990), but can also be induced by other oxidants such as nitrate and oxidized manganese and iron (Revsbech and Jørgensen, 1986). The increase of redox potential in the sediment was visually indicated by the increase in depth of the brown surface layer.
A clear oxygen concentration gradient across an immobile benthic boundary layer, 1 mm thick, was found in the non-stirred sediment core systems. The diffusion through this layer is equal, under steady-state conditions, to the sediment oxygen flux or consumption. The oxygen diffusion rate into the sediment, calcu-lated from Fick’s first law, was 40 – 50 mg O2/m2per h at approximately 25°C in the
two fish ponds and in the simulated pond with a high food load. Similar values were obtained from in situ measurements with static cores immersed onto the sediment of fish ponds (Shapiro and Zur, 1981; Boyd and Teichert-Coddington, 1994).
(Ola´h et al., 1987). Costa-Pierce et al. (1984), simulating wind resuspension with a stirrer, found SOD values in prawn ponds as high as 249 mg O2/m2 per h.
Madenjian et al. (1987) found a SOD of 240 mg O2/m2per h for prawn ponds using
whole pond methodology and SOD in catfish ponds, measured by Berthelson et al. (1996) was found to be 185 mg O2/m2 per h.
The DBL constitutes a barrier to mass transfer across the sediment – water interface. When the oxygen consumption by the sediment is high, as in fish ponds, the oxygen uptake of the sediment is largely governed by DBL hindrance (Hall et al., 1989; Jørgensen and Des Marais, 1990). Oxygen uptake, and diffusion of other substances is therefore highly sensitive to variations in DBL thickness and thus to flow velocity. As flow velocity increases, the DBL thickness decreases and the SOD increases (Sweerts et al., 1989).
5. Conclusions
Oxygen does not penetrate much deeper than 1 mm under quiescent conditions in intensive and semi-intensive fish pond sediments. Deeper penetration depends on bioturbation and mixing of sediment surface layers, commonly occurring in fish ponds.
Nitrate addition influenced the redox profile in the sediment on a micro-scale. Under quiescent conditions, a very thin, 1 – 4 mm oxidized layer separated the water from highly reduced sediment. By adding nitrate to the water, the thickness of the oxidized sediment layer was increased to about 15 mm, potentially reducing the exposure of fish to toxic anaerobic metabolites.
Most processes at the sediment, such as organic matter decomposition, are mediated by micro-organisms and because of the size of those organisms, take place on a micro-scale. Therefore, micro-electrodes such as the oxygen micro-electrode, have good potential for the study of fish ponds sediments. No other method has similar spatial and temporal resolutions, the result of the measurement is known within seconds and a new measurement can be taken as soon as a stable concentra-tion is established. The method is non-destructive and repetitive measurements can be made on the same place.
References
Avnimelech, Y., Zohar, G., 1986. The effect of local anaerobic conditions on growth retardation in aquaculture systems. Aquaculture 58, 167 – 174.
Avnimelech, Y., Kochva, M. and Hargreaves J.A., 1999. Sedimentation and resuspension in earthen fish ponds, J. World Aquaculture Soc. (000 – 000).
De Beer, D., Sweerts, J-P.R.A., 1989. Measurements of nitrate gradients with an ion-selective micro-elec-trode. Anal. Chim. Acta 219, 351 – 356.
Berthelson, C.R., Cathcart, T.P., Pote, J.W., 1996. In situ measurements of sediment oxygen demand in catfish ponds. Aquacult. Eng. 15, 261 – 271.
Boyd, C.E., Teichert-Coddington, D., 1994. Pond bottom soil respiration during fallow and culture periods in heavily-fertilized tropical fish ponds. J. World Aquaculture Soc. 25, 417 – 423.
Broecker, W.S., Peng, T.-H., 1974. Gas exchange rates between air and sea. Tellus 26, 21 – 35. Christensen, P.B., Nielsen, L.P., Revsbech, N.P., Sørensen, J., 1989. Microzonation of denitrification
activity in stream sediments as studied with a combined oxygen and nitrous oxide microsensor. Appl. Environ. Microbiol. 55 (5), 1234 – 1241.
Costa-Pierce, B.A., Craven, D.B., Karl, D.M., Laws, E.A., 1984. Correlation of in situ respiration rates and microbial biomass in prawn (Macrobrachium rosenbergii) ponds. Aquaculture 37, 157 – 168. van Dam, A.A., 1990. Modeling of aquaculture pond dynamics. In: Berka, R., Hilge, V. (Eds.),
Production enhancement in still-water pond culture. Proceedings of the FAO-EIFAC Symposium on Production Enhancement in Still-Water Pond Culture, 15 – 18 May 1990, Prague, Czechoslovakia. Engler, R.M., Patrick, W.H. Jr, 1973. Sulfate reduction and sulfide oxidation in flooded soil as affected
by chemical oxidants. Soil Sci. Soc. Am. Proc. 37, 685 – 688.
Faulkner, S.P., Patrick, W.H. Jr, Gambrell, R.P., 1989. Field techniques for measuring wetland soil parameters. Soil Sci. Soc. Am. J. 53, 883 – 890.
