Basic Terminology
and Definitions
The definition of overactive bladder (OAB) has changed over the years, from regarding it only as a symptom to understanding it as a complicated pathophysiological syndrome. The most common definition of OAB was established in 2002 by the International Continence Society (ICS) as urinary urgency with or without urge incontinence, usu-ally associated with frequency and nocturia, in the absence of proven pathological or metabolic disor-ders (such as lower urinary tract (LUT) infection, bladder cancer, benign prostatic enlargement) or other obvious pathology [1].
However, a review of the ICS 2002 terminol-ogy report, published in 2006, recommends re-placing certain terms in the definition given above. The revision suggests that “urge incontinence” should be replaced with “urgency incontinence” to emphasize the fact that the incontinence is due to urgency and not an urge (which can be under-stood as a „desire”); and that the word “frequency” should be substituted by the term “increased day-time frequency”. Thus, the revised OAB definition is urgency with or without urgency incontinence (UI), usually with increased daytime frequency and nocturia. “Urgency” is defined as “the complaint of a sudden compelling desire to pass urine, which is difficult to defer” [2, 3]. Thus, “urgency urinary
Łukasz Dobrek
1, Kajetan Juszczak
1,2, Marek Wyczółkowski
2, Piotr Jan Thor
1Overactive Bladder – Current Definition
and Basic Pathophysiology Concepts
Współczesna definicja i patofizjologia pęcherza nadaktywnego
1Department of Pathophysiology, Jagiellonian University Medical College, Kraków, Poland 2Department of Urology, Memorial Rydygier Hospital, Kraków, Poland
Abstract
Overactive bladder (OAB) is a clinical entity with a general prevalence of about 17% in adults, characterized by urgency with or without incontinence, usually with increased daytime frequency and nocturia. The pathophysiol-ogy of OAB is complex, involving disturbances of both the central nervous system (decreased suprapontine inhi-bition of micturition) and the peripheral nervous system (damage of the axonal paths in the spinal cord, loss of peripheral inhibition or enhancement of excitatory neurotransmission in the voiding reflex) as well as myogenic disturbances (structural and ultrastructural changes in the detrusor smooth muscle). The article briefly reviews the anatomy and physiology of the lower urinary tract and the current definitions and terminology used in the literature on OAB, with a particular focus on theories explaining the pathomechanism of the syndrome (Adv Clin Exp Med 2011, 20, 2, 119–129).
Key words: lower urinary tract (LUT), overactive bladder (OAB), OAB pathophysiology.
Streszczenie
Nadaktywny pęcherz moczowy (OAB) jest zaburzeniem występującym u około 17% dorosłej populacji, charakte-ryzującym się nagłą potrzebą oddania moczu wraz z nietrzymaniem moczu lub bez niego, przebiegającym zwykle ze zwiększoną częstotliwością mikcji i nokturią. Patogeneza OAB jest złożona, obejmuje zaburzenia o charakterze neurogennym – uszkodzenie centralnych ośrodków nerwowych regulujących czynność pęcherza oraz jego aferent-nego i eferentaferent-nego unerwienia oraz zaburzenia o charakterze miogennym ściany pęcherza. W niniejszym artykule krótko omówiono anatomiczne i fizjologiczne aspekty dolnych dróg moczowych, obecną definicję i epidemiologię OAB wraz z patofizjologicznymi koncepcjami rozwoju tego zespołu (Adv Clin Exp Med 2011, 20, 2, 119–129). Słowa kluczowe: dolne drogi moczowe (LUT), nadaktywny pęcherz moczowy (OAB), patofizjologia OAB.
Adv Clin Exp Med 2011, 20, 2, 119–129 ISSN 1230-025X
EDITORIAL
incontinence” is “the complaint of involuntary leakage accompanied by or immediately preceded by urgency”; “increased daytime frequency” is “the complaint by the patient [...] that he/she voids too often by day”, while “nocturia” is defined as “the complaint that the individual has to wake at night one or more times to void” [2, 3].
Urgency is a key symptom, essential for a di-agnosis of OAB. According to the definition of OAB, this clinical entity may be recognized with-out a complaint of incontinence. Moreover, abwith-out two-thirds of OAB patients do not suffer from in-voluntary leakage of urine [4]; this is termed “dry OAB”, as opposed to “wet OAB”, in which urinary incontinence (UI) presents. Apart from OAB uri-nary incontinence, there are also other types of UI, including stress urinary incontinence (“invol-untary leakage on effort or exertion, or on sneez-ing or coughsneez-ing”) and mixed urinary incontinence (“involuntary leakage associated with urgency and with exertion, effort, sneezing or coughing”) [4].
