Factors Affecting Bacterial Competence
for
Transfection
and
Transfection
Enhancement
H. T. EPSTEIN
Departmenztof Biology, Brandeis University, Waltham, Massachusetts02154
INTRODUCTION... 313
MATERIALS AND METHODS... 314
RESULTS... 314
DiSCUSSION... 317
LITERATURE CITED... 318
INTRODUCTION
There is at present no real insight into the physiological nature of the competent state of bacteria. Indeed, it is not even known whether there is asingle competent state or whether
com-petency depends on factors otherthan bacterial properties. Accordingly, it appeared desirableto study competency for two different deoxyribo-nucleic acid (DNA) preparations in the same kind of competent cells. This can be done by studying transformation and transfection in one bacterial species or by studying two kinds of transformationoroftransfection inonebacterial species. Bacillus subtilis exhibitscompetenciesfor transformationandfortwokinds of transfection. Thepresent work is a
comparative
study of the two kinds of transfection.Transfection in B. subtilis is characterized (7, 15-17) by two different dependencies on phage DNA concentration. For example, SP02 trans-fectslinearly with DNAconcentration (15), and SP82 transfectsasthethirdorfourthpowerof the DNA concentration (7). For
SP82,
Green(7-9)
hassupplied evidencethat thenonlinear classre-sults frominactivation of
incoming
phageDNA,
whichmakesgenetic
recombination necessaryfor the production ofreplicating phage
molecules. SP82DNA,moreover,differs from host DNA in that it containshydroxymethyluracil
inplace
of thymine [Kahan and Kahan,quoted
by
Green (7)].Thus,it is"foreign"DNAand wouldbeex-pected to be attacked
by
defensive cellularnu-cleases.
Experiment andexpectation agree in the inac-tivationof the SP82 DNA. It
is,
then,surprising
that the
linearly
transfecting
SPO2 DNAactually
(15) transfects lessefficiently
permicrogram
of DNA than does SP82 DNA. Further, SP02 DNAhas amolecularweightapproximately
25% of that of SP82 DNA,judged
fromX-ray
inac-tivation curves obtained in our laboratory. One would expect this smaller DNA to be extracted and purified in appreciably less damaged form than the DNA of SP82. SPO2 DNA is also likely
to behomologous to host DNA, because SPO2
appears to be a temperate phage (15) and
be-causeSPO2 DNA and host DNA have thesame
buoyant density in CsCl (15). In contrast tothe
expectations raised by this comparison of SPO2 DNAandSP82 DNA,onefindsthat, in the same batch of competent cells,alessdamaged and
ap-parently homologous DNA transfecting linearly with DNAconcentration ismarkedlyless efficient in transfection than a more damaged,
heterolo-gous DNA requiring genetic recombination for plaque formation.
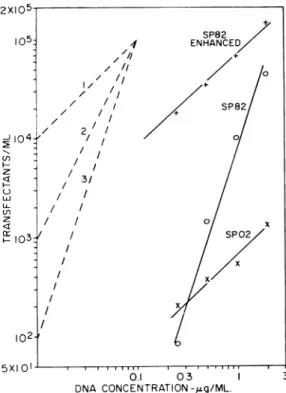
The situation is presented visually in Fig. 1, whichalso shows theenhancement and lineariza-tion of SP82 transfeclineariza-tion as a result of pre-ex-posure ofcompetent cells to ultraviolet (UV)-ir-radiated Escherichia coli DNA(5).After enhance-ment, the linear SP82 transfection is 100-fold
greaterthanSP02 transfection.
Thereare twoquite different kinds of explana-tions for the low SP02 DNA transfection: (i) SP02 may replicate poorly in competent host cells or (ii) lysogenization may have occurred with theconsequent loss of infective centers.
