Multiple Elements
and
Auto-Repression Regulate Roxl,
a
Repressor
of Hypoxic
Genes
in
Saccharomyces cereuiSiae
Jutta Deckert, Roberto Perhi,' Bhuvana Balasubramanian and Richard S. Zitomer
Department of Biological Sciences, The State University of New York at Albany, Albany, New York 12222
Manuscript received May 31, 1994 Accepted for publication November 21, 1994
ABSTRACT
The ROXl gene encodes a heme-induced repressor of hypoxic genes in yeast. Using RNA blot analysis
and a ROXI/ lacZ fusion construct that included the ROXl upstream region and only the first codon, we discovered that Roxl represses its own expression. Gel-retardation experiments indicated that Roxl was capable of binding to its own upstream region. Overexpression of Roxl from the inducible GAL1
promoter was found to be inhibitory to cell growth. Also, we found that, as reported previously, Hapl is partially responsible for heme-induction of ROXl, but, in addition, it also may play a role in ROXl
repression in the absence of heme. There is a second repressor of anaerobic ROXI expression that requires the general repressor Tupl /Ssn6 for its function.
B
AKER'S yeast contains a set of hypoxic genes thatare repressed under fully aerobic conditions and induced when oxygen is limiting ( ZITOMER and LOWRY 1992). Many of the hypoxic genes encode enzymes that use molecular oxygen as a substrate including oxidases, desaturases and reductases in heme, sterol, and fatty acid biosyntheses. Presumably the levels of these enzymes are increased to compensate for decreasing oxygen levels under hypoxic conditions. The importance of main- taining these reactions is illustrated by the fact that yeast cells grown in the complete absence of oxygen cannot synthesize membrane components and the growth me- dium must be supplemented with sterols and fatty acids
(ANDREASEN and STIER 1953, 1954). Other hypoxic genes encode oxygen related functions such as an alter- nate subunit V of cytochrome c oxidase, an alternate cytochrome c and an alternate mitochondrial ATP/ADP translocator (ZITOMER and LOWRY 1992); it is as yet unclear whether these alternate proteins increase the cell's respiratory capacity under hypoxic conditions.
The hypoxic genes share a common oxygen-depen- dent repression mechanism. Cells sense oxygen avail- ability through the levels of heme, the synthesis of which requires molecular oxygen ( MATTOON et al. 1979; LABBE-BOIS and LABBE 1990). Heme activates the transcription of the ROXl gene, which encodes the hyp- oxic repressor that binds to a 12-bp hypoxic consensus sequence ( LOWRY et al. 1990; BALASUBRAMANIAN et al. 1993). This sequence has been found in the upstream region of all the hypoxic genes sequenced to date ( ZI-
TOMER and LOWRY 1992)
.
In addition to the hypoxic-Curresponding authvr: Richard S. Zitomer, Department of Biological
'
Present address: ENEA, Agrobiotechnology Department, C.P. 2400, Sciences, University at Albany/SUNY, Albany, NY 12222.Roma 00100 A D , Italy.
Genetics 139: 1149-1158 (March, 1995)
specific Roxl repressor, the general repressors Tupl and Ssn6 are required for repression ( ZHANG et al. 1991; BALASUBRAMANIAN et al. 1993). Thus, the general phe- nomonology of hypoxic gene regulation can be de- scribed as follows. When cells are grown under fully aerobic conditions, heme accumulates to levels suffi- cient to induce ROXl expression and the hypoxic genes are repressed. When cells are limited for oxygen, heme levels fall, Roxl repressor levels are reduced and hyp- oxic gene expression is derepressed. It should be noted that although the hypoxic genes are all repressed by Roxl, they do not share a common transcriptional acti- vation mechanism, so they do not comprise a regulon of coordinately expressed genes.
The above scheme leaves some important questions unanswered. First, what is responsible for the hemede- pendent expression of ROXl? A logical candidate is the hemedependent transcriptional activator Hapl ( GUARE- NTE and FOSBURG 1989), and KENG ( 1992) reported that Hapl does indeed induce ROXl expression. We have h r - ther investigated this effect with some surprising results. Second, how are the correct Roxl levels maintained in the cell? This question arises because some of the hypoxic gene products are required at low levels for aerobic growth, for example those in the heme, fatty acid and sterol biosynthetic pathways, and their complete repres sion is undesirable. Consequently, Roxl levels must be controlled tightly, and here we report that this is achieved by Roxl repressing its own expression.
MATERIALS AND METHODS
The wild-type yeast strain RZ53-6 ( MATcY, trpl-289, ura3- 52, leu2-3,-112, a d e l - 1 0 0 ) has been described ( BALASUBRA-
MANIAN et at. 1993)
.
The deletion derivatives of this strain were derived by transformation with indicated DNA fragments containing the appropriate deletion alleles: RZ53-6Aroxl as described previously (LOWRY and ZITOMER 1988); lU53-6Atupl as described previously ( ZWC et al. 1991 ) ; and RZ53-6AhupI using a SalI fragment from the plasmid phapl::URA3 (see below). RZ53-6Assn6was constructed us- ing an SphI-XbuI fragment from the plasmid pssn6::LEU2. The double deletion strains RZ53-6rlt1, RZ53-6s6t1, and RZ53-6JD1 were constructed in the same manner and con- tain the deletions AroxlAtupl, Assn6AtupI and AroxlA- hapl, respectively. All constructs were confirmed by Southern blot analysis (SOUTHERN 1975).
Yeast cells were grown at 30" in either rich media, WD or synthetic media (SC) lacking specific growth requirements
(ROSE et al. 1990). For long-term anaerobic growth, the me- dia were supplemented to 20 pg/ml of ergosterol and 0.2% of Tween80. Transformations were carried out as described ( KLEBE et al. 1983).
Plasmids: General techniques for plasmid constructions have been described ( SAMBROOK et al. 1989). Enzyme reac- tions were carried out as specified by the manufacturer. Refer- ences to 5 ' and 3' sequences refer to the coding strand.
YEP (112) ROXI-2 (9E10) contains two copies of the myc epitope 9E10 coding sequence inserted at the end of the ROXl coding sequence ( BALASUBRAMANIAN et al. 1993). The vectors YCplac22, YCplac33 and YIplac211 have been de- scribed ( GIETZ and SUGINO 1988).
YCp (22) ROXlH was constructed by the insertion of a 2.8- kb HindIII fragment containing the ROXl coding plus flank- ing sequences from YCpROXl ( LOWRY and ZITOMER 1988) into the HindIII site of YCplac22 ( GIETZ and SUGINO 1988).
YCp ( 22) roxl-G50D was isolated by in vitro hydroxylamine mutagenesis of the ROXl gene that will be described else- where. The plasmid is identical to YCp (22) ROXlH with the exception of a single base pair substitution at codon 50 that changes it from a glycine to an aspartate codon.
YCp ( 2 2 ) rod-tl04 is identical to YCp ( 2 2 ) ROXlH except that it contains a termination codon at codon 104, followed by an XhoI site and a deletion through codon 359. It was constructed by generating two PCR fragments as will be de- scribed elsewhere.
