Copyright 0 1996 by the Genetics Society of America
A Novel Tyl-Mediated Fragmentation Method for Native and Artificial
Yeast Chromosomes Reveals
That
the Mouse Steel Gene
is a Hotspot for Tyl Integration
Jacob Z. Dalgaard,'
Mukti
Banerjee and M. Joan Curcio
Molecular Genetics Program, Wadsworth Center and S.U.N. Y. School of Public Health, David Axelrod Institute, Albany, New York 12201-2002
Manuscript received November 27, 1995 Accepted for publication February 22, 1996
ABSTRACT
We have developed a powerful new tool for the physical analysis of genomes called Tyl-mediated chromosomal fragmentation and have used the method to map 24 retrotransposon insertions into two
different mousederived yeast artificial chromosomes (YACs). Expression of a plasmid-encoded GAL1:Tyl fusion element marked with the retrotransposition indicator gene, ade2AZ, resulted in a high fraction of cells that sustained a single Tyl insertion marked with ADE2. Strains in which TylADE2 inserted into aYAC were identified by cosegregation of the ADE2 gene with the URA3marked YAC. TylADE.2 elements also carried a site for the endonuclease I-DmoI, which we demonstrate is not present anywhere in the yeast genome. Consequently, I-DmoI cleaved a single chromosome or YAC at the unique site of TylADE.2 insertion, allowing rapid mapping of integration events. Our analyses showed that the frequency of TylADEZ integration into YACs is equivalent to or higher than that expected based on random insertion. Remarkably, the 50-kb transcription unit of the mouse Steel locus was shown to be a highly significant hotspot for Tyl integration. The accessibility of mammalian transcription units to Tyl insertion stands in contrast to that of yeast transcription units.
R
ETROTRANSPOSONS are mobile DNA sequences that resemble retroviral proviruses. Duplicative transposition occurs by integration of a cDNA copy of the retrotransposon into a new site in the same ge- nome. The Tyl element of Saccharomyces cerevisiae is the most abundant of the five classes of retrotransposons in yeast, with -30-35 copies in a typical haploid labo- ratory strain. Tyl is also the most common insertional mutagen in yeast, in part because of its relaxed target specificity (reviewed in SANDMEYER et al. 1990; BOEKEand SANDMEYER 1991). Only a very degenerate consen- sus sequence for Tyl integration has been found (NAT-
SOULIS et al. 1989; JI et al. 1993). However, Tyl integra- tion is nonrandom, which probably reflects influences at several levels including DNA conformation, associa- tion of DNA-binding proteins, transcriptional activity and the condensation state of the chromatin (CRAIGIE 1992). Tyl elements are preferentially targeted to A/ T-rich intergenic regions and are frequently found within several hundred base pairs of RNA polymerase 111-transcribed tRNA genes (EIGEL and FELDMANN 1982; OYEN and GABRIELSEN 1983; JI et al. 1993). In contrast, coding regions are underrepresented as inte- gration targets (GARFINKEL et al. 1988; JI et al. 1993).
Currespading author; M. Joan Curcio, Molecular Genetics Program, Wadsworth Center and S.U.N.Y. School of Public Health, David Axel- rod Institute, P.O. Box 22002, Albany, NY 12201-2002.
E-mail: [email protected]
ter, Frederick, MD 21702-1201.
'
Present address: Frederick Cancer Research and Development Cen-Genetics 143 673-683 (June, 1996)
However, Tyl insertions into many RNA polymerase 11-transcribed genes have been isolated on the basis of their causing a selectable phenotype by activation or inactivation of the target gene (SCHERER et al. 1982; EIBEL and PHILIPPSEN 1984; PAQUIN and WILLIAMSON
674 J. Z. Dalgaard, M. Banerjee and M. J. Curcio
either native or artificial yeast chromosomes. The strat- egy, named Ty I-mediated chromosomal fragmentation (TMCF), uses a pGTyl element marked with ade2AI, a retrotransposition indicator gene based on the yeast ADE2 gene. To simplify the physical mapping of Tyl- ADEZ retrotransposition events, the recognition site for a site-specific endonuclease, I-Dm01 (DALGAARD et al. 1994), was also cloned into the Tylade2AIelement. The I-DmoI endonuclease is encoded by an archaeal intron from the hyperthermophile, Desulfurococcus mobilis
(DALGAARD et al. 1993). It belongs to a family of site- specific endonucleases that are characterized by the presence of a repeated amino acid motif, LAGLI- DADG, and by their recognition of long nonpalin- dromic cleavage sites (MUELLER et al. 1993). Several of these endonucleases have been shown to be very rare or noncutters in yeast and other large genomes and have proved useful for mapping purposes (THIERRY and DUJON 1992; LIU et al. 1993). The I-DmoI recognition site has been estimated to span 20 bp, with some varia- tion in the sequence being tolerated (DALGAARD et al. 1994). We demonstrate here that I-DmoI does not cleave the yeast genome, and that a chromosome that sustains an ADE2-marked Tyl insertion is fragmented in vitro at a unique location by I-DmoI.
In this report, we used TMCF to generate a collection of Tyl insertions into mouse-derived artificial chromo- somes. Unexpectedly, induction of the ade2AJmarked Tyl element resulted in a unique TylADE2 insertion in each cell. This allowed a simple visual assay for cosegre- gation of the ADE2 marker with the YAC URA3 marker to be used to identify strains with Tyl insertions onto YACs. Physical mapping of the insertions was then per- formed using I-Dm01 fragmentation. We have shown that YACs are as accessible to Tyl transposition as yeast chromosomes and have demonstrated that a 50-kb tran- scription unit on a mouse-derived YAC is a hotspot for Tyl insertion.
MATERIALS AND METHODS
Plasmid constructions: pGTylade2tV: A PCR fragment con- taining the ADE2gene (nucleotides 445-2366) was synthesized with the oligomers 7830 (GCCGCCGCCGCCATCGATAGATG TCCGCGGCTGACAAATGACTCTTGTTGCAGTG) and 7831
CTAGATAA GCTTCG) from plasmid pASZl0 (STOTZ and LINDER 1990). The fragment was digested with CluI and cloned into the ClaI site of pGTyl-H3CLA (GARFINKEL et al. 1988). A 104bp SnaBI-PUuII fragment containing an AI (YOSHIMATSU
and NAGAWA 1989) was cloned by blunt-end ligation into the StuI site of the ADE2 gene.
pGTylade2AI-Dl: The 28-bp oligomers dmol (GCGCCGG AACTTACCCGGCAAGGCATGC) and dm02 (ATGCCTTGC- CGGGTAAGTTCCGGCGCGC) were annealed and inserted into the unique Sac11 site at the 5' end of the ade2AZgene (within oligomer 7830 sequence) in pGTylade2AZ. The orien- tation of the latter oligomer is relative to the ADE2 gene coding strand.
pCTyladeZAAI-DP: A 1.7-kb BamHI-StuI fragment of pGTyl- (GCGCGCCCGCCCATCGATCGATCTATATTACTTGTTTT-
TABLE 1
Yeast strains
Strain Genotype YAG
JC418 MATa, ura?-167, his? A200, adeZA::hisG, trp1::hisG
AB1380 MATa ade2-1 canl-100 lys2-I trpl
ura?-52 his5 thr- y +
YPH925 MATa, leu2Al trplA63 ura?-52 ade2-101 his?A200 lys2-801 cyh2 karlA15
JC917 YPH925 YLE394
JC919 YPH925 G5H340
ade2AJDl containing the 5' half of the URA3gene was deleted and replaced by a 0.9-kb BamHI-EcoRV fragment from pGEM- HIS3 (CURCIO and GARFINKEL 1992) containing the yeast HZS3 gene.
pSPADE2: The 2.3-kb BgLII fragment containing the ADE2 gene was subcloned from pASZl0 (STOTZ and LINDER 1990) into the BgLII site of pSP70 (Promega).