It must be emphasized that a clinical diagnosis of OAB, based on the complaints described above, can result from abnormal detrusor function dur-ing the filldur-ing phase in a urodynamic test. Detrusor overactivity (DO) – a term used in urodynamics –refers to involuntary detrusor contractions dur-ing cystometry and is not a synonym for OAB. Detrusor overactivity – which can be neurogenic, myogenic or idiopathic – is always characterized by involuntary contractions of the detrusor dur-ing bladder filldur-ing [4]. Two kinds of DO may be distinguished: phasic and terminal. The first dem-onstrates a characteristic waveform and is typically seen in idiopathic detrusor overactivity. The latter is observed when a single detrusor contraction oc-curs and cannot be suppressed, leading to urine leakage (or even complete bladder emptying), even when the urine amount does not exceed critical volume and should be under normal control [3].
The Epidemiology
and Social Impact
of Overactive Bladder
OAB is common clinical entity, with global prevalence ranging from 3% to 43%, depending on the population studied, sex, age and differences in the accepted definition of OAB. Generally, the fre-quency of OAB increases with age and is more com-mon in women than men. Two large population-based studies conducted in Europe and the USA revealed a similar prevalence of approximately 17% of the general adult population [3]. Data from the European study indicated that the overall
preva-lence of OAB among the adults studied (18 years or older) was 16.6%, with slightly lower frequency in men (15.6%) than in women (17.4%) [5]. The US study was performed by the National Overac-tive Bladder Evaluation Program (NOBLE) and the results indicated that OAB is noted in 16% of men and 16.9% of women, and that 37% of the patients who met OAB criteria experience incontinence, while 63% do not. Dry OAB is more common in men than in women (13.6% vs. 7.6%); wet OAB is found mainly in women (9.3% vs. 2.4%) [6].
One of the latest studies estimating OAB epi-demiology is the EPIC study, based on current ICS guidelines. Conducted in Canada, Germany, Italy, Sweden and the UK, it revealed a prevalence of OAB in 11.8% of the adults participants (10.8% in men and 12.8% in women). OAB was more com-mon in men than in women after the age of 60, while under age 60 it was more common in women. Incontinence was reported in 49.2% of the women with OAB and 28.7% of the men [7].
The impact of OAB on quality of life (QoL) is considerable: OAB contributes to QoL limitations and impairment of physical activity, psychological well-being, social and sexual activity, occupational productivity and domestic habits [6, 8].
The results of the NOBLE study suggest that some risk factors can be distinguished. Age, diabe-tes, urinary tract infections and early menopause have been suggested [6]. Moreover, in female, ab-dominal hysterectomy predisposes to urinary continence, especially the development of urge in-continence [5].In both sexes, obesity was reported as an important OAB risk factor [5, 8]. Most diet and lifestyle factors were not found to be associ-ated with OAB [5].
An important element of bladder anatomy and physiology is the bladder urothelium, composed of three cell layers. The basal cell layer (inner layer) contains cuboidal cells that coat the lamina propria. The intermediate layer is built up of “club-shaped” cells, while the superficial apical layer contains hexagonal cells called umbrella cells, covered by proteins called uroplakins. In the past, the urothe-lium was viewed simply as a barrier between urine and the detrusor muscle. Nowadays it is regarded as a structure involved in the micturition reflex, metabolic secretion and inflammatory regulation. The suburothelial localization of afferent endings makes the urothelium a highly specialized sensory site, reacting to thermal, mechanical and chemical stimuli, and a transducer, able to release various neurotransmitters and alter afferent excitability (Table 1).
Urothelial cells release adenosine triphosphate (ATP), acetylocholine (Ach), nitric oxide (NO), prostaglandins and substance P by a calcium-de-pendent mechanism, similar to neurotransmitter release from nerve endings. These chemicals allow reciprocal communication with neighboring cells and with sensory nerves and other cells located in the detrusor muscle. These findings are of special interest because they offer potential pharmacolog-ical targets for future OAB medpharmacolog-ical agents [9, 10]. The LUT is innervated by a complex of af-ferent and efaf-ferent neurons, including both sym-pathetic, parasymsym-pathetic, NANC and somatic pathways. Generalizing, the sympathetic nervous system nerves originating from spinal segments T11-L3 control the inter-micturition periods, leading to urethral sphincter closure and to de-trusor relaxation, enabling the bladder to fill. The parasympathetic nerves, arising from segments S2-S4, are responsible for the relaxation of urethral sphincter and simultaneous detrusor contraction during micturition. Somatic motoneurons come from spinal segments S2-S4, contributing to rest-ing pelvic floor muscle tone and controllrest-ing the external urinary sphincter. The LUT is also sup-plied with afferent nerves, which are an element of the voiding reflex and organize micturition. Nor-mal micturition occurs in response to an afferent signal from the LUT and is under the control of brain and spinal centers. A general conviction ex-ists that pontine circuits are responsible for the shift from the filling (storage) phase to the emp-tying (voiding) phase. In adults, voiding is under the influence of central nervous mechanisms, and therefore also under voluntary control [4, 11].