To generate YCp ( 2 2 ) ROXIX, an XHOI site was placed into the ROXl 5 ' region of the plasmid YCp ( 2 2 ) ROXlH by site- directed mutagenesis using the oligonucleotide 5'GCGAAA- AGAAAATATAATAGGS ' and the Clontech site-directed mutagenesis kit. This replaced the sequence from -258 to -274 with an XhoI site.
YCp (22) ROXl/ lucZ was constructed as follows. The 5 ' se- quences of ROXl from a Hind111 site 1.5-kb upstream from the coding sequence through to the translational initiation codon was amplified by PCR using the primers 5 ' -
CGCGGTACCCATTGTTGATTGTCTAACTGC-3' and the re-
verse-sequencing primer (New England Biolabs) and Y G p (22) ROXlH as the template. The product was digested with KpnI and HindIII and ligated, along with the @nI-EcoRI adapter ( 5 '-AATTCGCTGCAGGTAC-3 ' and 5 "CCTGAGCG 3 ' ) with YEp358R, a lacZ fusion vector (ATCC 3'7733). The resulting plasmid contained an in-frame fusion of the ROXl 1.5-kb upstream region plus codon one to the lac2 coding sequence. To provide 3 ' transcriptional termination signals, the plasmid was digested at the 5' end of the ROXl sequence with Hind111 and the middle of the lac2 coding sequences with S a d , and the 3.2-kb fragment was ligated into 7.4kb AAAAAAAAACTTTGTTTAAGACTCGAGTGAAAAGGA-
HindIII- SacI digested YCp ( 33) AZ6, an U B I / lac2 fusion plasmid constructed by inserting the 7-kb SmaI fragment from YCpAZ6 ( LOWRY et aZ. 1990) into the SmaI site of YCplac33. In the resulting plasmid, YCp ( 33) ROXl/ lucZ the ANBl 5 ' and coding sequences were replaced with the ROXl upstream region and translational initiation codon. Finally, to obtain a plasmid with the yeast TRPl marker, the 5-kb HindIII-SmaI fragment containing the ROXl/ lacZ/ ANBl fusion from YCp (33) ROXl/ lacZ was ligated into the HindIII-SmaI di- gested YCplac22 vector to produce YCp ( 2 2 ) ROXl/ lucZ.
The plasmid YCp ( 22) ROXl/ lacZAXXb contained a dele- tion from -259 to -430. It was constructed as follows. The XhoI site was inserted into the YCp ( 2 2 ) ROXl/ lac2 fusion plasmid as described above for YCp( 22) ROXIX. The re- sulting plasmid was digested with XhoI and XbaI, the ends were filled in and the plasmid was recircularized. The plasmid YCp ( 2 2 ) ROXlAXXb was constructed in a similar fashion from the YCp ( 22) ROXIX.
The plasmid YCppGl/ ROXl contained a fusion of the ROXl coding sequence to the GAL1 upstream region and transcriptional start site. It was constructed as follows. YCpp GALl is derived from pMB272 (from MARK JOHNSTON) by ligation of the 0.7-kb EcoRI-BumHI fragment containing the GALl promoter (which includes the UAS, TATA box and the RNA start site) into EcoRI-BumHI site of YCplac22. This plasmid was digested with BamHI and PstI. The ROXl coding sequence was excised from pmalE/ ROXl (Balasubramanian et ul. 1993) as a KpnI-PstI fragment and ligated to YCpPGALl in the presence of a BumHI/ KpnI adapter (S'GATCACA- TAATGGGTACJ' and 5'CCATTATGTJ'). The adapter pro- vided an ATG codon.
The plasmid phapl:: URA3 contains a deletion of the HAPl gene from a PstI site 211 bp upstream from the coding se- quence to a BamHI site 1348 bp into the coding sequence. It was constructed by inserting the 3-kb SalI-PstI and the 390- bp BamHESalI fragments from pKKHAPl ( a gift from L. GU-
ARENTE) into the corresponding YIplac211 sites, which con- tains the URA3 gene. The resulting plasmid, when linearized with SalI and transformed into yeast cells, replaced the de- leted HAPl sequences with the URA3 gene plus vector se- quences.
The plasmid pssnb::LEU2 was constructed by inserting the LEU2-containing 3.&kb PstI fragment from YEp13 ( BOTSTEIN et al. 1979) into the PstI-digested plasmid pBSSSN6SpXb, which contains a 6 k b SphI-XbuI insert containing the SSN6 locus subcloned into pBS+ (Stratagene). pssn6::LEU2 has almost the entire SSN6coding sequence deleted (319 bp from the 3' end remain) and replaced with the LEU2 gene.
RNA blots: RNA was prepared and RNA blots carried out
as described ( ZHANG et al. 1991). In all cases, equivalent amounts of RNA were loaded on the gel as determined by probing the blot with the actin gene. The preparation of the various gene-specific probes has been described ( LOWRY and ZITOMER 1988). A Betascope (Betagen) was used to quanti- tate the amount of radioactivity hybridized on the membrane, and the values were normalized to the radioactivity hybridized to the actin mRNA.
P-galactosidase assays: For these assays, cells were grown in YPD media overnight to midlog phase from light inocula either with vigorous shaking for aerobic growth or in sealed jars containing a BBL GasPack for anaerobic growth. Assays
Auto-Repression by Roxl 1151
TABLE 1
Effect of ROW and H A P I mutants on the expression of ROW/lacZ fusion
P-galactosidase activity
Strain" 0 2 N!2 Fold induction ( 0 2 / N 2 )
Wild type 27.5 2 4.6 1.8 2 0.7
Arox1 130 2 22 5.0 ? 1.6
Ahapl 12.0 ? 3.2 2.5 2 0.7
AroxlAhapl 63.3 2 13 4.9 2 1.6
15 26
13 4.8
See MATERIAIS AND METHODS for strains. Values are means ? SD.
the fusion protein was digested with factor Xa as described ( BALASUBRAMANIAN et al. 1993).
G e l retardation was carried out as described ( BALASUBRA- MANIAN et al. 1993) with the following DNA. The hypoxic consensus sequence synthetic DNA and the nonspecific syn- thetic DNA were prepared and labeled as described ( BALASU-
BRAMANIAN el al. 1993). The ROXl DNA probe was prepared by digesting YCp ( 2 2 ) ROXIX with XhoI (at -274), filling in the ends using cu"PdCTP, digesting with Eagi (at -499) and purifymg the 229-bp fragment. Per binding reaction, 0.32 ng
( 15,000 cpm) of DNA was added.
DNA sequence analyses: DNA sequence analyses was car- ried out by the dideoxy-chain termination method of SANCER
et al. ( 1977) using sequenase (US Biochemicals) and primer
DNAs synthesized with an Applied Biosystems DNA Synthe- sizer.