Strains and media: The genotypes of the strains used in this study are listed in Table 1. Strain JC418 is an isogenic derivative of strain GRF167 (BOEKE et al. 1985). The trp1::hisG and ade2A::hisG alleles were introduced by the gene disrup- tion method of ALAN1 et al. (1987) using plasmids pNKY1009 [a gift from N. KLECKNER and described in A I A N I et al. (1987)] and pAADE2 [a gift from D. GOTTSCHLINC and described in ApARlc10 et al. (1991)l.
Strains JC917 and JC919 were made by chromoduction of
mousederived YACs YLE394 (LARIN et al. 1991) and G5H340 (also named FAK-G3; KUSUMI et al. 1993), respectively, into recipient strain YPH925 (SPENCER et al. 1994). Patches of strain WH925 and the donor strain AB1380 containing either YLE394 or G5H340 were replicated onto a single YPD plate
and mated -14 hr at 30". Mating mixtures were then repli- cated to SC -ura-trp+cyh and incubated for 4 days at 30" to select cells in which the URA3 and TWl-marked YAC had been chromoduced into the cycloheximide-resistant recipi- ent, YPH925. Papillae were repurified on SC-ura-trp+cyh, patched onto SC -ura-trp plates and then replicated to the appropriate media to verify that they were Ura', Trp', His-, Lys-, Can', and Leu- haploid strains.
Rich medium (YF'D) was prepared as described (ROSE et al. 1990). All synthetic complete media used for this study were obtained from BIO101, Inc. and contained limiting amounts of adenine (10 mg/L) to allow red color development in Ade- strains. Glucose (2%) was used as a carbon source except in galactose medium, which contained 2% galactose.
Transposition assays: Selection for Ade+ prototrophs that sustained TylADE2 transposition events was performed in strain JC418 transformed with plasmid pGTylade2AZ by the lithium acetate method (ITO et al. 1983). Single colonies grown at 30" on SC "ura medium were used to inoculate 5 ml cultures of SC -ura galactose. Cultures were grown for 2 days at 20". Subsequently, a 200-pl aliquot of each culture was plated on SC -ade medium and grown at 30". The cells were titered by plating onto WD. Independent Ade+ prototrophs obtained from separate galactose-induction cultures were re- struck on YPD plates and replicated to SC -ura plates to identify Ura- plasmid segregants. Genomic DNA was pre- pared, digested with Asp718 and hybridized to a "P-labeled ADE2 RNA probe.
Ty-Mediated Chromosome Fragmentation 675
ber of unspliced (Tylade2AI) and spliced (TylADE2) transpo- sition events resulting from induction of plasmid pGTyl-
ade2AZ, essentially as described previously (CURCIO and GARFINKEL 1991). Strain JC418 containing plasmid pGTyl- ade2AZwas pregrown on SC -ura plates at 30" and then re- struck onto SC -ura galactose plates and grown for 6 days at 20". Independent colonies were struck onto SC "ura plates and grown at 30" to terminate induction and select cells that maintained plasmid pGTylade2AZ. Ura- segregants were ob-
tained following nonselective growth on WD and then scored for Ade+ prototrophy by replicating onto SC -ade medium. Genomic DNA was prepared from each strain and digested with Asp718, as well as Asp718 plus StuI. Following electropho- resis on agarose gels, DNA was hybridized to a "P-labeled ADE2 RNA probe to determine the number of TylADE2 and Tylade2AZ insertions per cell.
Identification of strains containing TylADE2D insertions on YAG: Strains JC917 and JC919 were transformed with pGTylade2AZ-D2 by the lithium acetate method. Trans- formants were selected, purified and maintained on SC -ur- a-trp-his plates, which selects for both the YAC and the HZS3based pGTylade2AZ-D2 plasmid. Transposition was in- duced by inoculating 5-ml cultures of SC -his-ura-trp galac- tose medium and incubating for -24 hr at 20". A 400-pl aliquot of each culture was plated on SC -ade-ura-trp glu- cose plates and incubated at 30" for 5 days. White colonies were patched onto YPD and incubated for 2 days at 30". Patches were replicated to both SC -ura-trp-ade medium and to medium containing 5-fluoroorotic acid (SFOA, BOEKE et al. 1987) and grown at 30" for a 5-7 days. Strains that developed a pink color on 5-FOA plates but remained white on SC "ura-trp-ade medium were identified. Pink patches of cells were shown to contain a mixture of white Urn- Trp' Ade' and red Ura- Trp- Ade- colonies, indicating that the 5-FOA-resistant phenotype can arise by telomeric positional silencing or mutation of URA3 or as well as by segregation of the YAC. For strains identified by pink growth on 5-F0A, cells were taken from WD medium and struck for single colonies on YPD. The colonies were replicated to SC -ade, SC --ura, SC -trp and SC -his media to demonstrate that Ura- Trp- colonies that had segregated the YAC during nonselective growth were Ade-, indicating cosegregation of the TylADE2- D with the YAC. A single Ade+ Ura+ Trp+ His- colony was isolated from each of these strains and analyzed by I-Dm01 fragmentation.
I-Dm01 fragmentation of agarose-embedded chromosomal
DNA Chromosomal DNA embedded in agarose was pre- pared from 1 ml of an overnight culture in WD medium by the method of GERRINC et al. (1991). For digestion with I- DmoI, DNA in agarose plugs was incubated in 2 ml of 50 mM EDTA, 10 mM Tris-HC1 pH 8 containing 1 mM phenylmethyl- sulfanyl fluoride for 1 hr at 4" and subsequently washed twice in 2 ml of 50 mM EDTA, 10 mM Tris-HC1 for 1 hr at 4". After equilibrating twice with I-Dm01 buffer (50 mM Tris-acetate pH 8.0, 25 mM ammonium acetate, 10 mM magnesium acetate, 1 mg/ml bovine serum albumin) for 30 min at 50°, chromo- some plugs were incubated with 10 units of purified I-Dm01 (DALGAARD et al. 1994) in I-Dm01 buffer for 3 hr at 50". Subse- quently, agarose-plugs were washed in 50 mM EDTA, 10 mM Tris-HC1 and stored at 4".
Pulsed-field gel electrophoresis (PFGE): Using a Biorad CHEF Mapper apparatus, samples were electrophoresed on 1% agarose gels (SeaKem Fastlane) in 0.5X TBE buffer at 6 V/cm for 24 hr. Pulse times of 60-120 sec were used to analyze native yeast chromosomes (Figure 2); 40-75 sec pulse times were used for YAC fragmentation gels (Figures 3 and 4). The gels were stained with ethidium bromide and photo- graphed.
Hybridizations: DNA blots were prepared from PFGE and conventional agarose gels by transfer of DNA to Hybond N+ membrane (Amersham). The "P-labeled ADE2 RNA probe was synthesized in vitro from plasmid pSPADE2 using Ribo- probe Gemini System 11 (Promega). "P-labeled DNA probes were prepared by random priming of a DNA fragment using High Prime DNA Labeling Mix (Boehringer Mannheim) . The 0.4kb StuI-SmaI fragment of URA3 was used as a URA3 probe. The 5.5-kb full-length Steel cDNA, 1.0-kb Steel open reading frame (OW) cDNA and 4.0-kb Steel 3' untranslated region (UTR) cDNA were provided by M. BEDELL and are described in BEDELL et al. (1995a).