The first explanation starts from the above-mentioned observation that SPO2 DNA is homologoustohostDNAby thebuoyantdensity criterion.Itis alsoknown(3, 13) that DNA syn-thesisisarrested incompetent cells. Presumably, there hasbeen a sequestering of someenzyme(s) needed for DNA synthesis. Because of the ho-mologyofSPO2DNA to hostDNA, there may be a decreased opportunity for replication of SPO2DNA,whichneedstousehostenzymes for such purposes. During the period of arrested DNA synthesis,the SP02 DNA could beslowly degraded by host nucleases, thereby decreasing 313
BACTERIOL. REV. '4
,/A
//1
/ //,
///
I
//
/ 2 1 / 3/ /I /3/ A I tJ-SP82 ENHANCED//
E104-,
2I
-~ z Ul (n z z W- 103: 102-5XIOI I .. O. 03 DNA CONCENTRATION-/zg/ML.FIG. 1. Dependence of plaque formation on
concen-tration ofSP02 and SP82 DNA. Also shown isthe SP82 transfection enhancement by 1.5 ,ug of
Escher-ichiiacoliDNA givenabout104 ergs/mm2ofultraviolet
irradiationfromasterilamp.Thedashedlinesshowthe theoretical first, second, and thirdpowerdependencies onDNA conicenitration.
theeffectivetransfecting capacity. The sequester-ingcould belargely irrelevanttoSP82DNA
be-cause this large phage has the same molecular
weight of DNA as coliphage T4 (7), which
carries or induces enzymes essential for its own
replication, especially its own DNA polymerase
andligase (1, 6;B.Weiss, T. R. Live,and C. C. Richardson, Federation Proc., 62:395, 1967). Another result relevant to the experiments to
be described is the finding (7, 17) that
recom-bination frequencies are much higher for
trans-fection crosses than for the same crosses done with intactphages.Thiscould betheconsequence of at least two entirely different mechanisms.
First, as set forth above, competent cells may contain increased levels of nucleases attacking foreign DNA such as the
hydroxymethyluracil-containingDNAfromphageSP82.Second,
trans-fectionsareusuallydonewithaDNA
concentra-tion (1 to 2,ug/ml) equivalentto 25to 50
mole-cules of intactDNAper cell. Ifthisvaluereflects
DNAuptakeinthecompetentcellsin the culture,
transfection correspondstowhat maybetermed
an"instant pool" of phage DNA molecules. It is known that phage recombination frequencies increase with multiplicity of infection (14) and with time in thelatentperiod (2, 12).
The postulated increase in nuclease attack on
SP82 DNA should result in transfections which decrease in levelwith thelength oftime the pre-competent cells have grown in competency
me-dium. This conclusion isatvariance with the fact that,duringthatgrowth period, transfection first increases to a maximumbeforedecreasing. There-fore, there must beafactorwhich increases
com-petence atearlytimes. Theliteraturecontainstwo findings which may explain the increase in com-petency. First,there may be an increase inability to fix DNA irreversibly, since competency gen-erally followstheabilitytofix DNA(11).
Second,
Wolstenholme etal.(19) recentlyshowedthatthe number ofmesosomal sites per cell increases as
competency develops in B. subtilis. In addition, they showed an association of radioactively la-beled DNA with these sites. If mesosomal sites
are thus identified with DNA-replicating
sites,
incoming DNA should find aprogressively
in-creasing chance of becoming associated with areplicating siteas thenumber ofmesosomalsites increases. Then, once the number of sites has reached itsmaximum,competenceshouldremain constantunless it decreasesdue to nuclease attack
ontheincomingDNA.
MATERIALS AND METHODS
Allmediaused and themethods ofpreparation of phages, competent bacteria, and the various types of DNA have beendescribed in detail pre-viously (4, 5). The bacterium used in all experi-ments was B.subtilis strain SB1 (requiring histi-dine and
tryptophan).
Competent cells were obtained by diluting frozenprecompetentcells10-fold intocompetency medium and incubating them with shaking for various times. These cells are described as, e.g., 60-min competent cells if the incubation lasted
60 min. Transfections were generally done by adding0.1 mlof DNA solutionto0.5mlof
com-petent cells. Assays for plaque formation were
doneafter50minofincubationat37 C.