RESULTS
ROXl represses its own expression: To study the ex-
pression of R O X l , a gene encoding the hypoxic repres-
sor, we constructed YCp (22) R O X l / h c Z , a centro-
meric plasmid containing a fusion of 1.5 kb of the 5
'
ROXl sequence and the AUG translational initiation
codon to the h c Z coding sequence. This construct was
regulated as expected when transformed into wild-type
cells; Pgalactosidase activity was 15-fold higher in cells
grown aerobically compared with those grown anaero-
bically (Table 1 )
.
However, when the congenic Aroxlstrain carrying this plasmid was assayed for Pgalactosi-
dase activity, enzyme levels were increased substantially
under aerobic and, to a lesser extent, anaerobic growth
conditions (Table 1 )
.
These results suggested thatRoxl repressed its own synthesis.
To determine whether the R O X l / h c Z fusion re-
flected changes in the levels of ROXl RNA, we carried
out RNA blots using two mutant ROXl alleles that pro-
duce protein with no repressor activity. The first mu- tant, roxl-GSODcontained a single base pair substitution that resulted in an aspartate substituted for the wild-
type glycine at codon 50. This glycine is highly con-
served in the HMG DNA-binding domain ( BALASUBRA-
MANIAN et al. 1993). The second, roxl-tl04 contained
a nonsense mutation followed by a deletion that re-
sulted in termination of the coding sequence at position 104; the wild-type protein is 368 amino acids long. Both
mutant alleles were carried on centromeric plasmids
and when transformed into Aroxl cells, were incapable
of repressing expression of the hypoxic ANBI gene
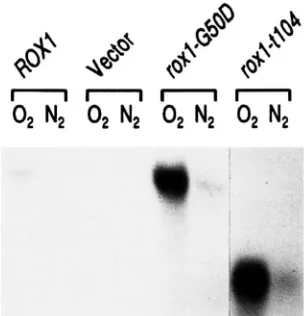
(data not shown). The results of a blot probed for
ROXl RNA are shown in Figure 1. The levels of ROXl
RNA were increased dramatically (6 and 9-fold, respec-
tively) in Aroxl cells carrying either of the mutant plas-
mids (lanes roxl-GSOD and roxl-tl04) compared to
those carrying a wild-type ROXl plasmid ( lanes R O X l )
.
As expected, the transcript from the deletion allele
(lanes roxl-1104) was much smaller than that in either
the wild type or point mutant. The derepression of the mutant RNA levels did not affect the oxygen induction of ROXl expression; as was found for the wild-type al-
lele, both mutant alleles showed substantially more
RNA accumulation in aerobic cultures (lanes 02) com-
pared with that in anaerobic cultures (lanes N 2 ) .
Roxl binds to its own upstream region: The consen-
sus sequence for Roxl binding, 5 'WATTGTT'CTC
(where
Y
represents pyrimidine) , has been proposed,""
FIGURE 1.-Overexpression of roxl mutant RNA. This RNA blot was carried out with RNA prepared from RZ53-6Aroxl transformants grown either aerobically (0,) or aerobically followed by 2 hr of anaerobic growth induced and maintained by bubbling nitrogen through the cultures (N,)
.
The cells were transformed with the following plasmids: YCp (22) ROXlH (lanes ROXl) ; YCplac22 (lanes Vector) ; YCp (22) roxl-G50D (lanes roxl-GSOD) ; and YCp (22) roxl-tl041152
-900 -880 - 8 6 0
- 8 4 0
- 8 2 0 -800
-780 - 7 4 0 - 7 6 0
-720 -700 - 6 8 0
-660 - 6 2 0 - 6 4 0
-600 -580 - 5 6 0
-540 -520 - 5 0 0
AAC TTGGGCATTT AGTCATGTAG CCGCCTAGCG AGCCTGGGTG CCGTCCTATC TCCTTTGTTC GTGCAAAGAG ACAGGAACAC ACACTGCGTT CTCTTGCGGC CGGTCTGGCG GACTCAGGGG TGCGGCGTTT GCTTAACCGG AGGGAATAAT AAAATCGGGG TGACGCAAGT ATGAAGTCAT GTGTGCTTAG CAATTACGTA GAGGGATTAG AAATAATAGT GTGCGGTTAT CGGAACCGGC TCTTGTTCCC GTTTAGAGCA ACCCAGGTGC AGGCGTACTT TAAAGTATTT TCTTTCTTTT TTTTCCTGCT ACTTACGCTA GGACGTGCGC AGCTCGAAAG CCGACGTCGG AGAGGCAGGT GATCTTCGCT
- 4 8 0 - 4 4 0 - 4 6 0
- 4 2 0
- 4 0 0 - 3 8 0
- 3 6 0 - 3 4 0
- 3 2 0
CGGCCACAAA TCCCCTGGAT ATCATTGGCC TGTCGAGGTA
T ~ C G T
GGAACTACCG GGAATTACTA TG-CAA TTQQAAATCT GGTAGGAAAA CCTTG-CTTGGCGATTGCTGACA AAGAAGAAAA GGGCCTATTG T T G n G C C T C T m T G T T O T T CTTCCT- -300
-CC GGTGTTCTTT GTGTCTTTTG TGTGTAGGTT CTTACTATTA TAGTGCTCTT TGCTATTATA TTTTCTTCGT TTTCACTTTG CGTAATGTAA CGGTCTTAAA CAAAGTTTTT
- 2 8 0 - 2 6 0
-240 -220
TTTTTTTCGC TCTTGCAT!PF TCCTTTTCTG CTCTATCTTA TTTGCTAATT GTAGTTTCAG
-180 -160
AAGTTTTACC TTAAATATAG CACTATTTTC CAGTTTTAAT GTTTCTTCTC ATTGCTTTCT
- 1 2 0
TTTATAATTT TCGCATATAA TTATACATTT ACGGTGTCTT AACTCTCCCT CTTCACCCCT
-60 -40
CATTATTCCA GAAAATACTA ATACTTCTTC ACACAAAAGA ACGCAGTTAG ACAATCAACA
1 2 0
ATG AAT CCT AAA TCC TCT ACA CCT AAG ATT CCA AGA CCC AAG AAC GCA TTT Met A s n P r o Lys Ser Ser T h r P r o Lys I l e P r o Arg P r o L y s A s n A l a P h e
I"-"- CTCGAG-""-I
- 2 0 0
- 1 4 0
- 1 0 0 -80
- 2 0
4 0
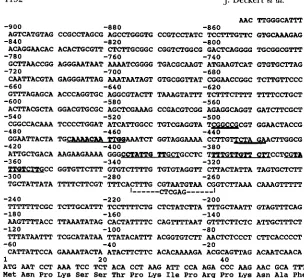
FIGURE 2.-Sequence of the ROXl upstream region. The best matches to the proposed hyp- oxic consensus sequence are underlined with the exact nucleotide matches in bold. The most 5' sequence is in the inverted orientation. The Ea$ and Xbd restriction sites referred to in the text are double underlined. The sequences that were deleted and replaced with an XhoI site in
YCp ( 2 2 ) ROXlX are indicated.