Normal voiding is preceded by bladder filling, which entails the accommodation of an increas-ing volume of urine with little or no change in intravesical pressure, with a closed sphincter and
the absence of involuntary bladder contractions. When the bladder is empty, its wall is folded and has high viscoelasticity, enabling it to preserve relatively low and constant pressure during the filling phase by expanding the smooth muscles and stretching the bladder. The progressive dis-tension of the bladder wall is the primary excita-tion that starts the micturiexcita-tion reflex. There are at least two afferents innervating the bladder: myeli-nated, mechanosensitive Aδ neurons, activated in response to both low (non-nociceptive) and high (nociceptive) pressures; and unmyelinated C-fi-bers, which respond to cold, heat or chemical irri-tation of bladder mucosa. Aδ fibers play the main physiological role in the voiding reflex, whereas C-fibers are regarded as primary nociceptive neu-rons and as the main fibers responsible for con-trollling micturition in fetuses, neonates and in adults with LUT damage. During bladder filling, when threshold pressure is achieved, afferent im-pulses conducted by the pelvic nerve are sent via dorsal root ganglia (DRG) and the spinal cord to the periaqueductal gray matter (PAG). This center plays a crucial role in the voiding reflex, communi-cating with the suprapontine regions. The central suprapontine voiding regulatory mechanisms are poorly understood, but some positron emission tomography (PET) studies suggest that the inferi-or frontal gyrus and anteriinferi-or cingulated gyrus are involved. These regions are parts of the limbic or emotional nervous system and are associated with the affective aspects of voiding. On the one hand, voluntary control over voiding is responsible for the motivation to empty the bladder before it be-comes overfilled; but on the other hand, is also responsible for making decisions to abstain from voiding if there are not social grounds to do so.
The ascendant PAG neurons project to the pontine tegmentum, where two different regions involved in the micturition reflex are located: the dorsomedially located M region (Barrington’s nu-cleus), which is also named the pontine micturition center (PMC), and the more laterally located L re-gion, which is regarded as a pontine storage center (PSC). Electrical stimulation of the PMC leads to urethral sphincter relaxation, following transient detrusor contraction; thus, the PMC organizes the normal voiding coordination of these structures. This results from the course of fibers originating from the PMC – some of them synapse in the sacral parasympathetic outflow spinal region, which pro-vides excitatory innervations to the detrusor. Other fibers conduct stimulatory signals to the urethral sphincter synapse on the sacral Onuf spinal nucle-us, causing the sphincter relaxation.
be-Table 1. Targeted receptors in the urothelium and on sensory neurons [9, 10] Tabela 1. Receptory i kanały jonowe urotelium oraz nerwów czuciowych będące potencjalnymi miejscami oddziaływania farmakologicznego [9, 10] Activator (Aktywator) Ion channel or receptor (Kanał jonowy lub receptor) Blocker (Bloker) Modulator (Modulator) Location (Umiejscowienie) Catecholamines (Katecholaminy) α – and β - subtypes urothelium nerve endings Acetylocholine (Acetylocholina) nicotinic muscarinic M1-M3 urothelium Nerve endings Adenosine triphosphate (ATP) purinergic receptors P2X, P2Y urothelium nerve endings Mechanical (Czynniki mechaniczne) Cold (Niska temperatura) ENaC amiloride aldosterone, CAP-1, Trypsin pH, Na
+, Ca
2+ urothelium nerve endings DRG Mechanical (Czynniki mechaniczne) Cold (Niska temperatura) Low pH (Niskie pH) ASIC amiloride Zn
2+, neuropeptide
FF
urothelium DRG detrusor
Heat – 43°C (Wysoka temperatura – do 43°C) Low pH (Niskie pH) Eicosanoids (Eikozanoidy) Arachidonic acid (Kwas arachidonowy) Anandamide (Anandamid) Capsaicin (Kapsaicyna) Resiniferatoxin (Resiniferatoksyna) TRPV1 capsazepine Na
+, Ca 2+, Mg 2+, adenosine
urothelium nerve
endings
DRG detrusor myofibroblasts
Noxious heat > 53°C (Wysoka uszkadzająca temperatura > 53°C) Mechanical (Czynniki mechaniczne) Growth factors (Czynniki wzrostowe) TRPV2
DRG urothelium? nerve
Table 1. Targeted receptors in the urothelium and on sensory neurons [9, 10] – cont. Tabela 1. Receptory i kanały jonowe urotelium oraz nerwów czuciowych będące potencjalnymi miejscami oddziaływania farmakologicznego [9, 10] – cd. Activator (Aktywator) Ion channel or receptor (Kanał jonowy lub receptor) Blocker (Bloker) Modulator (Modulator) Location (Umiejscowienie) Noxious cold (Niska uszkadzająca temperatura) Mechanical (Czynniki mechaniczne) Cinnamaldehyde (Aldehyd cynamonowy) Isothiocyanate (Izotiocyjanek) Garlic (Związki siarkowe izolowane z czosnku) Bradykinin (Bradykinina) TRPA1 Ca
2+, voltage
urothelium nerve endings DRG Cold (Niska temperatura) Menthol (Mentol) Icilin (Syntetyczny super agonista TRPM8-icilin) TRPM8 pH, Ca 2+ urothelium nerve endings DRG Cold (Niska temperatura) Heat (Wysoka temperatura) Mechanical (Czynniki mechaniczne) Voltage – leak current (Czynniki elektryczne) TREK-1 scorpion toxin arachidonic acid Na + ether halothane detrusor DRG ENaC – Epithelial Na
+ Channel.