DNA concentration in purified samples was
determined bymultiplying the optical density at
260 nmby 50toobtaintheresult inmicrograms per milliliter. DNA in nonpurified samples was
determinedbytheCeriotti method, with purified SP82DNA as thestandard.
RESULTS
Theinitialstudieswereof thetwoexplanations suggested for the low level ofSPO2transfection:
ovi|nts
I* I, Ir, I, I
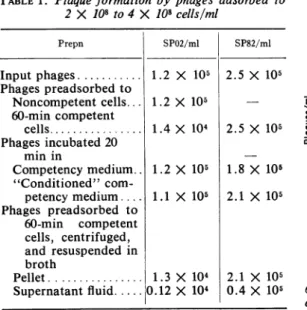
TABLE1. Plaqueformation byphages adsorbed to 2 X 108 to4X 108cells/ml Prepn SP02/ml Input phages... 1.2 X 105 Phages preadsorbedto Noncompetent cells... 60-min competent cells... Phagesincubated 20 min in Competencymedium.. "Conditioned" com-petencymedium.... Phages preadsorbed to 60-min competent cells, centrifuged, and resuspended in broth Pellet... Supernatantfluid... 1.2 X 105 1.4 X 104 1.2 1.1 X 105 X 105 1.3 X 104 0.12X 104 SP82/ml 2.5 X 105 1500F 1250 - InputPhages 1000 E 750 2.5 X 105 a, 1L 1.8 X 105 2.1 X 105 500 250[
K
0~~~~~~ 0-
<~
~~~~
* * 2 4 6 8 10 12 14 16Incubation Time (min)
2.1 X 105 FIG. 2. Plaqueformerspresent in a suspension of
0.4 X 105 60-min competent cells at various timesafteraddition
ofabout 103SP02phages.
(i) poor replication of SPO2 in SB1 due to
se-questering of enzymes associated with synthesis of host DNA and(ii) loss of infectivecenters
be-causeoflysogenizationbySPO2.
The first experiment simply measured the efficiency ofplating after adsorption of phages
on broth-grown SB1 cells and on precompetent SB1 cells grown in competency medium for various times.To1-mlamountsof suchcells were
added0.1-mlamountsofaphagesuspension
con-taining about 108 phages/ml. After 20 min for adsorption, the suspensions were diluted and
assayed for plaque-forming units. Table 1
con-tains datafromtypicalexperiments. SP82 phages had thesametiter whetherpreadsorbedto60-min
competent cells or plated directly on seed cells.
However, SPO2 phages had a 9-fold lower titer
when preadsorbed to 60-min competent cells
than when plated directly on seed cells. Similar
experimentsshowed thatSP02 plaque formation dropped from about 50% when preadsorbed to 0-min competent cells to 17% on 30-min cells,
11
%7o
on50-mincells, and5%on80-min cells.Controlsshown inTable1 weredonetocheck
adsorption and the effect of incubating phages in fresh competency medium and in "condi-tioned" competency medium (that remaining after SB1 cellshadbeen madecompetenttherein
for 90min,thecellsthenhavingbeenremovedby
centrifugation).
The kinetics of the loss of plaque-forming ability weremeasured by plating portions ofan
SPO2phage-infectedculture of60-mincompetent cells. Theresults of such an experiment (Fig. 2)
show that a very rapid loss of plaque-forming ability occurred in the first 2 minafter the cells
andphagesweremixed.
Ifthe loss of infective centers is due to lyso-genization, thebacteriashould survive the
infec-tion. At low multiplicities ofinfection, the
sur-viving cells cannot be measured directly, but at multiplicities somewhat greaterthan unity there should be a measurable increase in number of
survivors above those killed bytheadsorptionof
phages.