( LOWRY et al. 1990; BUUBRAMANIAN et al., 1993) and several candidate sequences are present in the region 5
'
to the ROXl coding sequence (Figure 2 ).
Because none of these were perfect matches, two lines of direct evidence were sought for Roxl binding. First, a deletion of the region containing these putative Roxl binding sites was constructed to determine its effect on ROXlexpression. Second, gel-retardation assays on the ROXl
upstream region containing these putative sites were performed using partially purified Roxl. As described below, the results indicated that Roxl is capable of bind- ing to its own regulatory region.
The effect of deleting the putative Roxl binding sites was tested in wild-type cells carrying a R O X l / 1acZfusion plasmid, YCp ( 2 2 ) R O X l / lacZ AXXb containing a de- letion from -259 to -430 (see Figure 2 )
.
Table 2 shows that &galactosidase levels were elevated greatly in suchtransformants grown aerobically as compared with the levels in cells carrying a wild-type ROXl / lacZ fusion. These elevated levels were comparable with those found in the ROXl deletion strain carrying a wild-type plas- mid. As expected for the loss of Roxl repression, anaer- obic expression was not dramatically increased. To de- termine whether the effect of the deletion was due to the loss of Roxl-dependent sites or the loss of a Roxl- independent repression site, the deletion plasmid was transformed into the congenic ROXl deletion strain and enzyme levels determined. If Roxl were capable of repressing expression from the deletion plasmid, we would expect even greater enzyme levels in the ROXl
deletion strain.
As
seen in Table 2, we observed no additional increase in P-galactosidase activity in these transformants over those determined for transformants carrying the deletion plasmid. Thus, at least the bulkTABLE 2
Effect of deletion of the putative Roxl binding sites on the expression of ROXl/lacZ fusion
~~ ~~
P-galactosidase activity'
Strain" Plasmid 0 2 N2
Wild type YCp (22) ROXl/lacZ 27.5 ? 4.6 1.8 f 0.7
Wild type YCp(22) ROXl/lacZAXXb 107 ? 14.4 4.5 ? 0.4
A roxl YCp (22) ROXl/ lacZ 130 ? 22.2 5.0 2 1.6
Aroxl YCp (22) ROXl/ lacZAXXb 102 -C 15.1 8.0 2 0.6
a See MATERIALS AND METHODS for strains.
Auto-Repression by Roxl 1153
ROXl
Consensus
Nonspecific"-
b a d
c;
ct
1
2 3 4
5
6 7 8 9 10 1112131415
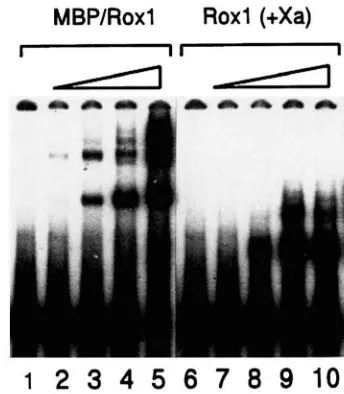
FIGURE 3.-Binding of the MBP/Roxl fusion to the ROXl upstream region. Gel retardation was carried out using the 229-bp XhoI-Eugi fragment from the ROXl upstream region (ROXI, lanes 1-5), the synthetic hypoxic consensus se-
quence 5'-GGG~CAGCCCATTGTTCTCCAG3' and 5'-
G G ~ G G C T C G A G A A C G G G C T G 3 ' (Consensus, lanes 6-10) and the synthetic DNA5'-TCGAGAAGAACAAAAGCT- TATl'TCTGAAGAAGACTTGGAGG3 ' and 5 "TCGAGCTC CAAGTCTTCTTCAGAAATAAGCTl"TCTTCTTG3' (Non- specific, lanes 11-15). Equivalent concentrations of each DNA (0.32 ng) was used. A protein extract containing par- tially purified MBP/Roxl fusion was titrated against each DNA as follows: lanes 1, 6 and 11, no protein; lanes 2, 7 and 12, 50 ng; lanes 3, 8 and 13, 100 ng; lanes 4 , 9 and 14, 200 ng; and lanes 5, 10 and 15,400 ng.
of the Roxl responsive sites are contained within the deleted region.
To determine whether the Roxl protein is capable of binding to the putative binding sites, gel-retardation experiments were carried out with a 229-bp labeled re- striction fragment containing sequences from -274 to -499 and Roxl expressed in and partially purified from
E. coli. The Roxl protein was expressed as a fusion to
the maltose binding protein (MBP)
,
and this fusion hasbeen shown to bind the hypoxic consensus sequence
specifically ( BALA~UBRAMANIAN et al. 1993). As is evi-
dent in Figure 3, the fusion protein forms at least three
different sized complexes with the ROXl regulatory re-
gion. The migration of the free DNA can be visualized
in lane 1 ( n o protein added to the reaction mix), and
lanes 2-5 show the complexes formed with increasing protein concentration. For comparison, the same pro- tein concentrations were allowed to form complexes with a synthetic DNA containing the hypoxic consensus sequence (lanes 6-10) and with a nonspecific synthetic
DNA which lacked any homology ( lanes 1 1-15 )
.
Addi-tionally, competition experiments between the labeled
ROXl 229-bp fragment and unlabeled consensus se- quence or unlabeled nonspecific DNA confirmed the
specificity (ANA RODRIGUEZ-TORRES, unpublished re-
It should be noted that the different migration rates
of the complexes seen in Figure 3 probably do not
reflect the sizes of the complexes, but rather the loca- tion of the binding sites on the fragment. We have
found that, like other HMG proteins ( GIESE el al.
1992), Roxl bends DNA when it binds (A. M. RODRI-
GUEZ-TORRES, unpublished results). As a consequence, a complex in which a single Roxl molecule is bound to a site close to the end of the fragment will migrate faster than a complex in which the protein is bound to
a site in the middle of the fragment ( Wu and CROTHERS
1984). This behavior also is reflected in the appearance of a faster migrating species at higher Roxl concentra- tions, presumably because a site near the end binds the protein less well than one in the middle.
To demonstrate that the complex formed with the
ROXl regulatory region was a result of Roxl binding, the partially purified protein extract was treated with the protease Xa before DNA was added. This protease cleaves the fusion protein at the MBP-Roxl junction. The result of this treatment is seen in Figure 4. The untreated protein (lanes 2-5) formed a slower-migrat- ing set of complexes than did the Xa treated protein
(lanes 7-10) as would be expected if binding were due sults).