RESULTS
Single Tyl insertion events generated using a new retrotransposition indicator gene: We constructed a new retrotransposition indicator gene based on the yeast D E 2 gene. One advantage of ADE2 as an indicator gene is that ADE2 prototrophs form white colonies that are easily distinguished from ade2 auxotrophic colonies, which are red (ROMAN 1956). Consequently, both gain and loss of a Tyl element marked with ADE2 can be visualized easily. To convert ADE2 into a retrotransposi- tion indicator gene, an artificial intron lacking flanking exon sequences (YOSHIMATSU and NAGAWA 1989) was
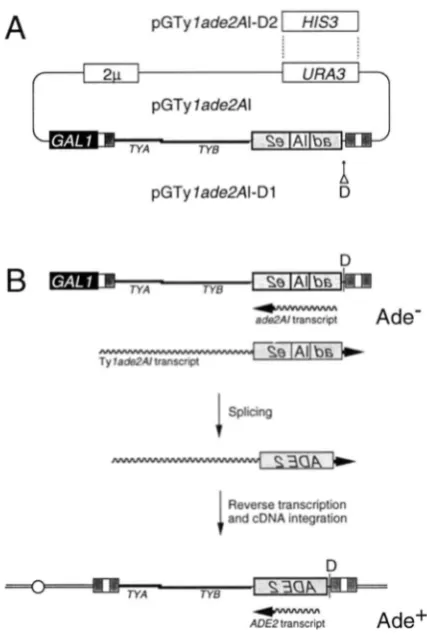
cloned into a unique StuI site in the ADE2 coding se- quence in an antisense orientation, thereby inactivating the gene. The disrupted ADE2 gene (ade2Al) was then cloned into the 3' untranslated region of the GAL1:Tyl- H3 fusion element on plasmid pGTyl-H3 (Figure l ) ,
such that the transcriptional orientations of Ty I-H3 and ade2Mwere opposed. This placed the AI in the correct orientation to be spliced from the Tylade2AItranscript. Use of the spliced transcript as a template for cDNA synthesis and genomic integration results in the forma- tion of a Tyl element containing a functional ADE2 gene and gives rise to an Ade' phenotype.
Transposition of the pGTylade2AI element was char- acterized in strain JC418 by growth of transformants on galactose for 2 days. Ade' prototrophs were generated at a frequency of 3.0%. In comparison, <2.2 X
Ade+ prototrophs were generated in control strains containing pGTyl-H3 marked with an ade2AI gene in the same transcriptional orientation as Tyl-H3, which renders the intron unspliceable from both transcripts. These results demonstrate that pGTylade2AIinduction gives rise to RNA-mediated transposition events at high efficiency. Southern blot analysis of DNA from 29 inde- pendent Ade+ prototrophs using an D E 2 probe re- vealed that 26 (92%) contained a unique TylADE2 in- sertion, while the other three strains had two insertions (data not shown).
This high proportion of colonies that had only one TylADE2 transposition event contrasts with earlier ob- servations for the his3AI indicator gene in pGTyI-H3
676
1.
Z. Dalgaard, M. Banejee and M . J . CurcioA
pGTylade2ALD21
-I
URA3pGTy lade2Al
h
pGTylade2Al-Dl D
Ade'
Tylade2Allranscript 29 IAllbs
1
SplicingJ
and cDNA integrationReverse transcription
D
A
P I A
FIGURE 1.-Plasmid and selection scheme. (A) Structure of transposition-induction plasmids used in the study. Plasmid pCTylnd~2AI contains the Tyl-H3 element, consisting of two long terminal repeats (LTRs; represented by tripartite rectan- gles) bracketing a region containing two partially overlapping ORFs (TYA anti TYlj). The upstream LTR was f k e d to the GAL1 promoter, represented by a black rectangle. A 104-bp artificial intron (AI) was inserted into the yeast ADE2 coding sequences in antisense orientation. This nde2AI gene was in- serted between TYB and the 3' LTR in the opposite transcrip tional orientation of the TyI-HS transcript, which places the AI in the sense orientation relative to transcription of Tyl. The yeast 2p origin of replication ( 2 p ) and the yeast URA3 gene (UXA3) are indicated. Plasmid pCTylnd~2AI-Dl has an I-Dm01 site (D) inserted between the nd~2AI gene and the Tyl 3' LTR. Plasmid pGTyld~2AI-D2 was constructed from pCTyInd~2Al-Dl by replacing the URA3 marker with the HIS3 gene. (D) Selection for Ade' retrotransposition events. Two transcripts in opposite orientations ( w a y lines) are generated from the (;AI,I:Tylnd~2A1fl1sion element: the galactose-induc- ible Tylnd~2AI transcript and the constitutive nde2AI tran- script. The AI is unspliceable from the latter since it is present in antisense orientation in the ALE2 gene. Since neither RNA encodes a functional ADE2 gene product, cells containing pGTylnde2Al plasmids are phenotypically Ade-. When the Tvlnd~2Altranscript is correctly spliced and used as a template for cDNA synthesis and integration, a TyIADE2 element is created. Cells that sustain TyIAZX2 transposition carry a con- stitutively expressed ADE2 gene and are phenotypically Ade'.
duced for expression of pGTylarlP2Al (Table 2). Follow- ing galactose-induction for 6 days, 36 independent colo- nies that retained the pGTylad~2Alplasmid throughout induction were selected on medium lacking uracil. Five of the 36 colonies were Ade' prototrophs. Genomic
TABLE 2
Comparison of Tyl retrotransposition using different indicator genes
Total Spliced Unspliced marked elements elements elements Plasmid per genome" per genome" per genome
pGTylnd~2Al 0.1 ( 5 / 3 6 ) 0.1 (3/36) 0.2 (8/36) pGTylhis3Al' 0.2 (.5/24) 1.5 (35/24) 1.7 (40/24)
"TyIADE2 or Ty1HI.73 insertions. "Tylnd~2AI or Tylhi,r3Al insertions.
' CL'R(:IO and GARFINKEI. (1991).
DNA was prepared from 36 plasmid segregants and ana- lyzed by Southern blotting with an ADE2 probe. The presence of a StuI site in the ADE2 gene that is restored by precise splicing of the AI was used to differentiate spliced and unspliced Tyl insertions. Eight of the strains were found to contain a single marked element. These included the five Ade' prototrophs, each of which had an ADE2"marked Tyl element with a StuI site recreated at the splice junction. The other three strains were Ade- auxotrophs that harbored an un- spliced Tylnd~ZAl element. The total transposition fre- quency was 0.2 marked elements per galactose-induced cell, which is 8.5-fold lower than the transposition fre- quency observed with pCTylhis3AI (CURCIO and GAR-
FINKEL 1991). Moreover, the fraction of insertions with spliced indicator genes (62.5%) is fivefold higher fol- lowing pCTylad~2AZ induction than for pGTylhis3AI induction (Table 2). The lower transposition frequency and elevated splicing efficiency of the pGTylade2AZele- ment explain the very low background of unspliced TylarlP2AI insertions in Ade' prototrophs that sustain TylADE2 transposition events. This feature of pGTyl- ad~2AI transposition dramatically simplifies mapping of marked Tyl insertions compared to the his3AI system, in which multiple insertion events complicate the physi- cal and genetic analysis of transposition events.