CAP-1 – Channel Activating Protein-1. DRG – Dorsal Root Ganglion. ASIC – Acid-Sensing Ion Channels. TRPV – Transient Receptor Potential Vanilloid. TRPA – Transient Receptor Potential Ankyrin. TRPM – Transient Receptor Potential Melastatin. TREK – Tandem P domain weak inwardly rectifying K
+ channel
(TWIK).
–
related
K
+ channel
(a subfamily of the K2P channel. ENaC – nabłonkowy kanał sodowy. CAP-1 – białko aktywujące kanał. DRG – zwój korzeni grzbietowych nerwów rdzeniowych. ASIC – kanały jonowe niezależne od napięcia aktywowane protonami (niskim pH). TRPV – receptor waniloidowy z grupy receptorów przejściowego potencjału. TRPA – receptor ankirynowy z grupy receptorów przejściowego potencjału. TRPM – receptor melastatynowy z grupy receptorów przejściowego potencjału. TREK – słaby dokomórkowy prostowniczy kanał potasowy pobudzany wzrostem zewnątrzkomórkowego stężenia jonów K
+ (podrodzina
kanałów
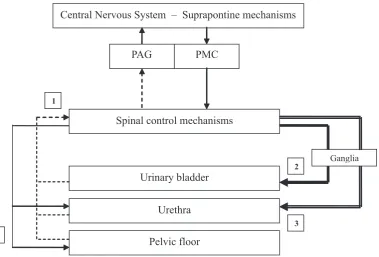
tween voiding, maintain tonic sphincter contrac-tion during periods of urine storage and relax the detrusor (see Figure 1) [4, 11, 12].
The final activation of muscarinic receptors results in bladder contraction. There are at least three subtypes of muscarinic receptors: M1, M2 and M3. M1 receptors appear to facilitate the fur-ther release of acetylocholine. M2 and M3 types act synergistically. The activation of M2 recep-tors, which predominate in the bladder, leads to inhibition of detrusor relaxation by diminishing sympathetic activity. This enhances the detrusor response to M3 receptor activation, although di-rect partial stimulation of bladder contraction by M2 subtypes also seems possible [4, 13]. Studies by Matsui et al. [14] and Stengel et al. [15] with M2 and M3 knockout mice demonstrated that M2 receptors also play a role in contraction, but not the most important one. In M2/M3 double
knockout animals, responses to muscarinic agents in vitro were eliminated, suggesting that these two receptors are involved in contraction. M3 subtypes are regarded to be essential in mediating cholin-ergic-induced contractions of the detrusor. It is generally believed that contraction of the detrusor smooth muscle is a consequence of the activation of M3 muscarinic receptors due to acetylocholine release from efferent endings. The results of stud-ies concentrating on the molecular mechanisms of muscarinic receptor activation in the bladder are also ambiguous. M3 receptors act by produc-ing inositol triphosphate (IP3) and diacyloglycerol (DAG) as second messengers, while M2 receptors are thought to mediate a reduction in cAMP level (thus, the effect is the opposite of beta-adrenergic relaxation). Recent studies suggest that after the administration of muscarinic receptor agonists, other pathways may be involved: inhibition of
po-Fig. 1. Storage reflexes: During bladder filling, increasing afferent activity (the dotted line; 1) occurs. There is no para-sympathetic efferent drive (the bold line; 2) organizing bladder contractions. The para-sympathetic outflow (the double line; 3) to the urethral smooth muscle and the somatic outflow (the single continuous line; 4) to urethral and pelvic floor striated muscles are increased, keeping the outflow region closed [4, 5]. Voiding reflexes: During bladder empty-ing, an increased parasympathetic efferent drive occurs (the bold line; 2), organizing bladder contractions. The sym-pathetic outflow (the double line; 3) to the urethral smooth muscle and the somatic outflow (the single continuous line; 4) to urethral and pelvic floor striated muscles are turned off, enabling the outflow region to relax [4, 5]
Ryc. 1. Okres międzymikcyjny: podczas wypełniania pęcherza dochodzi do wzrostu impulsacji aferentnej (przerywana linia; 1). W tym okresie brak eferentnej impulsacji parasympatycznej (gruba linia; 2), kontrolującej skurcze pęcherza. Aktywność włókien sympatycznych mięśniówki gładkiej cewki moczowej (podwójna linia; 3) oraz włókien somaty-cznych unerwiających mięśnie poprzecznie prążkowane cewki i dna miednicy (cienka linia; 4) jest zwiększona, co powoduje zamknięcie drogi odpływu moczu [4, 5]. Okres mikcyjny: podczas opróżniania pęcherza występuje wzrost eferentnej impulsacji parasympatycznej (gruba linia; 2), odpowiedzialnej za skurcze pęcherza. Dochodzi do zaha-mowania aktywności włókien sympatycznych mięśniówki gładkiej cewki moczowej (podwójna linia; 3) oraz włókien somatycznych unerwiających mięśnie poprzecznie prążkowane cewki i dna miednicy (cienka linia; 4), co umożliwia relaksację drogi odpływu moczu [4, 5]
Central Nervous System – Suprapontine mechanisms
PAG PMC
Spinal control mechanisms
Urinary bladder
Urethra
Ganglia 1
2
3
Pelvic floor
tassium channels or activation of calcium influx via L-type channels with subsequent activation of the Rho kinase system [13].