MBP/Roxl
Roxl (+Xa)
"
Deckert
GLUCOSE
GALACTOSE
I i
w
YCppGlROXl YCppGlROXl
to the MBP/Roxl fusion in the untreated samples and
to free Roxl in the Xa-treated samples. Thus, the results
of these in vitro binding experiments provide very
strong evidence that the repression of ROXl expression
is mediated by a direct binding of the Roxl repressor. Overexpression of Roxl inhibits cell growth: We be- lieve that the self-repression of Roxl serves the purpose of preventing complete repression of certain hypoxic
genes, such as HEM13, ERG11 and OLEI, whose prod-
ucts are required at lower levels during aerobic growth
for heme and membrane biosyntheses. Even were these
genes to contain poor Roxl binding sites, high levels of Roxl still would cause complete repression. To test this hypothesis, we transformed wild-type cells with a plasmid carrying the deletion of the Roxl binding sites, YCp (22) ROXlA XXb, which should result in a fourfold
increase in Roxl cellular levels (see Table 2 ) . We de-
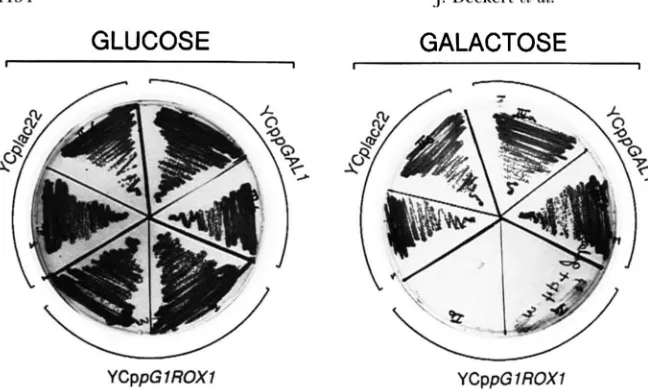
tected no effects of Roxl overexpression on the growth of these transformants under laboratory conditions. To further increase Roxl levels, we expressed Roxl from
the GALl promoter using the plasmid YCppGl/ ROXI.
This plasmid contains a fusion of the entire upstream region and transcriptional start site of the GALl gene
to the entire coding sequence of ROXI. Northern and
western blot analyses demonstrated that Aroxl cells car-
rying this plasmid and incubated on galactose produced
extremely high levels of ROXl mRNA and protein com-
pared with wild-type cells (data not shown). These
transformants did not grow in liquid galactose media, and their lack of growth is evident on the plates pre- sented in Figure 5. Cells transformed with a wild-type
ROXl plasmid or the vector grew well on both glucose and galactose, but cells carrying the galactose-inducible
ROXl could not grow on galactose. The addition of either heme or the membrane components ergosterol
and tween 80 did not restore growth (data not shown )
.
Although we can not rule out a specific effect of Roxl overexpression on galactosedependent growth or the possibility that there are weak Roxl binding sites u p
FIGURE 5.-Effect of Roxl overexpression
on cell growth. RZ53-6Aroxl was trans-
formed with the following plasmids: the
vector YCplac22; YCppGALl, the vector with the GAL1 promoter; and YCppGl/
ROXl, the fusion of the GAL1 promoter to
the ROXI coding sequence. Two indepen-
dent transformants of each were streaked
on selective media lacking tryptophan
(ROSE et al. 1990) containing either 2%
glucose or 2% galactose.
stream of essential genes not normally regulated by Roxl, the data do support the hypothesis that overex- pression of Roxl causes complete repression of a set of
hypoxic genes that are required for aerobic growth, and
that the self-repression by Roxl has evolved to prevent such an occurrence.
Hapl regulation of ROXI: Hapl is a hemedepen-
dent transcriptional activator and was reported to be
the activator of hemedependent ROXI expression
( KENC 1991). Recently, USHINSKY and KENC (1994) suggested that Hapl also might serve to repress anaero- bic ROXl expression. To further investigate this regula-
tion, we constructed HAP1 deletion and ROXI,
HAP1
doubledeletion strains and assayed ROXl expression.
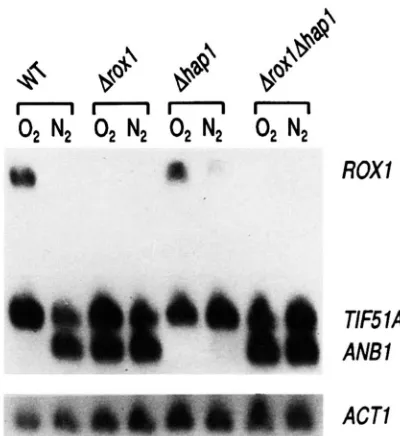
Figure 6 shows the results of a blot of RNA prepared
from wild type ( W T ) , Aroxl, Ahapl and AroxlAhapl
cells grown under aerobic ( 0 2 ) or anaerobic ( N2) con-
ditions. As expected, ROXl RNA levels were reduced
aerobically in the Ahupl mutant, but increased some-
what anaerobically, confirming that Hapl can function as both an activator and repressor. Interestingly, how- ever, there was still a significant difference between aer- obic and anaerobic RNA levels, indicating that Hapl was not solely responsible for heme regulation.
The effect of the Ahupl mutation on ROXl expres-
sion was further investigated using the ROXI/ lucZ fu-
sion (Table 1 )
,
and the results of the enzyme assaysagreed with those of the RNA blots. Enzyme levels were reduced more than twofold in aerobically grown cells
and increased slightly,
-
1.4fold, in anaerobicallygrown cells in the Ahapl mutant compared with wild-
type cells. This same affect was observed when enzyme
levels in the Aroxl mutant were compared with those
in the AroxlAhapZ double mutant, although the overall
levels were much higher because of the relief from Roxl
repression. Thus, the net effect of the HAP1 deletion
was a reduction in extent of aerobic induction from 15- to 4.8-fold. Consequently, although Hapl does contrib
Auto-Repression by Roxl 1155
for the entire effect. Also, the high levels of expression
observed both aerobically and anaerobically in the dou- ble mutant indicates that Hapl can not be the sole
transcriptional activator of ROXl.
The effect of the HAP1 deletion on ANBl and
llF51A
expression: The increase in the anaerobic expression of ROXl in the Ahupl deletion strain, although small,can be seen indirectly in the effect of the Ahupl muta-
tion on the expression of an hypoxic gene. The blot in
Figure 6 also was probed for the transcripts of the heme-
induced/ hypoxic homologues TF51A/ ANBl. These
genes encode eIF5A homologues, putative translational
initiation factors ( METHA et ul. 1990; SCHNIER et ul.
1991 ) and are so similar that an ANBl probe visualizes
both transcripts.
As
seen in Figure 6, ANBl RNA levelswere reduced greatly in Ahupl cells grown anaerobi-
cally. This anaerobic repression of ANBl expression
could have been due to the small level of derepression of ROXl in the Ahupl mutant or to a Roxl-independent
pathway. The epistatic relationship between ROXl and
HAP1 in a doubledeletion mutant distinguished be-
tween these possibilities.
As
can be seen in this figure,the double mutant accumulated ANBl RNA at high
levels both aerobically and anaerobically, characteristic
of the Aroxl mutant (compare Aroxl and AroxlAhupl
lanes). Thus the effect of the Ahupl mutation on ANBl
expression can be explained as an indirect effect caused
through increased anaerobic expression of ROXl.