Fragmentation of yeast chromosomes at TyImE2
integration sites: To allow rapid mapping of
ADEZ-
Tv-Mediated Chromosome Fragmentation 677
A
1 2 3 4 5 6 7 8 9 1 0
I - D m O l -
+
-
+
- +-
+ - +(680 kb) XI
-
(400 kb) XI'
-
(280 kb) XI"
-
w,
VI1-
x111,xv1-
I1,XIV- x
-
XI-
v,VIl/-
IX-
111-
VI - 1FIGURE 2.-1-Ihd fraglllrntario~~ ol'!cxt rhromosonrcs that sustain TyIAnE2and TyIADL2-D transposition. (A) PFGE analysis of chromosomes from intlepentlcnt Adc' prototrophs obtained following galactose-induction of plasmid pGTylnd~2Alor pGTyI- ndt72AI-Dl. Odd-numbered lanes contain undigested chromosomes, and even-numbered lanes contain chromosomal DNA di- gested with I-IhoI. Lanes 1 and 2: an Ade' strain carrying a TyIAIIE2 element (lacking an I-Dm01 site); lanes 9-10: four independent Ade' strains with TylAIII.2-D elements. The chromosome that sustained a n AIIE2-marked transposition event and fragment3 created by I-DmoI digestion are indicated by white arrows. In lanes 4 and 8, the smaller chromosome fragments of "80 and 120 kh, respectively, were electrophoresed off the gel. The position and approximate size of I-Dm01 fragments of chromosome X I , which sustained a TyIAIIE2-D insertion in the strain analyzed in lanes 5 and 6, are indicated on the left. The positions of yeast chromosomes are indicated on the right. (B) The DNA blot was hybridized to '"P-labeled ADE2 RNA.
of 0.05%. Chromosomal DNA was prepared from 18 independent Ade' Ura- strains, digested with I-DmoI, and separated by PFGE. Four examples are shown in Figure 2A, lanes 3-10. Each genome was cleaved by I-DmoI at a single position, resulting in the disappear- ance of a single chromosome band and the appearance of two smaller-sized bands. For each DNA sample, the combined size of the two I-DmoI fragments was equal
to that of the cleaved chromosome. For example, I-DmoI digestion of the genomic DNA sample in lane 6, Figure 2A resulted in loss of the 680-kb chromosome
XI and the appearance of two new bands, -400 and 280 kb in size, the latter of which comigrates with chro- mosome ll1. Southern analysis (Figure 2B) using an ADE2 probe verified that in each strain the chromo- some containing the TylADE2-D element is fragmented by I-DmoI digestion, and that a specific fragment con- taining the ADE2 marker gene is generated. In all 18 Ade' strains analyzed, a single TyIADE2-D integration event introduced by transposition-induction was shown
to be a unique site of I-Dmol cleavage.
Selection for TylADE2 transpositions into YACs: The features of the pGTylad~2Al transposition system allowed a simple scheme to be used to identify TyI- ADE2-D insertions into YACs. Since virtually all Ade' strains isolated after a brief transposition-induction sus- tained one TyIADE2-D insertion, strains in which Tyl- ADE2-D integrated into a URA3, TXPI-marked YAC should show cosegregation of the Ade' phenotype with the Ura' and Trp' phenotypes. Cosegregants can be
identified visually on medium that selects for Ura- cells that have segregated the YAC, since they will become Ade- and develop a red color.
To test the validity of this identification scheme, trans- position was induced in two yeast strains containing mouse chromosomal DNA-derived YACs, YLE394 or G5H340. YAC YLE394 has recently been shown to con- tain the murine Steel locus, which encodes mast cell growth factor (BEDEL.L et nl. 1995b). G5H340 contains the 3' end of the mouse reekgene, a large locus whose product, reelin, is involved in brain development
(BAR
et al. 1995). For this analysis, the YACs were transferred from the standard YAC library strain AF51380 to strain YPH925 by chromoduction. This allowed us to make use of the ade2-101 and his3A200 mutations in strain YPH925 for selection of Tyl transposition and plasmid
maintenance, respectively.
Plasmid pGTyIn&2AI-D2, a derivative of pGTy1- nd~2AI-Dl in which the plasmid URA3selectable marker
was replaced by the HIS3 gene (Figure lA), was intrc- duced intoYAGcontaining strains, JC917 (YLE394) and JC919 (G5H340). Expression of plasmid pCTylnde2AI- D2 was then induced in galactosecontaining medium for 24 hr. Following transposition-induction, Ura' Trp' Ade' prototrophs were selected. These colonies had maintained the URA3, TW1-YAC throughout induc- tion, and sustained TyIUE2-D transposition events. Al- ternatively, an Ade' phenotype might result from gene conversion in this strain background, since the a&2-
678 J. Z. Dalgaard, M. Banerjee and M. J. Curcio
TABLE 3
Frequency of TyIADE2D insertion onto YACs
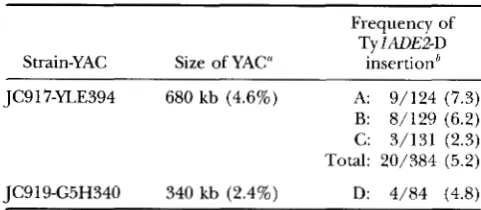
Frequency of Ty lADE2-D Strain-YAC of Size YAC" insertion'
JC917-YLE394 680 kb (4.6%) A 9/124 (7.3)
B: 8/129 (6.2) C: 3/131 (2.3) Total: 20/384 (5.2)
JC919-G5H340 340 kb (2.4%) D: 4/84 (4.8)
I' Physical size of the YAC in kilobases, and the percentage
of the total chromosomal DNA in the yeast strain represented by the YAC in parentheses.
'The number of Ade' Ura+ Trp' cosegregants identified after nonselective growth divided by the total number of Ade+ Ura' Trp+ colonies obtained following transposition-induc- tion. The calculated percentages are shown in parentheses.
with the ade2AI gene to create a functional ADE2 gene in the chromosome or on the plasmid. In fact, we no- ticed that -5% of the Ura' Trp+ Ade' colonies selected developed a pink color. The pink colonies were found to have an unstable Ade' phenotype that cosegregated with a His' phenotype, suggesting that they harbored plasmid-borne TylADE2 elements. These recombinants probably arose by gene conversion of the HZSmarked pGTylade2AI-D2 donor plasmid by either the ade2-101 allele or by spliced ADE2-containing cDNA. Pink Ade+ prototrophs were therefore eliminated from further analysis.
To identify Ura+ Trp' Ade+ strains in which a Tyl- ADE2-D element had inserted onto the YAC, Ura- se- gregants were selected on medium containing 5-FOA. A small percentage of the strains developed a pink color after several days growth on 5-FOA. When these strains were grown nonselectively, the ADE2 marker cosegre- gated with the URA3 and TRPl YAC markers, consistent with their having sustained TylADE2-D integration into the YAC. Twenty cosegregants were identified among 384 Ade' Ura' Trpf prototrophs isolated following ga- lactose-induction of pGTylade2AI-D2 in strain JC917 in three independent experiments (Table 3; A-C). Thus, the frequency of TylADE2-D transposition onto the 680- kb YAG, YLE394, was 5.2%, which is consistent with the 4.6% expected based on the physical size of the YAC relative to the total yeast genome. The 340-kb YAC, G5H340, in strain JC919 was also shown to be an effi- cient target for TylADE2-D insertion, as four cosegre- gants were identified among 84 white Ade+ Uraf Trp+ colonies obtained by transposition-induction (Table 3;
D). This frequency of 4.8% is twice that expected based on the percentage of the entire genome that G5H340 YAC DNA represents (2.4%). Moreover, these frequen- cies may be an underestimate of the fraction of de nouo transposition events on the YAG, since the total number of Adef Ura' Trpf prototrophs may include colonies
that sustain gene conversion of the adr2-101 allele. In summary, the data demonstrate that YAC DNA can be a target for Tyl insertion at a similar or higher fre- quency than that of yeast genomic DNA.