Additionally, spontaneous contractile activity in the bladder is observed, called “micromotions”. It was formerly believed that any bladder contrac-tions prior to voiding are inappropriate and are indicative of bladder overactivity. Nowadays it is agreed that a “normal” bladder exhibits some spontaneous activity, but also that an exaggeration of micromotions during filling-phase contractions may be attributed to detrusor overactivity. There are still questions about which cells generate spon-taneous phasic activity. Detrusor myocytes are known to be among the cells that have contractile activity. However, most studies indicate that inter-stitial myofibroblasts are pacemakers [13]. These cells are similar to the interstitial cells of Cajal, lo-cated in the digestive tract, which are responsible for its basic electrical rhythm, organizing motor activity. Other researchers favor the opinion that the interstitial cells in the detrusor modulate the spread of action potentials along the muscle bun-dles rather than being pacemakers of spontaneous activity [10]. In any case, there is no doubt that bladder micromotions are an important element in physiological voiding as well as being one of the possible elements of bladder overactivity [10, 13].
Summarizing, the efferent loop driving the nor-mal voiding reflex includes the PAG with the su-prapontine controlling centers, the M-region of the pontine micturition center, the lumbar and sacral spinal cord, the bladder and the urethra, with acti-vation of the M3/M2 muscarinic receptors.
Theories Regarding
the Pathophysiololgy
of Overactive Bladder
The pathophysiology of OAB may be neuro-genic, myogenic or – in the case of symptoms that cannot be explained – idiopathic.
The Neurogenic Theory
of OAB: Disturbances
of Central Mechanisms
Because the bladder is under nervous control, it is obvious that pathological changes affecting the central or peripheral nervous system can result in voiding disturbances. The mechanisms responsible for neurogenic OAB include decreased suprapon-tine inhibition of micturition, damage to the axonal paths in the spinal cord, a loss of peripheral
inhibi-tion or enhancement of excitatory neurotransmis-sion in the voiding reflex. Common disorders that are associated with OAB via these disturbances in-clude stroke, spinal cord injury, Parkinson’s disease (PD) or multiple sclerosis [8, 11].
The prevalence of post-stroke incontinence is from 57% to 83% [11, 16]. In patients with cor-tical lesions, urinary incontinence results from uninhibited detrusor contractions due to damage to the cerebral inhibitory centers, and does not involve detrusor-sphincter dyssynergia. Accord-ing to Sakakibara et al. [16], the anteromedial frontal lobe and its descending tonic inhibitory pathways are responsible for micturition dysfunc-tion in stroke patients. An animal model of stroke also confirmed that cortex ischemia plays a role in stroke-associated OAB: detrusor overactivity was observed in rats 30 minutes after experimental occlusion of the middle cerebral artery with sub-sequent putamen and cortex ischemia [17]. This pathomechanism of post-stroke OAB may be re-lated to several neurotransmitter disturbances, including glutamate (with NMDA receptors), do-pamine, GABA or nitric oxide (NO). It was found that overactivity can be reversed by NMDA recep-tor antagonists, selective D2 receprecep-tor blockers or GABA-ergic agents [11].
In spinal cord injury, urinary dysfunction is re-lated to the severity and area of the spinal cord af-fected by the disease. OAB symptoms develop days to weeks after spinal damage, and spinal lesions above the sacral level lead to detrusor overactivity. The most important role in this pathomechanism is apparently played by capsaicin-sensitive C-fi-bers that can fire at low intravesical pressures. The increased mechanosensitivity of C-fibers may arise from synaptic reorganization of the dorsal root ganglion (DRG) supplying the bladder. This phe-nomenon is manifested by neuron enlargement and electrical instability, and a high expression of low-threshold tetrodotoxin-sensitive sodium channels [11, 18].
detrusor overactivity may be related to a lack of D1 receptor activation [19, 20].