This anaerobic repression by Hapl also can be seen
in the effect of the Ahupl mutation on TF51A RNA
levels, which were increased in Ahupl cells grown an-
aerobically compared with the levels detected in anaer-
obically grown wild-type and Aroxl cells. Interestingly,
here the effect of the Ahupl mutation was epistatic to
the Aroxl mutation, which had no effect on TF51A
RNA levels, as can be seen by comparing anaerobic
RNA levels in the AroxlAhupl double mutant with that
in either single mutant. These results indicate that
Hapl is responsible for anaerobic repression of both
ROXl and TF51A, but we can not determine whether this is a direct or indirect effect from these data.
The effect of a TUPl and SSN6 deletions on ROXl
expression: We reported previously that deletion of the
TUPl gene caused increased expression of ROXl
(ZHANG et ul. 1991 )
.
Because Roxl repression of thehypoxic genes requires the general repressors Tupl and
Ssn6 and because these present studies indicate Roxl represses its own expression, it is possible that the effect
of the TUPl deletion can be simply due to its require-
ment for Roxl self-repression. If this were the case,
then the increase in ROXl expression in a AroxlAtupl
double mutant should be the same as that in either
single mutant because both mutations would disrupt
the same pathway. We constructed both A t u p l and
AroxlAtupl strains that were congenic with the wild
type and Aroxl deletion strains used in these studies,
ROX7
ACTl
RGURE 6.-Effect of HAP1 deletion on ROXl, ANBl and TF51A RNA levels. This RNA blot was camed out with RNA prepared from RZ53-6 (lanes W T ) , RZ53-6Arox1 (lanes A r o x l ) , RZ53-6Ahupl (lanes Ahupl) and RZ53-6JD1 (lanes AroxlAhupl) grown either aerobically ( 02) or anaerc- bically ( N2) as described in Figure 1 . The blot was probed for ROXl, ANBl and TF51A RNAs simultaneously, then stripped and probed for actin RNA ( A C T l )
.
The position of the different RNA species are indicated to the right of the blot. No ROXl mRNA is visible in third, fourth, seventh and eighth lanes because these cells contain a ROXl deletion.and transformed them with the ROXl /lacZfusion. Also,
since Tupl functions in a complex with Ssn6, we also
constructed congenic Assn6 and Assn6Atupl strains to
determine whether the effects of Tupl and Ssn6 were equivalent. The results of &galactosidase assays are pre- sented in Table 3. Under aerobic growth conditions the
Atupl deletion strain accumulated about twofold more enzyme than did the wild type. This increase was less
than observed for the Aroxl mutant. The increase in
enzyme levels observed in AroxlAtupl double mutant
was about the same as that seen in the Aroxl strain,
indicating that Tupl and Roxl act through the same pathway for aerobic repression.
Surprisingly, the results under anaerobic growth con- ditions were quite different. Enzyme levels were greatly
increased in the Atupl strain compared with either wild
type (9-fold) or the Aroxl mutant (3.3-fold). Levels in
the AroxlAtupl double mutant were also much greater
than that for the wild type or Aroxl mutant. We inter-
pret these results as indicating that at least part of the
hemedependent expression of ROXl is due to an un-
identified anaerobic repressor that requires Tupl for
its function. It also should be noted, that the increased
anaerobic expression seen in the A t u p l mutants indi-
cates that there is an activator of ROXl expression that
TABLE 3
Effect of TUPl mutants on the expression of ROXl/lacZ fusion
P-galactosidase activity’
Strain” 0 2 N2 Fold induction ( 0 2 / N 2 )
Wild type 27.5 t 4.6 1.8 5 0.7 15
A r o x l 130 ? 22 5.0 t- 1.6 26 A t u p l 53.0 t 8.9 16.3 5 2.6 3.2 A r o x l A t u p l 136 t- 5.7 21.9 2 2.8 6.2
A s s n 6 146 t- 18 19.0 2 3.4 7.7
A s s n 6 A t u p l 44.9 5 7.0 17.7 2 2.9 2.5
a See MATERIALS AND METHODS for strains.
bThe values for the wild-type and A r o x l strains are the same as those presented in Table 1, representing averages of experiments carried out many times during the course of this study. Values are means 5 SD.
Interestingly, the SSN6 deletion caused a greater in- crease in aerobic ROXl expression than that seen for the TUPl deletion and about the same as the increase observed for the ROXl deletion. The Assn6Atupl dou- bledeletion mutant contained equivalent P-galactosi- dase enzyme levels in aerobic cells as the A t u p l deletion indicating that effect of the TUP1 deletion is epistatic to that of the SSN6 deletion. Whereas these results sug- gest that Tupl and Ssn6 may not play equivalent roles in the aerobic self-repression of ROXl expression, it is possible that these results reflect an indirect effect of these repressors. Both T W l and SSN6 mutants are ex- tremely flocculent and grow slower than wild-type cells. Even with vigorous shaking, we can not be certain that cells contained within large clumps are fully aerobic. Consequently differences in ROXl expression between the A t u p l and Assn6 mutants may result from subtle differences in flocculence and, thereby, aeration. Un- der anaerobic conditions, the effects of the SSN6, TUP1
and SSN6-TWl deletions were similar indicating that at least under these conditions, Tupl and Ssn6 act through a common pathway.
DISCUSSION
This study represents our initial attempts to answer the question, “what regulates the regulators?” in this case, the Roxl repressor of hypoxic genes. We have obtained evidence for three, and probably four, regula- tory elements: the heme-dependent Hapl transcrip- tional activator, previously implicated in ROXl regula- tion ( KENG 1992; USHINSKY and KENG 1994) ; a Ssn6/
Tupl-dependent repressor that repressed anaerobic ex- pression; Roxl itself that repressed its own expression; and an unidentified transcriptional activator, which is responsible for the Hapl-independent aerobic expres- sion of ROXl observed in the Ahapl deletion strains and the anaerobic expression observed in the A t u p l
strains. Thus, the regulatory region of ROXl appears to be quite complex, and its analysis may provide some
interesting insights into how different elements interact to regulate transcription.
Hapl regulation: We showed previously that ROXl
is transcriptionally induced by heme (LOWRY and ZI-
TOMER 1988), and KENG (1992) reported that, in a
HAPl deletion strain, ROXl expression was uninduced. Our results with a hapl-deletion strain led to a different conclusion; whereas Hapl certainly contributes to
ROXZ induction, it is responsible for only part of the heme response. The difference between our findings may be related to growth conditions. KENG performed her experiments in a Ahernl background and grew cells in low us. high 6-aminolevulinate to mimic anaerobic us. aerobic conditions. It is possible that the low level of 6-aminolevulinate was still high enough to derepress the second regulatory mechanism we found.
We also confirmed USHINSHY and KENG’S (1994)
findings that Hapl was responsible for anaerobic re- pression as well as aerobic induction of ROXl
.