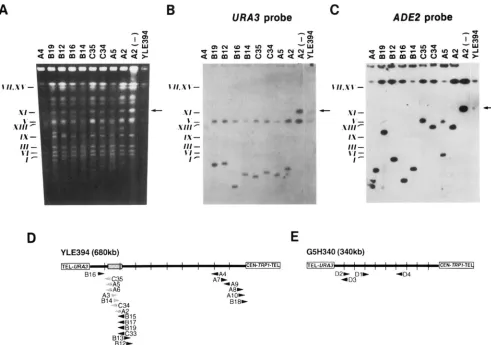
Mapping TylADE2 integration sites on YAGs by I-Dm01 fragmentation: Fragmentation with I-Dm01 was used to determine the location of the 20 TylADE2-D insertions identified phenotypically to be on the 680- kb YAC, YLE394. In PFGE analysis, YLE394 was not cleaved by I-Dm01 (Figure 3, A and B). However, YAC DNA in each strain containing a TyZADE2-D element was cleaved once by I-Dm& resulting in loss of the 680- kb YLE394 band (Figures 3A and 4B, indicated by
arrow) and the appearance of two smaller fragments. These results demonstrated that a single TyIAlK2-D element transposed onto the YAC in each strain. In
Southern analysis, one of the I-Dm01 fragments hybrid- ized to a URA3probe in each strain (Figure 3B and data not shown). The size of the URA3hybridizing fragment indicated the distance of the TylADE2-D element from the URA3marked terminus of the YAC. Hybridization to an ADE2 probe also detected one I-ll~aoI fragment in each strain (Figure 3C and data not shown). ( h n i - gration of the ADE2"hybridizing band with the URA3 hybridizing band was characteristic of TylADE2-D events oriented so that Tyl transcription proceeds to- ward the TRPl terminus. In strains with transposition events in the opposite orientation, ADE2 hybridized to the second YAC fragment generated by I-Dm01 diges- tion. The data were compiled to generate a map of the
20 TyIADE2-D insertions on YLE394 (Figure 3D). This analysis revealed two preferred regions of integration, each spanning -100 kb. Interestingly, almost all Tyl- ADE2-D insertions occurred into these domains, while the DNA segment of -350 kb that separated the two domains was refractoq to transposition. Six TylAlX2-
D elements were clustered in a 100-kb region near the TRPl-terminus. Another 13 transposition events were found in a region beginning
-
100 kb from the bEA3 terminus. Interestingly, the location of five of these Tyl- ADE2-D insertions in strains B13, B15, B17, B19 and C33 was indistinguishable within the resolution of PFGE, indicating that they were tightly clustered. The 20th TylADE2-D element, B16, mapped -50 kb from theURA3terminus.
In addition, we demonstrated that each of the four Ade' Ura' Trp' cosegregants obtained by transposi- tion-induction in strain JC919 also sustained a single TyIADE2-D transposition into YAC G5H340. A map o f the insertions, generated by I-Dm01 fragmentation and hybridization of PFGE blots to URA3 and A l X 2 probes, is shown in Figure 3E. Interestingly, three of the four TyIADE2-D elements integrated within -100 kb o f the URA3 terminus, a position that coincides with the re- gion of the YAC that contains the 3' end of the mouse
Ty-Mediated Chromosomc Fragmentation 679
A
URA3 probe
C
ADE2 probe
I-
IX- I
-
'7 7- " C C C II'll. .x I
-
.x1
-
. x l l l - t -
1.Y
-
111-
1 1 -
I C
D
E
G5H340 (340kb)
D 2 b Dl+ 4 0 4
4 0 3
FIGURE 3.-Mapping TylADE2-D insertions on YACs by I-Dm01 fragmentation. (A) PFGE analysis of chromosomes prepared from yeast strains identified in cosegregation analysis as having sustained TyIADE2-D insertion into YLE394 (see Table 3).
Chromosomes were digested with I-DmoI, except for strain A2 DNA, which was not digested. The arrow in each panel indicates the location of the unfragmented YLE394 YAG, which comigrated with chromosome X I . The positions of selected yeast chromo- somes are indicated on the left as a size reference. (B) The DNA blot was hybridized to a "P-labeled URA3 probe. (C) The blot was stripped and reprobed with a y2P-labeled ADE2 RNA probe. URA3 and ADE2 probes also hybridized to chromosome V (600 kb) and chromosome XV (1.1 Mb), respectively. Yeast Chromosome (Biorad) and Lambda Ladder (Gibco-BRL) markers were used to estimate the sizes of observed fragments (not shown). (D) Graphic map of TyIAIIE2-D insertions on YLE394. The shaded rectangle represents the 50-kb mouse S t e l gene transcription unit. Sections between vertical lines represent 1/10 of YLE394 DNA (68 kb). The position of each TyIADE2-D element is given by the back edge of an arrowhead. The orientation of the TyI element is indicated by the direction of each arrowhead. Shaded arrowheads represent TyIAIIE2-D elements that are located in the Sted transcription unit (see Figure 4 and text). (E) Graphic map of four TyIADE2-D insertions on YAC G3H340 (Table
3). Sections between vertical lines represent 1/10 of G5H340 DNA (34 kb).
The mouse Steel transcription unit is a hotspot for Tyl integration: The insertion of 13 of 20 (65%) Tyl- ADE2-D elements into a region representing only -15% of YLE394 DNA was particularly notable, since their location coincided with that of the only gene so far identified on YLE394, the mouse Sfwl gene (BEDEIL et nl. 1995b). To more precisely determine the position of the TylADE2-D element5 relative to the S f w l tran- scription unit, PFGE Southern blots were reexamined using a 5.5-kb full-length Sted cDNA probe (Figure 4A).
The probe hybridizes to an estimated 50 kb of mouse genomic sequences that include the 0.2-kb 5' UTR, a
42-kb region spanning the ORF, and an 8-kb 3' UTR (BEDELI. et nl. 1995a). The S~PPI cDNA probe detected both I-Dm01 fragment5 of the YAC in strains A5, R14,
C34, C35 (marked with closed arrowheads in Figure
4A), A3 and A6 (data not shown), indicating that these six strains contain TylADE2-D insertions in the Sfeel transcription unit. In contrast, only the TRPI-terminal fragment in strain B16, in which the TyIADE2-D inser- tion is located 50 kb closer to
URA3,
hybridized to the S t w l cDNA probe (Figure 4A). The position of the six TyIADE2-D elements in Sfeelwas further delimited usingJ. Z. Dalparcl, M. k m c j e e and M. J. Curcio
C
Steel ORF probe
A
Steel cDNA probe
L ,
"".
"m
"*
c
FIGLIRI. 4.-Mapping of TyZADE2-D insertions t o the S/wl transrl-iption ullit o n YLE394. (A) The PFGE b l o t sho\c.n i n Figure
3 was stripped antl hybridized to a 'v2P-labeled-9wl cDNA probe. Thc arrow on thc right of the blot denotes the position of the unfragmentedYAC. Closed arrowheads in samples from strains R14, C X , C34 and 115 indicate the two I-DmoI-generated fragment5 of YIX394. Rands marked with open arrowheads in strains A4, R14, C3.5, and A5 are I-DmoI-generated fragment5 of spontaneous deletions in YI,E394 (see text for explanation). (D) Chromosomal DNA from the strains indicated were digested with I-Dm01
and analyzed by PFGE. The position of unfragmented YLE394 is indicated bv the black arrow to thc left of the gel. (C) Sollthern analysis of R gel hybridized to a ""P-labeled 1 .O-kb S t w l ORF cDNA. Bands marked with closed arrowheads in samples C34, C35, A3 antl A5 are the two I-Dm01 frazmrnts of YI.E394. Open arrowheads denote bands resulting from spontaneous deletions in YAC YLE.794.