Table 2 shows the results ofanexperiment in which 75-min competent cells were incubated
withvariousmultiplicitiesofSPO2phagesfor 20
TABLE2. Bacterial and SP02 phage survivalin infected75-min competent cells
(A) (C) (D) (E)
(F)(G
Experimental (B) Expected Experimental Input Plaques(G)
infection e-(A) bacteria bacteria put foues (F)/(E)
multiplicity surviving surviving phages found
3.7 0.025 0.13 X 108 0.133 X 108 19.2 X 108 1.3 X 108 0.067
1.23 0.294 1.52 X 108 2.3 X 108 6.4 X 108 0.2 X 108 0.031
0.37 0.692 3.6 X 108 4.5 X 108 1.92 X 108 0.08 X 108 0.042
BACTERIOL. REV. min before being platedfor plaque formation and
for colony formation. Adsorption of phages by such competent cells was always rapid and was
essentially 100%/- within 5 min. At a multiplicity of3.7, theexpected number ofsurvivingbacteria wasequaltotheexperimentalnumber,whichwas
0.025 of the starting number of bacteria. The efficiencyofplatingof theinputphageswas0.067, in agreement with figurespresented earlier.If the missing phages had lysogenized thebacteria, the number ofsurviving cells would havegreatly
ex-ceeded the number of cells actually found. A similar conclusion may be drawn from the data obtained with a multiplicity of 1.23. At lower multiplicities, the effect could not be detected
evenif itexisted,but the lossof
plaque
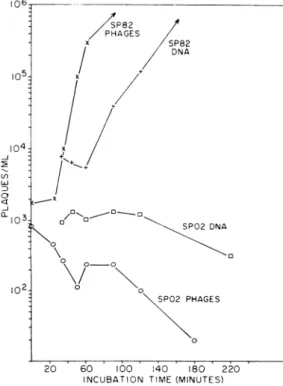
formation by theinput phageswasroughly constant for all multiplicities.Thepostulatedsequestering of DNAsynthesis enzymes was next tested by obtaining one-step growth curves forthetwophagestrains, both by infection with viable phages and by transfection with phage DNA (Fig. 3). A
typical
curve wasobtained for SP82 phages adsorbed on 90-min
106
I
\1~1
0 0o0/
\
3P82 DNA SP02 DNA 0SP02 PHAGES SP02 PHAGES 0 20 60 100 140 180 220 INCUBATION TIME(MINUTES)FIG. 3. Plaque -formerspresent ina suspensionof
90-min competentcells atvarious times after addition ofSP02 or SP82phlagesortheirDNA. At 20 min
after1,ugof eachtypeofDNAwasadded,further DNA
uptake wasstopped by additiont of2,ug of deoxyribo-nuclease.
competent cells. The latent period of about 40 min was the same as that found by Green (7), and in our laboratory, for infection of broth-grown cells. The burst size was more than 100. SP82 DNA transfection yielded a one-step growth
curve similar to that for phage infection except for theincreased latentperiodtypically found for transfection (7, 18). However, after exposure of 90-min competent cells to SPO2 phages or SPO2 DNA, there was noincrease in plaque formation for at least 3 hr after infection was initiated. In our laboratory, the latent period for SPO2 phage infection of broth-grown cells was about 32 min, with a burst size of about 100. A single experi-ment on SPO2 phage infection of 0-min compe-tentcells also gave no burst for more than 3 hr. The slow decrease of SPO2 plaque formation indicates a slow inactivation of the phage DNA. The lysogenization interpretation was further tested by obtaining growth curves for SPO2 phages and SPO2 DNA on competent cells which had beenirradiated for 3 min, a UVdose which maximizes transfection enhancement for phage SP82 (4). Iflysogenization werethecauseof loss of infective centers, the irradiation might be ex-pected to inhibit lysogenization and drive the phages into the lytic cycle. However, growth curveson such irradiated cells showed no trans-fection enhancement and no increase in plaque formation for at least 3 hr.
Itseemsprobable that thereis a sequestering of enzymes needed for SPO2 growth in competent cells, and that formation ofthe needed enzymes isinduced by SP82 phages or phage DNA infect-ingsamplesof the same competent cells.