Although in both studies this increased anaerobic ROXl expres- sion appeared to be small, its effect was greatly magni- fied in the expression of the ANBl gene; anaerobicANBZ RNA levels were repressed almost completely in the Ahapl strain. Because the ROXl deletion was epi- static to the HAPl deletion in the double mutant, we concluded that the anaerobic repression of ANBl must be due to the accumulation of Roxl. It should be noted that KENG (1992) found that in a HAPl mutant heme- dependent HEM13 expression was lower than wild type in the presence of heme in agreement with the findings presented here with aerobic ANBl expression, but, un- like our results with anaerobic ANBl expression,
Auto-Repression by Roxl 1157
We found that this anaerobic repression by Hapl
was also manifest in the accumulation of TlF51A RNA.
Deletion of HAP1 resulted in increased anaerobic and
somewhat decreased aerobic TlF51A RNA levels indicat-
ing a role for Hapl in TF51A repression anaerobically.
This is not the first report of an apparent Hapl-anaero-
bic repression activity. ZITOMER et al. (1987) observed
that point mutations in the Hapl binding site of the
CYC7 gene caused both decreased aerobic and in- creased anaerobic expression. Thus Hapl may play a
role as a repressor in the absence of heme as well as its
better characterized role as a heme-dependent tran- scriptional activator. These results do not distinguish between a direct or indirect effect of Hapl. However,
recently FITLOVICH et al. ( 1993) reported that in crude
extracts Hapl bound to its target UAS both in the pres-
ence and absence of hemin and that the complex
formed in the absence of hemin migrated more slowly. Perhaps this slower Hapl-containing complex repre- sents a repression complex.
Tupl /Ssn6 regulation: The Tupl / Ssn6 complex
functions as a general repressor of a number of systems
including the hypoxic genes ( ZHANC et al. 1991; BALA-
SUBRAMANIAN et al. 1993) , a-specific genes ( LEMONTT et
al. 1980; KELEHER et al. 1992) and catabolite-repressed
genes (SCHULTZ and CARLSON 1987; TRUMBLY 1988;
WILLIAMS and TRUMBLY 1990). In each of these cases,
the complex functions through specific DNA-binding
proteins that control when repression occurs: Roxl
(ZHANC et al. 1991; BALA~UBRAMANIAN et al. 1993), a2
( LEMONTT et al. 1980) and Migl ( NEHLIN et al. 1991 )
,
respectively for the three examples above. Although
these cases represent the best documented roles of
Tupl / Ssn6, there must be a number of other repres-
sion systems that utilize this complex based upon the
pleiotropy of TUPl or SSN6 mutations, which include
flocculence ( FUJITA et al. 1990) and effects on plasmid
stability ( THRASH-BINGHAM and FANGMAN 1989). We
believe we have uncovered another role for this repres- sion complex in the anaerobic repression of Roxl.
There are clearly two roles for Tupl
/
Ssn6 in theregulation of ROXl expression. The first involves the
self-repression by Roxl, which, based upon the require- ments for Roxl repression of the hypoxic genes, would
be expected to require the Tupl / Ssn6 complex. This
was seen to be the case here in the increased aerobic
expression of ROXl in a TUPl or a SSN6deletion strain.
It is evident that the aerobic Tupl effect was mediated through Roxl, as the derepression in a double mutant
AroxlAtupl was not additive, but the same as in the &-ox1 single mutant,
The second role for Tupl / Ssn6 involves anaerobic
repression and was demonstrated by the ninefold in-
crease in anaerobic ROXl expression in the Atupl,
Assn6 or double mutant. We concluded that this dere- pression was independent of Roxl because the anaero-
bic derepression in a Aroxl mutant was very small and
the larger Atupl effect was epistatic to the much weaker
A r o d effect in the AroxlAtupl double mutant. This
pattern of anaerobic ROXl expression in these mutants
is very different than that for the aerobic expression
discussed above and leads to the conclusion that there
is a anaerobic repressor that requires Tupl / Ssn6 for
its function. We believe that the combination of the Hapl induction/ repression and the anaerobic repres- sion by this yet to identified repressor is responsible for
the heme regulation of ROXl.
A constitutive transcriptional activator? Two lines of
evidence suggest that Hapl is not the only transcrip-
tional activator of ROXl expression. First, in Ahapl mu-
tants, the reduction of aerobic expression of ROXl was
not dramatic; for example, the high level of P-galactosi-
dase activity from the ROXl/ lucZfusion in the AroxlA-
hap1 mutant was only twofold lower than that in the
Aroxl mutant (see Table 1 )
.
Therefore, some otherelement must be driving ROXl expression. Second, in
the TUPl deletion strains, significant levels of anaerobic
expression of ROXl were observed. Because Hapl is
only known to activate transcription aerobically ( GUA-
RENTE and FOSBURG 1989), these results suggest an-
other activator is functioning. We do not have sufficient
data to determine whether this activator functions con- stitutively or contributes in some way to the heme regu-
lation of ROXl.
Roxl self-repression: We have demonstrated here that Roxl represses its own expression. This effect does
not play a role in heme regulation of ROXl expression;
indeed the difference between aerobic and anaerobic expression is greater in the absence of this self-repres- sion. Rather we believe that this self-repression is re- quired to prevent the accumulation of excess Roxl in the cell. Complete aerobic repression of at least some of the hypoxic genes would be detrimental to the cell.
For example, the product of the HEM13 gene is re-
quired for heme synthesis, the ERG11 and CPRl gene
products are required for sterol synthesis and the OLE1
product is required for fatty acid synthesis; complete aerobic repression of the expression of these genes would leave the cell respiratory deficient and increase
its growth requirements ( ZITOMER and LOWRY 1992).
On the other hand, genes such as ANBl, COX56 and
AAC3 have aerobically expressed homologues and their
expression appears to be completely repressed ( ZI-
TOMER and LOWRY 1992). This differential repression can be achieved by designing binding sites with varying
affinities for Roxl, but such a system would require that
the cellular concentrations of Roxl be kept at a level that would allow the tight sites to be occupied all the time whereas the weak sites would be occupied only part of the time. By placing weak sites upstream of the
hibit cell growth, although growth was not restored by supplementing the media with the products of the known hypoxic genes. It is possible that there are addi- tional, essential functions repressed by Roxl or that when sufficiently overexpressed Roxl gratuitously re- presses other genes. In any event, it appears to be advan- tageous to the cell to control Roxl levels.
This study was supported by National Institute of General Medical Sciences grant GM-26061.
LITERATURE CITED
ANDREASEN, A., and T. STIER, 1953 Anaerobic nutrition of Saccharc- myces cereuisiae. I. Ergosterol requirement for growth in defined medium. J. Cell. Comp. Physiol. 41: 23-36.
ANDREASEN, A., and T. STIER, 1954 Anaerobic nutrition of Saccharo- myces cereuisiae. 11. Unsaturated fatty acid requirement for growth in defined medium. J. Cell. Comp. Physiol. 4 3 271-281. BAIASUBRAMANIAN, B., C. V. LOWRY and R. S. ZITOMER, 1993 The
Roxl repressor of the Saccharomyces cereuisiae hypoxic genes is a specific DNA-binding protein with a high-mobility-group motif. Mol. Cell. Biol. 13: 6071-6078.