Southern hybridizations with the Steel cDNA and Stpel
ORF probes also detected a third I-Dm01 fragment in some strains (indicated by open arrowheads in Figure 4, A and C). The amount of this fragment varied between chromosomal preparations ( e . g , two different chromo- somal preparations of strain A5 in Figure 4C). This observation and the fact that the third fragment was proportionally smaller than the TRPI-terminal I-Dm01 fragment detected by both Steel probes strongly suggest that it result.. from a spontaneous deletion in YLE394. The deletion fragment was not detected by URA3 o r
Steel probes in I-Dm01 fragmented DNA from strains with TyIADK2-D insertions located just downstream of the
Steel ORF, such as B12 (Figures 3B and 4, A and C), indicating that deletion occurs downstream of Steel.
However, in I-DmoI-fragmented DNA from strains A4 and A3, A5, C34 and C35, the deletion fragment was detected by the Steel ORF probe (Figure 4C). The data demonstrate that deletion occurs in the middle of YLE394 between .'&x1 and the cluster of TyIADE2-D in- sertions near the TRPIend. Deletion does not result in
loss of the I-Dm01 site in any of the strains analyzed. Because a deletion fragment is detected in DNA from YLE394 prior TylADE2-D insertions (data not shown), this common deletion is probably unrelated to TyI- mE2-D integration.
Seven TylADE2-D insertions in strains A2, B12, B13, B15, B17, B19 and C33 mapped just downstream of
the Steel ORF (Figure 3D). In DNA samples from these strains, the full-length Steel cDNA probe hybridized strongly to the URA3terminal I-Dm01 fragment but also
weakly to the TRPI-terminal fragment, suggesting that they might be located in the 3' UTR (Figure 4A, note very light hybridization to a second band in B19 and B12). However, weak cross-hybridization between the
Steel cDNA probe and sequences at the TRPl-terminus was detected in strains with TylADE2-D elements
l o -
cated far away from Steel; therefore, we could not con- clusively determine if these seven TylADE2-D elements were in the Steel 3' UTR by I-Dm01 fragmentation. There- fore, Southern analysis of EcoRIdigested genomic DNA was performed using a 4.0-kb .$tee1 3' UTR cDNA probe. One TylADE2-D insertion in strain A2 disrupted a 2.7- kb EmRI fragment (data not shown). These data indi- cated that the TylADE2-D element in A2 is near the
Steel polyadenylation site contained within the 2.7-kb EcoRI fragment (BEDELI. et nl. 199%). The remaining six insertions mapped
-
10-25 kb downstream of theSteel polyadenylation site.
In summary, we found that seven of the total 384 TylADE2-D insertions in strain JC917 were distributed throughout the 50-kb SPPI transcription unit. This o h
servation is highly significant in chi-square analysis ( P 0.001). The data indicate that the Steel transcription unit is a hotspot for TyI integration relative to the en- tire strain genome. This is the first demonstration that
a mammalian gene on a YAC can be not only a target but a strongly preferred site of integration for the Tyl retrotransposon.
DISCUSSION
Ty-Mediated Chromosome Fragmentation 681
of Tyl insertions on endogenous or artificial yeast chromosomes. A galactose-inducible Tyl element has been marked with a retrotransposition indicator gene, ade2AI, that allows single-step phenotypic selection of colonies that sustain a transposition event. The indica- tor gene has a relatively high efficiency of splicing (62.5%), but Tyl elements marked with ade2AItrans- pose at a low frequency (Table 2). Because of these features, most of the Adef colonies isolated following a brief induction of pGTylade2AI expression contain a single Tyl insertion carrying the spliced ADE2 allele. In addition, we have marked the pGTylade2AI ele- ment with an I-DmoI cleavage site, which we demon- strated was not present anywhere in the yeast genome. Consequently, the position of a transposed TylADE2-
D element can be determined by an I-DmoI restriction digest, which fragments the yeast genome at a unique location on a single chromosome (Figure
2 ) .
The single ADE2-marked transposition events charac- teristic of TMCF also allow rapid identification of colo- nies in which integration onto YACs has occurred. A simple cosegregation assay that makes use of the color difference between Ade+ prototrophs and Ade- auxo- trophs was performed to demonstrate that two different YAGs were targeted for TylADE2-D insertion as or more efficiently as yeast genomic DNA (Table 3). Recently we have demonstrated that the frequency of TylADE2-
D transposition onto a third 700-kb mouse-derivedYAC was equivalent to that predicted by the size of the YAC relative to the yeast genome (unpublished results). These results show that despite significant differences in the organization of mammalian genomic DNA com- pared to yeast (e.g., lower gene density, presence of large introns, highly repetitive intergenic regions), mammalian genomic DNA can serve as an efficient tar- get for Tyl retrotransposition.
Although the frequency of TylADE2-D insertion onto the YACs was approximately that predicted by their physical size relative to the entire yeast genome, Tyl insertions were not randomly located on the YACs (Fig- ure 3, D and E). On the 680-kb YAG, YLE394, all but one of the 20 TylADE2-D insertions were found in two regions of -100 kb each, separated by a 350-kb region that sustained no Tyl insertions (Figure 3D). One of these preferred integration domains included five inser- tions so tightly clustered that their locations could not be distinguished on pulsed-field gels. This pattern of a very few highly used integration regions, including strongly preferred integration sites and large regions that are refractory to transposition, resembles that for another chromosome analyzed as a target for de novo Tyl integration specificity, yeast chromosome III ( JI et al. 1993). Similarly, the Rous sarcoma virus was shown to utilize only very few genomic hotspots in vivo (SHIH et al. 1988). The association of host proteins or some other specific in vivo property is thought to be involved in the configuration of Tyl integration hotspots, be-
cause in vivo hotspots are not preferred integration tar- gets for
Tyl
transposition in vitro ( JI et al. 1993). The appearance of a strong Tyl hotspot on extrinsic YAC DNA suggests that specific sequences on the YAC DNA are recognized in yeast, causing the formation of dis- tinct chromatin domains that are distinguished by the Tyl integration complex.The nonrandom pattern of Tyl integration on the two YACs analyzed in this study suggests the interesting possibility that Tyl is efficiently targeted to these YACs
(Table 3) because mammalian transcription units are preferred regions for Tyl integration. This hypothesis is strongly supported by the finding that the 50-kb Steel transcription unit onYLE394 is a highly significant hots- pot relative to the entire yeast genome. It is not yet known if there is a second transcription unit on YLE394 coinciding with the second common site of Tyl integra- tion (M. BEDELL, personal communication). However, we have recently learned that G5H340 encodes -30-
40 exons of the mouse reeler gene, beginning at the URA?-terminus, as well as two additional transcriptional units (or portions thereof) further downstream toward the TRPl-terminus (A. GOFFINET, personal communica- tion; BAR et al. 1995). Three of the four TylADE2-D elements mapped here (Figure 3E) are in a region that coincides with the location of the reeler gene. We are currently investigating whether the reeler locus on G5H340 is also a hotspot for Tyl transposition.
