RESPONSE TO WOUNDING1
C. S. LEVINGS I11 AND C . W. STUBERZ
Department of Genetics, North Carolina State University, Raleigh, North Carolina 27607
Manuscript received July 12, 1971 Revised copy received September 16, 1971
ROWNING is a common degradative phenomenon of injured cIr senescent
'
vegetable tissue (SZENT-GYORGYI 1969). I t is due to enzymatic oxidation of polyphenolics resulting in quinones which form brown colored complexes with protein. When fresh corn silks (stigmas) were ground in a n aqueous solution and allowed to stand, it was observed that the brei normally turned brown. Occasionally, ground silks from certain inbreds were found which did not turn brown hut remained yellow-green (i.e., no change in color). Later, it was learned that the browning or nonbrowning phenotypes could be determined on the plant by cutting back fresh silks, as in preparation for controlled pollination, and observing the cut ends after about 15 minutes. In this instance, cut ends become brown in the browning phenotype and remain unchanged in the ncnbrowning. The distinction between browning and nonbrowning types is also evident with senescent silks of mature ears. In this case, senescent silks from the browning type appear very dark brown while nonbrowning are tan in color.This study was conducted to determine the inheritance and chemical basis of the browning and nonbrowning response to wounding of corn silks. W e will show that the browning response is controlled by a single dominant locus which per- mits, primarily, the synthesis of dihydroxyl flavoaes by the silks and that when the silks are injured these flavones are enzymatically oxidized resulting in brown pigment formation. The nonbrowning type will he shown to be due to a recessive allele which when homozygous blocks the synthesis of the dihydroxyl flavones. Absence of the flavone substrates precludes brown pigment formation in response to wounding.
MATERIALS A N D METHODS
Inbred lines T61 and NC232, the browning type, were crossed with the nonbrowning inbreds, Kys and NC236, respectively. The resulting F,'s were backcrossed t o each of the parents. Field grown backcross progenies were scored for the browning and nonbrowning phenotype by cutting back fresh silks and observing the cut silk ends approximately 15 min later.
Contribution from Department of Genetics, North Carolina Agriculture Experiment Station, Raleigh and Plant Science Research D:vision, Agricultural Research Service, U.S. Department of Agriculture, cooperating. Paper No. 3507 of the Journal Series. The work was supported i n part by Cooperative Agreement No. 12-14-100-9798(34) between the N . C . Agricultural Experiment Station and the Plant Science Research Division, Agricultural Research Service, U.S. Department of Agriculture. T h e use of trade names i n this publication does not imply endorsement by the North Carolina Agricultural Expenment Station of the products named, nor criticism of s.milar ones not mentioned.
Associate Professor of Genetics, N.C. State Unirersity: and Research Geneticist, Plant Science Research Division, Agricultural Research Service, U.S. Department of Agriculture. Raleigh, N.C. 27607.
Silk samples for chemical analyses were taken from ear shoots from 1 to 6 days after silk
emergence. Care was exercised to avoid insect damaged and dying silks. Husks were stripped away and all silks were removed from the ear to form the sample.
Crude samples of silk phenols free of enzymes were prepared by grinding 5 g of silk in 5 ml of 10% perchloric acid with a chilled mortar and pestle. After grinding, 20 ml of H,O was added and the brei was allowed to stand 20 min in ice. The mixture was then centrifuged for 15 min a t 17,300 x g. The supernatant fraction was decanted and p H was adjusted to 5.0 with KOH. This solution was stored on ice for 10 min, then centrifuged for 15 min a t 17,300 X g, and the supernatant fraction saved.
Crude samples of silk enzymes free of phenols were prepared by grinding silks in PVP
(polyvinylpyrrolidone; Polyclar AT, General Aniline and Film Corp.) (LOOMIS and BATTAILE 1965) at the rate of 1.5 g per gram of silks in 5 ml of H,O. The mixture was centrifuged a t 17,300 x g for 15 min and the supernatant fraction decanted and stored on ice. PVP was purified by the method described by ANDERSEN and SOWERS (1967).