BOTSTEIN, D., S. C. FALCO, S. E. STEWART, M. BRENNAN, S. SCHEER et
al., 1979 Sterile host yeasts ( S H Y ) : a eukaryotic system of biologi- cal containment for recombinant DNA experiments. Gene 8:
FUJITA, A,, S. MATSUMOTO, S. KUHARA, Y. MISUMI and H. KOBAYASHI, 1990 Cloning of the yeast SFL2 gene: its disruption results in pleiotropic phenotypes characteristic for tup1 mutants. Gene 89: 93-99.
FYrl.OVICH, S., M. GERVAIS, C. AGRIMONTI and B. G u m , 1993 Evi- dence for an interaction between CW1 (HAP1 ) activator and a cellular factor during hemedependent transcriptional regula- tion in the yeast Saccharomyces cereuisiae. EMBO J. 12: 1209-1218. GIESE, R, J. COX and R. GROSSSCHEDL, 1992 The HMG domain of
Lymphoid Enchancer Factor 1 bends DNA and facilitates assem- bly of functional nucleoprotein structures. Cell 69: 185-195. GIETZ, R. D., and A. SUGINO, 1988 New yeast-Escha'chia coli shuttle
vectors constructed with in uitro mutagenized yeast genes lacking six-base pair restriction sites. Gene 7 4 527-534.
GUARENTE, L., and S. L. FOSBURC, 1989 Communication between mitochondria and the nucleus in the regulation of cytochrome genes in the yeast Saccharomyces cereuisiae. Annu. Rev. Cell Biol. 5 153-180.
KELEHER, C. A,, M. J. REDD, J. SCHUL.TZ, M. CARISON and A. D. JOHN- SON, 1992 Ssn6-Tup1 is a general repressor of transcription in yeast. Cell 68: 709-719.
KEN(;, T , 1992 HAPl and ROXl form a regulatory pathway in the repression of HEM13 transcription in Saccharomyces cereuisiae.
Mol. Cell. Biol. 12: 2616-2623.
KLEBE, R. J., J. V. HARRISS, 2. D. S wand M. G. DOUGLAS, 1983 A general method for polyethyleneglycol-induced genetic transfor- mation of bacteria and yeast. Gene 2 5 333-341.
LABBE-BoIs, R., and P. LABBE, 1990 Tetrapyrrole and heme biosynthe- sis in the yeast Saccharomyces cermisiue, pp. 235-285 in Biosynthesis
of Heme and Chlorophylls, edited by H. A. DAILEY. McGraw-Hill Publishing, New York.
LEMONTT, J. F., D. R. FUGIT and V. L. MACKAY, 1980 Pleiotropic mutations in the TUP1 locus that affect the expression of mating- type dependant functions in Saccharomyces cerevisiae. Genetics 9 4 899-920.
17-24.
LOWRY, C. V., and R. S. ZITOMER, 1988 ROXl encodes a heme- induced repression factor regulating ANBl and CYC7of Saccharo- myces cerevisiue. Mol. Cell. Biol. 8: 4651-4658.
LOWRY, C. V., M. E. CERDAN and R. S. ZITOMER, 1990 A hypoxic consensus operator and a constitutive activation region regulate the ANBl gene of Saccharomyces cereuisiae. Mol. Cell. Biol. 10: 5921-5926.
MATTOON, J. R., W. E. LANCASHIRE, H. K. SAUNDERS, E. CARVAJM,, D. R. MALAMUD et al., 1979 Oxygen and catabolite regulation of hemoprotein biosynthesis in yeast, pp. 421-435 in Biochemical
and Clinical Aspects of Oxygen, edited by W. S. CAUGHEY. Academic Press, New York.
MEHTA, K. D., D. LEUNC, L. LEFEBVRE and M. SMITH, 1990 The ANBl
locus of Saccharomyces cermisiaeencodes the protein synthesis initi- ation factor eIF4D. J. Biol. Chem. 265: 8802-8807.
NEHLIN, J. O., M. CARLBERG and H. RONNE, 1991 Control of yeast GAL genes by MIGl repressor: a transcriptional cascade in the glucose response. EMBO J. 10: 3373-3377.
ROSE, M., and D. BOTSTEIN, 1983 Construction and use of gene fusions to lac2 (PGal) that are expressed in yeast. Methods Enzy- mol. 101: 167-180.
ROSE, M. D., F. WINSTON and P. HIETER, 1990 Methods in Yeast Genetics, A Laboratory Course Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
SAMBROOK, J., E. F. FRITSCH and T. MANIATIS, 1989 Molecular Cloning:
A Labmatory Manual, 2nd Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
SANGER, F., S. NICKLEN and A. R. COWLSON, 1977 DNA sequencing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA. 7 4 5463-5467.
SCHNIER, J., H. T. SCHWEI-BERCER, Z. SMET-MCBRIDE, H. A. KANC and J. W. B. HERSHEY, 1991 Translation initiation factor 5A and its hypusine modification are essential for cell viability in the yeast
Saccharomyces cereuisiae. Mol. Cell. Biol. 11: 3105-3114.
SCHULTZ, J., and M. CARLSON, 1987 Molecular analysis of SSN6, a gene functionally related to the SNFl protein kinase of Sacrharc- myces cereuisiae. Mol. Cell. Biol. 7: 3637-3645.
SOUTHERN, E. M., 1975 Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98: 503- 517.
THRASH-BINCHAM, C., and W. L. FANGMAN, 1989 A yeast mutation that stabilizes a plasmid bearing a mutated ART1 element. Mol. Cell. Biol. 10: 809-816.
TRUMBLY, R. J., 1988 Cloning and characterization of the CYCX gene mediating glucose repression in yeast. Gene 73: 97-111. USHINSKY, S. C., and T. KENc, 1994 A novel allele of HAPl causes
uninducible expression of HEM13 in Saccharomyces cereivisiae. Ge- netics 136: 819-831.
WILI.LAMS, F. E., and R. J. TRUMBLY, 1990 Characterization of TUPI, a mediator of glucose repression in Saccharomyces cerevisiae. Mol. Cell. Biol. 10 6500-6511.
WU, H.-M., and D. M. CROTHERS, 1984 The locus of sequence-di- rected and protein-induced DNA bending. Nature 308: 509-513.
ZHANG, M., L. S. ROSENBLSIM-VOS, C. V. LOWRY, K. A. BOA= and R. S. ZITOMER, 1991 Ayeast protein with homology to P-subunits of G proteins is involved in control of heme-regulated and catab olite-repressed genes. Gene 97: 153-161.
ZITOMER, R. S., and C. V. LOWRY, 1992 Regulation of gene expression by oxygen in Saccharomyces cermisiae. Microbiol. Rev. 56: 1-1 1. ZITOMER, R. S., J. W. SELLERS, D. W. MCCARTER, G. A. HASTING, P.
WICK et al., 1987 Elements involved in oxygen regulation of the
Saccharowyces cermisiae CYC7gene. Mol. Cell. Biol. 7: 2212-2220.