The finding that the Steel transcription unit was a preferred region for Tyl integration was unexpected, especially since yeast coding regions have been shown to be relatively poor targets for Tyl transposition. For example, ORFs on chromosome 114 including unex- pressed genes such as those found in the silent mating cassettes, were used as Tyl targets at a much lower fre- quency than expected if transposition were random ( JI
682 J. Z. Dalgaard, M. Banerjee and M. J. Curcio
integration. If so, TMCF could be very useful as a simple method of locating transcription units on YACs.
The introduction of a specific double-strand break in a chromosome dramatically simplifies large-scale physical mapping, cloning and sequencing of genomic DNA. Several strategies have previously been developed for generating single or very few specific cleavages in large genomes. One technique uses site-specific cleav- age in vitro through Cre-lox recombination (QIN et al.
1995). The Achilles' heel and nested chromosomal frag- mentation methods utilize homologous alignment or recombination to generate a unique site that can be cleaved by a restriction enzyme in vitro (KOOB and SZY- BALSKI 1990; THIERRY et al. 1991; COLLEAUX et al. 1993). A related strategy involves homology-mediated inser- tion of a linear telomere sequence for in vivo chromo- somal fragmentation (PAVAN et al. 1990). However, frag- mentation can only be achieved at predetermined sites using these methods, since alignment of homologous sequences is required. In contrast, a large number of independent Tyl integration events into targets that are not preselected can be obtained using TMCF. This feature may allowYAC DNA derived from any organism, even those lacking dispersed repetitive elements com- monly used for homologous targeting, to be analyzed by TMCF. And unlike the method commonly used for YAC fragmentation, telomere-sequence insertion at in- ternal sites (PAVAN et al. 1990), both ends of the frag- mented YAC are retained and available for analysis us- ing TMCF.
TMCF can expedite the characterization and manip ulation of YAC DNA in several ways. Restriction maps of DNA flanking the I-Dm01 site can be constructed using total genomic DNA by partial restriction digestion of I-DmoI-fragmented DNA and hybridization to an ADE2 or chromosomal-end probe. A set of strains with Tyl insertions at different locations on the YAC would allow overlapping restriction maps to be developed. Furthermore, overlapping chromosomal fragments generated by I-Dm01 fragmentation could be applied to the sorting of cosmid or lambda libraries (THIERRY and
DUJON 1992). Another application of TMCF is to render internal YAC DNA accessible for subcloning, anchored PCR or DNA sequencing. Because a unique fragment is generated by digestion with I-DmoI and a frequently cutting endonuclease, sequences flanking TylADE2-D insertions can be cloned directly into PUD718, a vector containing a polylinker and an I-Dm01 site (DALGAARD
et al. 1993). This strategy obviates the need for purifica- tion of YAC DNA from pulsed-field gels, which is fre- quently problematic. A third application of the system is to substitute I-Dm01 fragments for entire YACs in exon trapping and direct cDNA selection schemes to simplify these analyses. Finally, YACs containing TylADE2-D in- sertion mutations within putative genes could be used to construct transgenics. Moreover, strains with Tyl-
ADE2-D insertion mutations in genes could be used to
facilitate the secondary introduction of subtle nonse- lectable mutations by using loss of the ADE2 marker as a visual screen for introduction of the new mutation.
Galactose-inducible Tyl elements marked with HZS? or his?AIhave proven useful as insertional mutagens in yeast (GARFINKEL et al. 1988; KAWAKAMI et al. 1992). One drawback of their use, however, is that several marked Tyl transposition events are typically found in transposi- tion-induced colonies. These transposition events can include ones marked with an unspliced his3AI allele, which cannot be followed phenotypically (STRATHERN et al. 1994). The high frequency of single, spliced Tyl- ADE2-D insertions and the presence of a restriction site that is unique in the genome greatly simplify isolation, genetic analysis, and cloning of Tyl insertional muta- tions. In addition, strains containing the pGTylade2AI element could be very useful for screening for cellular genes that are required for Tyl transposition. Since galactose-induced colonies have a high degree of sec- toring from red to white because of TylADE2 transposi- tion events (M. J. CURCIO, unpublished observation), transposition mutants could be identified by a lower frequency of sectoring.
As illustrated by this report, another advantage of TMCF is that it can be used to rapidly map Tyl inser- tions on a large genomic target. Consequently, the ques- tion of how Tyl targeting to chromatin domains is affected by different genetic backgrounds can be ad- dressed by TMCF. Using a relatively small target, the CAN1 locus of yeast, it has been demonstrated that rad6A mutants have a reduced target-site specificity (LIEBMAN and NEWNAM 1993). Identification of host mutants in which Tyl insertion into YACs is random would be useful for some applications of TMCF, such as the physical analysis of large genomic regions.
The authors gratefully acknowledge M. BEDELL for providing a characterized YAC and DNA probes and for thoughtful comments on the manuscript. We are grateful to J. MONTGOMERY for providing YAC strains, J. KOWALSKI for technical assistance, M. BELFORT, M. BRYK, A. KIAR, C. LICHTENSTEIN and R. MOFSE for helpful comments on the manuscript and the Molecular Genetics Core Facility for oligo- mer synthesis and DNA sequencing. The research of J.Z.D. was sup- ported by Danish Research Council Ph.D. grant 11-9348-1 and Na- tional Institutes of Health (NIH) grant G"44844 to M. Belfort. This work was supported in part by NIH grant GM-52072 to M.J.C.
LITERATURE CITED
ALANI, E., L. CAo and N. KLECKNER, 1987 A method for gene disrup
tion that allows repeated use of URA3 selection in the construc- tion of multiply disrupted yeast strains. Genetics 116: 541-545.
APARICIO, 0. M., B. L. BILLINGTON and D. E. GOTTSCHLING, 1991 Modifiers of position effect are shared between telomeres and silent mating-type loci in S. cereviszue. Cell 66: 1279-1287. BAR, I., C. LAMBERT DE ROLJVROIT, I. ROYAUX, D. B. KRIZMAN, C.
DERNONCOURT et ul., 1995 A YAC contig map containing the
reeler locus with preliminary characterization of candidate gene fragments. Genomics 26: 543-549.
Ty-Mediated Chromosome Fragmentation 683
and N. A. JENKINS, 199513 Deletion and interallelic complemen- tation analysis of Steel mutant mice. Genetics 1 4 2 935-944. BOEKE, J. D., and S. B. SANDMEYER, 1991 Yeast transposable ele-
ments, pp. 193-262 in The Molecular and Cellular Biology of the Yeast Saccharomyces, edited by J. R. BROACH, J. PRINCLE and E. JONES. Cold Spring Harbor Laboratory Press, Cold Spring Har-
bor, NY.
BOEKE, J. D., D. J. GARFINKEL, C. A. S r n r s and G. R. FINK, 1985 Ty elements transpose through an RNA intermediate. Cell 40: 491-500.
BOEKE, J. D., J. TRUEHEART, G. NATSOULIS and G. R. FINK, 1987 5- Fluoroorotic acid as a selective agent in yeast molecular genetics. Methods Enzymol. 154 164-175.
BOEKE, J. D., H. Xu and G. R. FINK, 1988 A general method for the chromosomal amplification of genes in yeast. Science 239: 280-282.
BURNS, N., B. GRIMWADE, P. B. ROSS-MACDONALD, E.-Y. CHOI, K. FIN-
BERG rt al., 1994 Large-scale analysis of gene expression, protein localization and gene disruption in Saccharomyces cerevisiae. Genes Dev. 8: 1087-1105.
COI.I.EAUX, L., C. ROUGEULLE, P. AVNER and B. DUJON, 1993 Rapid physical mapping of YAC inserts by random integration of I-Sce
I sites. Hum. Mol. Genet. 2: 265-271.