In multiple sclerosis, voiding disturbances are mostly due to spinal lesions, and arise from demy-elinating plaques of white matter, especially in the posterior and lateral columns of the cervical cord. As in PD patients, symptoms of voiding dysfunc-tion include frequency, urgency, incontinence and poor urinary stream [11, 21].
The neurotransmitter and receptor abnor-malities implicated in the central nervous system diseases mentioned above may be glutamatergic (both NMDA and AMPA), dopaminergic, cholin-ergic and abnormalities of the GABA system [11]. There are some studies that attach importance to serotoninergic abnormalities, because there is a link between decreased serotonin levels (depres-sion, anxiety) and OAB occurrence [11].
Additionally, serotonin reuptake inhibitors and selective 5HT1A/5HT2 receptor agonists have been found to improve nervous control of the ure-thral outlet [11].
The Neurogenic Theory of OAB:
Disturbances of Peripheral
Afferent Control
In current OAB pathophysiology, a great deal of attention is devoted to afferent elements of the micturition arc. As described above, mechanical stimuli trigger myelinated Aδ fibers and transmit impulses via the sacral spinal cord to the brain-stem and then to periaqueductal gray matter, ter-minating in the pontine micturition center. Un-myelinated C-fibers appear to be responsible for signalling irritation and noxious stimuli, and are also capable of triggering the voiding reflex.
In OAB, interruption of descending spinal pathways leads to a reorganization of afferent fibers, and the C-fibers become predominant. Sensitization of these fibers increases bladder ex-citability and lowers the pain threshold. This is supported by the desensitizing effect of capsaicin. This compound is neurotoxic, and when adminis-tered intravesically, it ameliorates OAB symptoms without impairing normal micturition, proving that C-fibers are involved in bladder overreactiv-ity. In subcutaneous administration in rats, cap-saicin also increased the volume threshold for ini-tiating voiding [22].Resiniferatoxin is considered 1000 times more potent an agent than capsaicin, and is able to develop C-fiber desensitization in lower concentrations that do not produce noxious side effects. It has been stated that resiniferatox-in stimulates the vanilloid receptors and resiniferatox-inhibits tachykinin release [22]. These local mechanisms
seem interesting in terms of future pharmacologi-cal studies seeking agents to ameliorate OAB.
Several other factors may be related to neu-rogenic OAB pathomechanisms associated with disturbed afferents activity, such as bradykinin, histamine, nitric oxide (NO) or substance P (SP), vasoactive intestinal peptide (VIP), calcitonin-gene related peptide (CGRP) and other peptides. There is evidence that chronic inflammatory blad-der is characterized by overexpression of nitric oxide synthase and NO release from capsaicin-sensitive afferents [22, 23]. NO is a molecule that relaxes the smooth muscles around the bladder outlet and thus contributes to the urethral pres-sure drops that precede involuntary bladder con-traction. VIP seems to be an inhibitory agent in efferent parasympathetic pathways and – along with SP – an excitatory factor in bladder afferents. It has been discovered that the VIP concentration in overactive bladder smooth muscles is reduced when compared to a normal bladder, leading to the conclusion that a loss of the inhibitory impact of VIP may promote bladder overactivity. These findings are also promising for future therapies to ameliorate OAB [22, 23].
The role of calcium influx in unstable myocytes is also emphasized in current OAB pathophysiolo-gy. In addition to the neurotransmitter disturbanc-es mentioned above, poor calcium ion regulation has been proposed as another potential mechanism leading to increased cells depolarization and de-trusor instability. Verapamil was the first calcium channel blocker that was shownto reduce detrusor contractility and increase the intravesical pressure at which micturition is induced, although potential cardiovascular side effects limit the usage of vera-pamil as an OAB drug [24]. Another effective agent in calcium channels blocking is propiverine, which also has antimuscarinic properties, inhibiting blad-der overactivity in rats more effectively than oxybu-tynin, the classic cholinolytic [25]. Agents affecting calcium influx are also expected to be introduced in future OAB treatment.
OAB may be also related to an exaggerated re-sponse to muscarinic agents. In vitro, these agents provoke a greater bladder response in comparison to normal detrusor muscle, probably suggesting denervation. Although it has not been established for all types of OAB, in some conditions associated with bladder outlet obstruction, enhanced sensi-tivity to cholinergic agents was found [13].