Hall, P.O.J., Anderson, L.G., Rutgers van der Loeff, M.M., Sundby, B., Westerlund, S.F.G., 1989. Oxygen uptake kinetics in the benthic boundary layer. Limnol. Oceanogr. 34 (4), 734 – 746. Hargreaves, J.A., Tucker, C.S., 1996. Evidence for control of water quality in channel catfishIctalurus
punctatusponds by phytoplankton biomass and sediment oxygenation. J. World Aquacult. Soc. 27, 21 – 29.
Hussenot, J., Martin, J.-L.M., 1995. Assesment of the quality of pond sediment in aquaculture using simple, rapid techniques. Aquacult. Int. 3, 123 – 133.
Jenneman, G.E., McInerney, M.J., Knapp, R.M., 1986. Effect of nitrate on biogenic sulfide production. Appl. Environ. Microbiol. 51 (6), 1205 – 1211.
Jørgensen, B.B., Des Marais, D.J., 1990. The diffusive boundary layer of sediments: oxygen microgradi-ents over a microbial mat. Limnol. Oceanogr. 35 (6), 1343 – 1355.
Jørgensen, B.B., Revsbech, N.P., 1985. Diffusive boundary layers and the oxygen uptake of sediments and detritus. Limnol. Oceanogr. 30 (1), 111 – 122.
Lindeboom, H.J., Sandee, A.J.J., 1984. The effect of coastal engineering projects on microgradients and mineralization reactions in sediments. Water Sci. Technol. 16, 87 – 94.
Lorenzen, K., Struve, J., Cowan, V.J., 1997. Impact of farming intensity and water management on nitrogen dynamics in intensive pond culture: a mathematical model applied to Thai commercial shrimp farms. Aquacult. Res. 28, 493 – 507.
Madenjian, C.P., Rogers, G.L., Fast, A.W., 1987. Predicting night time dissolved oxygen loss in prawn ponds of Hawaii: Part I. Evaluation of traditional methods. Aquacult. Eng. 6, 191 – 208.
Ola´h, J., Sinha, V.R.P., Ayyappan, S., Purushothaman, C.S., Radheyshyam, S., 1987. Sediment oxygen consumption in tropical undrainable fish ponds. Int. Rev. Ges. Hydrobiol. 72, 297 – 305.
Piedrahita, R.H., 1991. Modelling water quality in aquaculture ecosystems. In: D.E. Brune and J.R. Tomasso (Eds.), Aquaculture and water quality. Advances in World Aquaculture 3, World Aquacul-ture Society, pp. 322 – 362.
Ripl, W., 1976. Biochemical oxidation of polluted lake sediments with nitrate — a new lake restoration method. Ambio 5 (3), 132 – 135.
Reddy, R.C., Feijtel, T.C., Patrick, W.H. Jr, 1986. Effects of soil redox conditions on microbial oxidation of organic matter. In: Chen, Y., Avnimelech, Y. (Eds.), The Role of Organic Matter in Modern Agriculture. Martinus Nijhoff, Dordrecht, pp. 117 – 156.
Revsbech, N.P., Jørgensen, B.B., 1986. Micro-electrodes: their use in microbial ecology. Adv. Microb. Ecol. 9, 293 – 352.
Revsbech, N.P., Ward, D.M., 1983. Oxygen micro-electrode that is insensitive to medium chemical composition: Use in an acid microbial mat dominated by Cyanidium caldarium. Appl. Environ. Microbiol. 45, 755 – 759.
Revsbech, N.P., Jørgensen, B.B., Blackburn, T.H., 1980a. Oxygen in the seabottom measured with a micro-electrode. Science 207, 1355 – 1356.
Revsbech, N.P., Jørgensen, B.B., Blackburn, T.H., Cohen, Y., 1983. Micro-electrode studies of the photosynthesis and O2, H2S and pH profiles of a microbial mat. Limnol. Oceanogr. 28 (6), 1062 – 1074.
Santschi, P.H., Bower, P., Nyffeler, U.P., Azevedo, A., Broecker, W.S., 1983. Estimates of the resistance to chemical transport posed by the deep-sea boundary layer. Limnol. Oceanogr. 28, 899 – 912. Shapiro, J., Zur, O., 1981. A simple in situ method for measuring benthic respiration. Water Res. 15,
283 – 285.
Sweerts, J-P.R.A., 1990. Oxygen consumption, mineralization and nitrogen cycling at the sediment – wa-ter inwa-terface of North Temperate lakes. PhD thesis, Groningen, p. 136.
Sweerts, J-P.R.A., StLouis, V., Cappenberg, T.E., 1989. Oxygen concentration profiles and exchange in sediment cores with circulated overlying water. Freshwater Biol. 21, 401 – 409.
Sweerts, J-P.R.A., Kelly, C.A., Rudd, J.W.M., Hesslein, R., Cappenberg, T.E., 1991. Similarity of whole-sediment molecular diffusion coefficients in freshwater sediments of low and high porosity. Limnol. Oceanogr. 36 (2), 335 – 342.