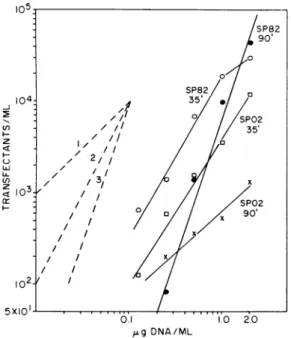
Evidencewasnextsoughtfor thehypothesized nucleases attacking the incoming transfecting DNA. If the enzyme level changes, the extent of inactivation of the DNA should change corre-spondingly. This deduction was tested by meas-uring the number of transfectants as a function of the phage DNA concentration. Transfecting
DNAwasadded tocellsafter 35 min and after 80
to 90min of incubation in competency medium. Plaqueformationwasassayedafteranadditional
50minofincubation.
As shown in Fig. 4, SP82 DNA transfection hadagreaterdependenceonDNAconcentration
on 90-min competent cells than on 35-min com-petentcells.Therefore,therewas agreaterlevel of nucleases at thelater time.
SPO2 DNA transfection showed the opposite pattern: greater transfection dependence on
35-min competent cells. Plaque formation was nearly linearonthe90-min competentcells, indi-cating that there was little attackontheDNA at
thattime.Presumably,then,at35minanenzyme
ispresent which can attack SPO2 DNA, and this
316
EPSTEIN ,1 a) D 1023 lo2: *IIs /0"0-- 0° 0}s9DNA/ML
FIG. 4. Dependence of transfection on DNA
con-centrationfor phages SP02 andSP82 incubated witht
35- anid 90-min competent cells. Dashled lines as in
legend forFig.1.
enzyme is subsequently sequestered along with
theDNA-synthesizing enzymes, as would be
ex-pected for cells preparing to take in DNA ho-mologous tothat ofthe host.
The 10-fold higher level ofSP02 transfection onthe35-min competentcellsindicatesanearlier development of maximal competency for this phage DNA. This point was studied directly by measuring the transfection with 1 ,ug of phage
DNA on cells incubated for increasingtimes in the competencymedium. The cells wereexposed
for50minbeforeplaque formationwas assayed. From the resultsof such experiments (Fig. 5), it is obvious that SP02 transfection peaks earlier
than that of SP82.Thus,whenSP02transfection is measured at thetime of maximal SP82 trans-fection,theSP02 transfectionlevel isdepressed. The SP02data inFig. 1 are, sotospeak,
meas-urementstakenatthe"wrong"time. DISCUSSION
The heuristicmodelunderlyingtheexperiments presented hereincharacterizes growth ofcells in
precompetency media as making the cells, in
essence, diploid.To be abletoincorporateextra DNA, thesecellshave to makeroom for the in-coming DNA, and they accomplishthis by
halt-ingDNA synthesis forone cell division. Thisis
achieved for B. subtilis by the amino acid step
down involved in diluting precompetency cells
104
Io-n
C:)
I{' '1lo
20 40 60 80 100 120
TIME OF DNAADDITION (MINUTES)
FIG. 5. Twenty-minute pulses oftransfectingDNA were given to cells at various times afterprecompetent cells were diluted 1:10 into competency medium. A
l-,Igamountof SP02 DNA orSP82DNA was added
to 1 ml of cells, and 2
4.g
ofdeoxyribonuclease was added 20 min later. The transfected cells were incu-batedfora totalof 50 miii before being dilutedand platedforplaqueformation.into competency medium. Among the missing amino acids are those
associaaed
with cell wall synthesis.The result ofthesemetabolic derangements is that there is anabilitytofixDNAand a place to putit afterentry.If theincoming DNA is an intact cell molecule, itcan go to a freereplicating site. If the DNAis notintact,thecells canincorporate someofthe DNAbythe process known as trans-formation. As this incorporated DNA is ho-mologous to host DNA, it cannot immediately bereplicated becausethe arrestofDNAsynthesis was accomplished by sequestering needed en-zymes.Phagesbringinginheterologous DNA can effectsynthesisoftheneededenzymes if the DNA is that of thelarge virulent phages such as SP82.