changes in the detrusor smooth muscle may be a key factor in the development of OAB [22]. The normal bladder has high compliance during the filling phase, while an overactive one exhibits some structural changes when observed under micro-scope. An increased density of elastin and collagen within smooth muscle bundles has been reported as a change that may affect electrical conduction and reduce compliance and emptying. A clinical entity associated with bladder wall reconstruction and its reduced compliance is bladder outflow ob-struction. In an experimental rat model of bladder outflow obstruction, detrusor muscle hypertrophy, spontaneous electrical activity, partial denervation and abnormal cell junction were observed. Ob-struction has also been shown to increase spinal reflexes involving C-fibers [22]. Elbadawi et al. [26] studied structural abnormalities in geriatric pa-tients with OAB, using electron microscopy. They found that overactive detrusor muscle contained widened intercellular spaces, an abundance of pro-trusion junctions and ultraclose abutments, which may result in a “chainlike linkage” of cells. This finding corresponds to the theory of abnormal electrical cell excitation. In the normal bladder, the foci of electrical activity are spread across a small number of coupled neighbouring cells. This means that, physiologically, excitatory nerve input must be delivered to a large number of smooth cells to trigger an organized bladder contraction. In OAB muscle, dysfunctional electrical coupling between smooth muscle cells enables small foci of electri-cal impulses to be propagated beyond their normal range. As a consequence, it can be concluded that the normal bladder is characterized by scattered contractions which appear locally, while in an overactive bladder these local activities are more prone to be global. The electrical abnormalities may be due to the distinctive structural changes mentioned above [13, 22].
The Autonomous Bladder
Theory
There is also a relatively new hypothesis that posits that the detrusor is modular – made up of distinct separate areas of muscle. Each module is supplied by one intramural ganglion or by a node of interstitial cells called a myovesical plexus. Dur-ing normal bladder functionDur-ing, autonomous electrical activity of various modules is observed, which does not lead to micturition contractions. In pathological conditions there can be a synchro-nization of activity between modules, which may be propagated through a network of intramural ganglia or myovesical plexuses. These changes
may result in excessive excitatory input or a fail-ure of inhibitory input, thus predisposing to blad-der overreactivity. Obviously, one can classify this hypothesis as special kind of myogenic OAB the-ory, although it is less specific with respect to the mechanism [3, 27–29].
A Unifying Theory:
A Compromise View
The most widely accepted theories of the pathophysiology of OAB are the neurogenic and myogenic ones. They are not mutually exclusive, and can be regarded as complementary. The basic mechanisms underlying these two concepts inter-act to produce the clinical manifestation of overac-tive bladder [22]. It remains uncertain whether it is neurogenic or myogenic disturbances that initiate involuntary detrusor activity and affect the second component. For example, smooth muscle changes may be responsible for promoting the pathologi-cal spread of electripathologi-cal activity and inducing the emergence of pathological nerve pathways. This is suspected to be mediated by nerve growth factor (NFG), which is synthesized by the structurally al-tered bladder wall. NFG concentration was signifi-cantly higher in specimens of overactive bladders than in normal ones; however, the role of this mol-ecule in OAB pathomechanisms is unknown so far and requires further study [30, 31]. On the other hand, neurologic changes (sensitization of periph-eral afferent pathways with enhanced neuroendo-crine activity of these fibers, and reduced activity of central inhibitory nerves supplying the muscles) may contribute to alternations in the sensitivity of the detrusor. Thus, determining which changes are the cause and which are a secondary consequence requires further study [22, 32].
Undoubtedly, further progress in understand-ing the pathophysiology of overactive bladder, with a focus on the various neurotransmitters contributing to disturbed central and peripheral innervation, is a promising area of research for fu-ture OAB pharmacotherapy.
Conclusions
References
Abrams P, Cardozo L, Fall M, Griffiths D, Rosier P, Ulmstein U, van Kerrebroeck P, Victor A, Wein A:
[1] The
standardisation of terminology of lower urinary tract function: report from the Standardisation Sub-committee of the International Continence Society. Neurourol Urodyn 2002, 21, 167–178.
Abrams P, Artibani W, Cardozo L, Dmochowski R, van Kerrebroeck P, Sand P:
[2] Reviewing the ICS 2002
termi-nology report: The ongoing debate. Neurourol Urodyn 2006, 25, 293. Hashim H, Abrams P:
[3] Overactive bladder: an update. Curr Opin Urol 2007 17, 231–236. Chu FM, Dmochowski R:
[4] Pathophysiology of overactive bladder. Am J Med 2006, 119, 3S–8S. Tubaro A, Palleschi G:
[5] Overactive bladder: epidemiology and social impact. Curr Opin Obstet Gynecol 2005, 17, 507–511.
Freeman RM, Adekanmi OA:
[6] Overactive bladder. Baillieres Best Pract Res Clin Obstet Gynaecol 2005, 19, 829–841. Irwin DE, Milsom I, Hunskaar S, Reilly K, Kopp Z, Herschorn S, Coyne K, Kelleher C, Hampel C, Artibani W, [7]
Abrams P: Population-based survey of urinary incontinence, overactive bladder and other lower urinary tract symptoms in five countries: results of the EPIC study. Eur Urol 2006, 50, 1306–1314.
Wein AJ, Rackley RR:
[8] Overactive bladder: a better understanding of pathophysiology, diagnosis and manage-ment. J Urol 2006, 175, S5–S10.