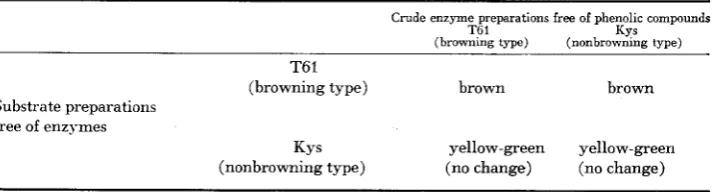
Crude enzyme preparations free of phenolic compounds (PVP treatment) and phenolic prepa- rations free of enzymes (perchloric acid treatment) from the browning line T61 and the non- browning line Kys were combined in four possible combinations (Table 2). Combinations were made so that amounts of enzyme and phenolic preparation mixed wire derived from fresh silks of similar weight. Mixtures were stirred, allowed to stand for 15 min and then classified for brown pigment formation
A combination of column and paper chromatography was used in isolating unknown phenolic compounds. Fresh corn silk samples were ground with a Waring blender in an acidified methanol solution (80 methanol: 19 H,O: 1 HCI, v/v/v) a t the rate of 3 ml of solvent per gram of fresh silk. The mixture was stored from 1 to 3 days in a refrigerator and then filtered and concentrated with a rotary evaporator. These samples were introduced to a column with PVP adsorbent and eluted with 60% ethanol. PVP was prepared for chromatography by repeated washing in water and 60% alcohol until clear. Although the PVP was not sieved, many fine particles were dis- carded by pouring off the liquid before the finest particles had s d e d after stirring. Fifteen-ml fractions were collected on a fraction collector and similar fractions, based on paper chromatog- graphy, were combined. Column chromatography gave only partial separation of phenolic comaounds.
Paper chromatography methods for flavonoids are well established (SEIKEL., 1962). TWO-
dimensional paper chromatography in two solvent systems, (1 ) n-butanol-acetic acid-water
(BAW 4:1:5, upper phase) and (2) 15% acetic acid (HOAc) on Whatman No .I paper was used to determine total flavonoids. Isolation and purification of compounds was carried out with Whatman No. 3 MM paper in either one or two dimensions. In one-dimensional chromatography mixtures were banded across the width of paper and run in an appropriate solvent for separation. Compounds were visualized on paper with visible and ultraviolet light and the following reagents: (1) ammonia, (2) 1% aluminum chloride solution in ethanol, and (3) Benedict’s solution. Ammonia fumes and ultraviolet light were employed where a nondestructive visualization was necessary. Benedict’s reagent distinguishes ortho-dihydroxyl flavonoids by forming a copper complex between adjacent hydroxyl groups (SEIKEL, 1964.). This complex is dark on chromato- grams in ultraviolet light, while flavonoids lacking adjacent hydroxyl groups in the B-ring do not form the complex and appear as various shades of yellow t o yellow-green.
Compounds for visible and ultraviolet spectrophotometry were eluted from sections cut from chromatograms with spectroscopic grade methanol. Spectral measurements were made in methanol alone and in methanol containing each of the following reagents: (1) sodium acetate, (2) sodium methoxide, (3) aluminum chloride, (4) aluminum chloride-HC1-solution, and (5) sodium acetate-boric acid solution (JURD 1962 and MABRY, MARKHAM and THOMAS 1970). Acid hydrolyses of flavonoids were performed according to the procedures described by MABRY et al. 1970.
RESULTS
493
TABLE 1
Backcross segregation results x2 test for fit to a 1:l ratio for browning and nonbrowning silk response to wounding
Browning Nonbrowning
Backcross’ type type Total x 2 P
(NC236 x N C W 2 ) NC236 316 331 647 ,348 .5-.7
(Kys x T61) Kys 264 25 1 515 .328 .5-.7
* NC236 and Kys are nonbrowning types. T61 and NC232 are browning types.
(Kys and NC236) respectively, resulted in F, progenies of the browning pheno- type. When the Fl’s were backcrossed to the nonbrowning parent, a reasonable fit to a 1:l segregation ratio was obtained (Table 1). Backcrosses of the Fl’s to the browning parent resulted in only browning type progenies. These data indi- cated a monohybrid segregation with browning dominant to the nonbrowning trait.
The results of combining crude enzyme preparations free of phenolics (PVP treatment) and phenolic preparations free of enzymes (perchloric acid treat- ment) from the browning line T61 and the nonbrowning line Kys are presented in Table 2. The main diagcnal of Table 2 reconstitutes the original inbred lines and gave the predicted response. When the phenolic preparation from T61, a
browning type, was added to the enzyme preparation from Kys, a nonbrowning type, a typical brown pigment was formed. Therefore, Kys contained the en- zymes necessary for browning reaction. However, when phenolic preparations from Kys were combined with enzyme preparations from T61, no browning occurred. Clearly, Kys lacked the phenolic substrate necessary for brown pig- ment formation.