Twoexplanations of the low SP02transfection
wereput forth: poor growth on competent cells andlysogenization ofthe hostcells.Theevidence ofthe studies is that there is little, if any, lyso-genization, and that there is a sequestering of DNA synthesis enzymes. Although the present work has focused on DNA polymerase as the likely missing enzyme, the work of Kammen et al. (10) makes it likelythat the ligase (6; Weiss
0103 0 z-~0 0102 z
;
:,_ _ _6~ -o -oBACTERIOL. REV. etal., Federation Proc., 26:395, 1967.) isalso not
functioning, thus leading to the possibility of a coordinated repression of both enzymes. The high recombination frequencies for SP82 trans-fection crosses canbe accounted for by the high nuclease levels indicated by the experiments in Fig.4.Thedata inFig.4also permit theinference that replicatingsites are involved in competency becausetheriseincompetencyfor SP82DNAand SP02 DNA is the same through about 60 min.
Thus,
there seems to be a commonfactorwhich would likelybe one associated with cellproperties because the phages are so different from each other. One obvious cellproperty
isthe numberof replicating sites. Support for this interpretation hasbeenobtained by measuringthe UVcapacity ofcompetent cells. The capacity of90-min com-petent cells is several times that of 0-min com-petent cells.Itshould also be noted that Table 1 showed a 10%efficiencyofplatingSP02phages on60-min competent cells. Therefore, the "true" transfec-tionlevel for SPO2DNAcould be 10 timeshigher thanthatfound,orcloseto 105/ml. Thisvalue is similar to that for SP82 transfection, indicating that it is the enzyme sequestering which is re-sponsible forthelowSP02 transfection level.
Combining the results of Fig. 4and 5 permits the conclusion that thetime for maximal trans-fection for SP82 would be less than 90 min, if measuredwith 0.2 ,ug ofDNAper ml, compared with 90 min when measured with 2 ,ug/ml. The net result depends on DNA fixation, amount of DNA availableto recombine, and levels of nu-cleases. Thus, theterm "competency"appears to be too
general
since its expression dependsonat leastonefactorextraneoustothebacterium itself. The experiments with intact phages were pre-sumed to involve attachment to representative cells in the competent culture, of which only about 0.1%areactually transfectable. As shown inTable 2, thispresumption
has been tested by measuring theefficiency
of plating of SPO2 phagesathighmultiplicities;
similarlow efficien-cieswereobtained.Thus,the enzymesequestering must beoccurringinallcells in the culture, not only in the transfectable ones. The most likely reasonfor theinability ofthe vastmajorityof the cellsto betransfected would be that DNA fixa-tion and enzymesequestering are notnecessarily coupled. However, it is possiblethat DNA fixa-tion is maximal atall
times in the competency mediumand thatnuclease levels are insufficiently reduced in most cells evenwhen DNA synthesis enzymes havealready been sequestered.Finally,the results ofpreliminary experiments ontheDNAper cellsupport the model set forth at thebeginningofthis discussion. The DNA per
colony-forming unit in 0- to 30-min competent cells is about twice that for 90-min competent cells. Thedifficulty in thesemeasurements is the making of an accurate determination of the number of cells in each colony-forming unit. Length distributions have been determined from micrographs, and there is no change in the aver-agelength, which corresponds to about one and one-halfofthesmallest units measured. Still, no conclusioncanbedrawn about the DNA percell until the length ofthe individual cells has been determined. The data of Wolstenholme et al. (19) permittheconclusion that broth-growncells and 90-min competent cells have similar dimensions, butthere is noinformation about the dimensions ofthe0-min cells. [Sincethepresentation of this paper, R. N.Singhand M. P.Pitale(J.Bacteriol., 95:864-866, 1968) havereportedthe existenceof uninucleate and binucleate populations in com-petent cultures of B. subtilis, the uninucleate cells mostlikelybeingtheonesactually
competent.]