Moore CK, Goldman HB:
[9] The bladder epithelium and overactive bladder: what we know. Curr Urol Rep 2006, 7, 447–449.
Araki I, Du S, Kobayashi H, Sawada N, Mochizuki T, Zakoji H, Takeda M:
[10] Roles of mechanosensitive ion
chan-nels in bladder sensory trasduction and overactive bladder. Int J Urol 2008, 15, 681–687. Andersson KE, Pehrson R:
[11] CNS involvement in overactive bladder. Pathophysiology and opportunities for phar-macological intervention. Drugs 2003, 63, 2595–2611.
Griffiths DJ:
[12] Cerebral control of bladder function. Curr Urol Rep 2004, 5, 348–352. Kumar V, Cross RL, Chess-Williams R, Chapple CR:
[13] Recent advances in basic science for overactive bladder. Curr Opin Urol 2005, 15, 222–226.
Matsuni M, Motomura D, Karasawa H, Fujikawa T, Jiang J, Komiya Y, Takahashi S, Taketo MM:
[14] Multiple
functional defects in peripheral autonomic organs in mice lacking muscarinic acetylocholine receptor gene for the M3 subtype. Proc Natl Acad Sci USA 2000, 97, 9579–9584.
Stengel PW, Gomeza J, Wess J, Cohen ML:
[15] M2 and M4 receptor knockout mice: muscarinic receptor function in cardiac and smooth muscle in vitro. J Pharmacol Exp Ther 2000, 292, 877–885.
Sakakibara R, Hattori T, Yasuda K, Yamanishi T:
[16] Micturitional disturbance after acute hemispheric stroke: analysis of the lesions site by CT and MRI. J Neurol Sci 1996, 137, 47–56.
Kaidoh K, Igawa Y, Takeda H, Yamazaki Y, Akahane S, Miyata H, Ajisawa Y, Nishizawa O, Anderssen KE: [17]
Effects of selective beta 2 and beta 3 – adrenoceptor agonists on detrusor hyperreflexia in conscious cerebral infarcted rats. J Urol 2002, 168, 1247–1252.
Shefchyk SJ:
[18] Spinal cord neural organization controlling the urinary bladder and striated sphincter. Prog Brain Res 2002, 137, 71–82.
Albanese A, Jenner P, Marsden CD, Stephenson JD:
[19] Bladder hyperreflexia induced in marmosets by 1-methyl-4-phenyl-1,2,3,6-tetrahydropiridine. Neurosci Lett 1988, 87, 46–50.
Seki S, Igawa Y, Kaidoh K, Ishizuka O, Nishizawa O, Andersson KE:
[20] Role of dopamine D1 and D2 receptors in
the micturition reflex in conscious rats. Neurourol Urodyn 2001, 20, 105–113. Fernandez O:
[21] Mechanisms and current treatment of urogenital dysfunction in multiple sclerosis. J Neurol 2002, 249, 1–8.
Goldberg RP, Sand PK:
[22] Pathophysiology of the overactive bladder. Clin Obstet Gynecol 2002, 45, 182–192. Dmochowski RR:
[23] The puzzle of overactive bladder: controversies, inconsistencies and insights. Int Urogynecol J 2006, 17, 650–658.
Saito M, Kondo A:
[24] Effects of verapamil on bladder instability induced by partial outflow obstruction in rat. Int Urol Nephrol 1998, 30, 543–555.
Kwak II, Lee JG:
[25] Inhibitory effects of propiverine, atropine and oxybutynin on bladder instability in rats with infravesical outlet obstruction. Br J Urol 1998, 82, 272–277.
Elbadawi A:
[26] Pathology and pathophysiology of detrusor incontinence. Urol Clin North Am 1995, 22, 499–512. Drake MJ, Mills IW, Gillespie JI:
[27] Model of peripheral autonomous modules and a myovesical plexus in normal and overactive bladder function. Lancet 2001, 358, 401–403.
Drake MJ:
[28] The integrative physiology of the bladder. Ann R Coll Surg Engl 2007, 89, 580–585. Drake MJ, Harvey IJ, Gillespie JI:
[29] Autonomous activity in the isolated guinea pig bladder. Exp Physiol 2003, 88, 19–30.
Dupont MC, Persson K, Spitsbergen J, Tuttle JB, Steers WD:
[30] The neuronal response to bladder outlet obstruc-tion, a role for NGF. Adv Exp Med Biol 1995, 385, 41–54.
Tanner R, Chambers P, Khadra MH, Gillespie JI:
[31] The production of nerve growth factor by human bladder smooth muscle cells in vivo and in vitro. BJU Int 2000, 85, 1115–1119.
Brading A:
Address for correspondence:
Łukasz Dobrek
Department of Pathophysiology Jagiellonian University Medical College Czysta 18
31-121 Kraków Poland
E-mail: [email protected]
Conflict of interest: None declared