CRAIGIE, R. 1992 Hotspots and warm spots: integration specificity of retroelements. Trends Genet. 8 187-190.
CURCIO, M. J., and D. J. GARFINKEI., 1991 Single-step selection for Tyl element retrotransposition. Proc. Natl. Acad. Sci. USA 88: CURCIO, M. J.. and D. J. GARFINKEI., 1992 Posttranslational control of Tyl retrotransposition occurs at the level of protein pro- cessing. Mol. Cell. Biol. 12: 2813-2825.
DALGM, J. Z., R. A. GARRETT and M. BEI.FORT, 1993 A site-specific endonuclease encoded by a typical archaeal intron. Proc. Natl. Acad. Sci. USA 90: 5414-5417.
DALGAARD, J. Z., R. A. GARRETT and M. BELFORT, 1994 Purification and characterization of two forms of I-DmoI, a thermophilic site- specific endonuclease encoded by an archaeal intron. J. Biol. Chem. 269: 28885-28892.
EIBEI., H., and P. PHIIJPPSEN, 1984 Preferential integration of yeast tranSpOSdbk element Tyl into a promoter region. Nature 307: 386-388.
EIGEI., A,, and H. FEI.DMANN, 1982 Tyl and delta elements occur adjacent to several tRNA genes in yeast. EMBO J. 1: 1245-1250. GARFINKEI., D. J., M. F. MASTRANCELO, N. J. SANDERS, B. K. SWER
and J. N. STRATHERN, 1988 Transposon tagging using Ty ele- ments in yeast. Genetics 120: 95-108.
GERRING, S. L., C. CONNELLY and P. HIETER, 1991 Positional m a p ping of genes by chromosome blotting and chromosome frag- mentation, pp. 57-76 in Guide to Yeast Genetics and Molecular Biology, edited by C. GUTHRIE and G. R. FINK. Academic Press, San Diego.
ITO, H., Y. FUKUDA, K. MURATA and A. KIMURA, 1983 Transforma- tion of intact yeast cells treated with alkali cations. J. Bacteriol.
JI, H., D. P. M o ~ R F - , M. A. BIOMBERG, L. T. BRAITEM, D. F. VOWAS
rt al., 1993 Hotspots for unselected Tyl transposition events on yeast chromosome III are near tRNA genes and LTR sequences. Cell 73: 1007-1018.
KAWAKAMI, K., B. K. SWER, D. J. GARFINKEL, J. N. STRATHERN and
Y. N.WURA, 1992 Ty element-induced temperature-sensitive mutations of Saccharomyces cerevisiae. Genetics 131: 821-832. KOOB, M., and W. S ~ ~ ~ S K I , 1990 Cleaving yeast and Escherichia coli
genomes at a single site. Science 250 271-273.
KLWMI, K., J. S. SMITH, J. A. SEGRE, D. S. KOOS and E. S. LANDER, 1993 Construction of a large-insert yeast artificial chromosome library o f the mouse genome. Mamm. Genome 4 391 -392. 936-940.
153: 163-168.
LARIN, Z., A. P. MONACO and H. LEHRACH, 1991 Yeast artificial chro- mosome libraries containing large inserts from mouse and hu- man DNA. Proc. Natl. Acad. Sci. USA 88: 4123-4127. LIEBMAN, S. W., and G. NEWNAM, 1993 A ubiquitin-conjugating en-
zyme, RAD6, affects the distribution of Tyl retrotransposon inte- gration positions. Genetics 1 3 3 499-508.
LIU, S. L., A. HESSEI. and K. E. SANDERSON, 1993 Genomic mapping with I-CeuI, an intronencoded endonuclease specific for genes for ribosomal RNA, in Salmonella spp., Escherichia coli, and other bacteria. Proc. Natl. Acad. Sci. USA 90: 6874-6878.
MUELLER, J. E., M. BRYK, N. LOIZOS and M. BELFORT, 1993 Homing endonucleases, pp. 111-143 in Nucleases, edited by S. M. LINN, R. S. LLOYD and R. J. ROBERTS. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N Y .
NATSOULIS, G., W. THOMAS, M. C. ROGHMANN, F. WINSTON and J. D. BOEKE, 1989 Tyl transposition in Saccharomyces cerevisiae is non- random. Genetics 123: 269-279.
OYEN, T. B., and 0. S. GABRIELSEN, 1983 Non-random distribution
of the Tyl elements within nuclear DNA of Saccharomyces cereuis-
iae. FEBS Lett. 161: 201-206.
PAQUIN, C. E., and V. M. WILLIAMSON, 1986 Ty insertions at two
loci account for most of the spontaneous antimycin A resistance mutations during growth at 15°C of Saccharomyces cereuisiaestrains lacking ADHI. Mol. Cell. Biol. 6: 70-79.
PAVAN, W. J., P. HIETER and R. H. REEVES, 1990 Generation of dele- tion derivatives by targeted transformation of humanderived yeast artificial chromosomes. Proc. Natl. Acad. Sci. USA 87: 1300-1304.
QIN, M., E. LEE, T. ZANKIX and D. W. OW, 1995 Site-specific cleavage Acids Res. 23: 1923-1927.
of chromosomes in vitro through Cre-lox recombination. Nucleic ROMAN, H., 1956 Studies of gene mutation in Saccharomyces. Cold
Spring Harbor Symp. Quant. Biol. 21: 175-185.
ROSE, M. D., F. WINSTON and P. HIETER, 1990 Methods in Yeat Genet-
ics: A Laboratmy Course Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N Y .
SANDMEYER, S. B., L. J. HANSEN and D. L. CHALKER, 1990 Integration specificity of retrotransposons and retroviruses. Annu. Rev. Ge- netics 2 4 491-518.
SCHERER, S., C. MANN and R. W. DAVIS, 1982 Reversion of a pro- moter deletion in yeast. Nature 298: 815-819.
SHIH, C.-C., J. P. STOYE and J. M. COFFIN, 1988 Highly preferred targets for retrovirus insertion. Cell 53: 531-537.
SPENCER, F., Y. HUGERAT, G. SIMCHEN, 0. HURKO, C. CONNELLY et al., 1994 Yeast karl mutants provide an effective method for YAC transfer to new hosts. Genomics 22: 118-126.
STOTZ, A., and P. LINDER, 1990 The ADE2 gene from Saccharomyces cereuisiae: sequence and new vectors. Gene 95: 91-98.
STRATHERN, J. N., M. MASTRANGELO, L. A. RINCKEL and D. J. GARFIN- KEL, 1994 Ty insertional mutagenesis, pp. 1 1 1 - 120 in Molecular Genetics of Yeast: A Practical Approach, edited by J. R. JOHNSTON.
Oxford University Press, Oxford.
THIERRY, A., and B. DUJON, 1992 Nested chromosomal fragmenta- tion in yeast using the meganuclease I-SceI: a new method for physical mapping of eucaryotic genomes. Nucleic Acids Res. 20:
THIERRY, A., A. PERRIN, J. BOER, C. FAIRHEAD, B. DUJON et al., 1991 Cleavage of yeast and bacteriophage T7 genomes at a single site using the rare cutter endonuclease I-SceI. Nucleic Acids Res. 19: 189-190.
WILKE, C. M., S. H. HEIDLER, N. BROWN and S. W. LIEBMAN, 1989 Analysis of yeast retrotransposon Ty insertions at the CAN1 locus. Genetics 1 2 3 655-665.
YOSHIMATSU, T., and F. NAGAWA, 1989 Control of gene expression by artificial introns in Saccharomyces cerevisiae. Science 244: 1346-
1348. 5625-5631.