ACKNOWLEDGMENTS
This investigation was supported by grant GB 4497 fromtheNational ScienceFoundation.
I amindebted toMarilynFlynnfor excellent
tech-nicalassistance.
LITERATURECITED
1. Aposhian, H. V., and A. Kornberg. 1962.
En-zymatic synthesis ofdeoxyribonucleic acid. IX. Thepolymeraseformedafter T2bacteriophage
infection of Escherichia coli: a new enzyme. J. Biol. Chem. 237:519-525.
2. Doermann, A. H. 1953. The vegetative state in thelifecycleofbacteriophage: evidencefor its
occurrence and its genetic characterization. Cold SpringHarborSymp. Quant. Biol. 18:3-11.
3. Ephrussi-Taylor, H., and B. A. Freed. 1964.
Incorporation ofthymidine and amino acids into deoxyribonucleic acid and acid-insoluble cell structures in pneumococcal cultures
syn-chronized for competence to transform. J. Bacteriol. 87:1211-1215.
4. Epstein, H. T. 1967. Transfection enhancement by ultraviolet light. Biochem. Biophys. Res.
Commun. 27:258-262.
5. Epstein,H.T., and I. Mahler. 1968. Mechanisms of enhancement of SP82 transfection. J. Virol.
2:710-715.
6. Gefter, M. L., A. Becker, andJ. Hurwitz. 1967. Theenzymatic repair of DNA. 1. Formation ofcircularXDNA.Proc.Natl. Acad.Sci. U.S.
58:240-247.
7. Green, D. M. 1964. Infectivity ofDNA isolated from Bacillus subtilis bacteriophage, SP82. J.Mol.Biol. 10:438-451.
8. Green, D. M. 1966. Intracellularinactivation of infective SP82 bacteriophage DNA. J. Mol. Biol.22:1-14.
9. Green, D. M. 1966. Physical and genetic charac-terization of sheared infective SP82 bacterio-phageDNA. J. Mol. Biol. 22:15-22.
10. Kammen, H. O., R. J. Wojnar, and E. S. Canellakis. 1966. Transformation in Bacillus subtilis. II.The development and maintenance
of thecompetent state.Biochim.Biophys. Acta 123:56-65.
11. Lerman, L. S., and L. J. Tolmach. 1957. Genetic transformation. I. Cellular incorporation of
DNA accompanying transformation in pneu-mococcus.Biochim.Biophys. Acta 26:68-82.
12. Levinthal, C., and N.Visconti. 1953. Growthand recombination in bacterial viruses. Genetics
38:500-511.
13. McCarthy, C., and E. W. Nester. 1967. Macro-molecularsynthesis innewly transformedcells
of Bacillus subtilis. J. Bacteriol. 94:131-140. 14. Mosig, G. 1962. The effect of multiplicity of
in-fection on recombination values in
bacterio-phage T4D. Z. Vererbungslehre 93:280-286.
15. Okubo, S., and W. R. Romig. 1965. Comparison of ultraviolet sensitivity of Bacillus subtilis bacteriophage SP02 and its infectious DNA.
J. Mol.Biol. 14:130-142.
16. Okubo, S., and W. R. Romig. 1966. Impaired transformability of Bacillus subtilis mutant
sensitivetomitomycin C and ultraviolet radia-tion. J. Mol. Biol. 15:440-454.
17. Okubo, S., B. Strauss, and M. Stodolsky. 1964. The possiblerole of recombination inthe infec-tion ofcompetentBacillussubtilis by bacterio-phage deoxyribonucleic acid. Virology
24:552-562.
18. Reilly, B. E., and J. Spizizen. 1965. Bacteriophage deoxyribonucleate infection of competent
Bacillus subtilis. J. Bacteriol. 89:782-790. 19. Wolstenholme, D. R., C. A. Verneulen, and G.
Venema. 1966. Evidence for theinvolvement of membranousbodies in theprocessesleadingto
genetic transformation in Bacillus subtilis. J.Bacteriol. 92:1111-1121.