A rigorous study of the enzyme responsible for the browning response was not undertaken, but a preliminary characterization indicates it was polyphenol oxidase. Polyphenol oxidase oxidizes polyphenolic compounds in the presence of molecular oxygen forming quinones which condense with proteins or them- selves to f o r m the brown pigments. Crude enzyme preparation from corn silks rapidly catalyzed colored pigment formation with 0.1
%
solutions of the followingTABLE 2
Color reaction obtained b y mixing all combinations of enzyme and substrate from the silks of inbred lines T6l and Kys, a browning and nonbrowning type, respectively
Crude enzyme preparations free of phenolic compounds
T6 1 KYs
(browning type) (nonbrowning type)
T61
(browning type) brown brown
Substrate preparations free of enzymes
KY 5 yellow-green yellow-green
orthodihydroxyl compounds: catechol, chlorogenic acid, caffeic acid, L-dopa, and rutin. It catalyzed a slow color reaction with hydroquinone, a paradihy- droxyl compound, and no reaction with the meta-dihydroxyl compound, resorc- inol or the following monohydroxy1 compounds: phenol, m-cresol, guaiacol, and 2,4-dichlorophenol. Browning could be inhibited by strong reducing agents, abscorbate and L-cysteine, and the copper chelator, diethyldithiocarbamate.
In
the presence of diethyldithiocarbamate, the addition of a hydrogen acceptor, h.ydrogen peroxide, invoked browning probably due to peroxidase enzymes. Fifteen minute boiling of enzyme preparations was sufficient to destroy poly- phenol oxidase activity.
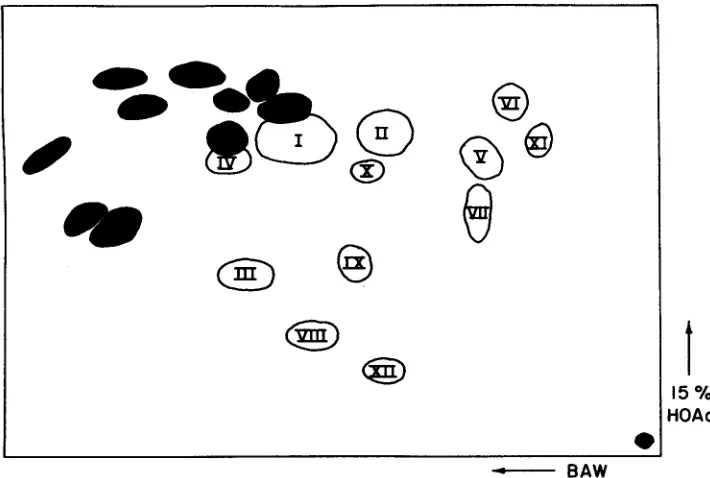
Two-dimensional paper chromatograms (BAW and HOAc) of silk extracts from inbreds T61, a browning type, and Kys, a nonbrowning type, revealed significant differences in constitution (Figure 1). Shaded spots on Figure 1 are those compounds which were common to both the browning and nonbrowning types, T61 and Kys, respectively. These compounds appeared as either blue or purple colored spots in ultraviolet light. Spraying with AlCl, solution altered color intensity and shade. Anthocyanin spots have been omitted since their presence or absence is independent of the browning response. Unshaded spots with Roman numeral designations are those compounds which are unique to the browning type. Compounds are numbered according to color intensity, com-
n
t
I5 08%
HOAc
-
BAWTABLE 3
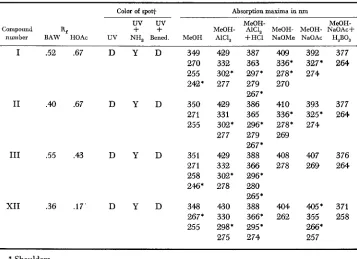
Diagnostic chromatographic attributes and absorption maxima for selected compounds present in the browning phenotype
Color of spot+ Absorption maxima in nm
uv
uv
MeOH- MeOH-Compound R,
+
+
MeOH- AlCI, MeOH- MeOH- NaOAc+ number BAW -HOAc UV NH, Bened. MeOH AICI, +HE1 NaOMe NaOAc H,BO,I .52 .67 D Y D 349 429 387 4.09 392 377
270 332 363 336' 327* 2644 255 302' 297' 278' 274 242' 277 279 270
267'
I1 .# .67 D Y D 350 429 386 410 393 377
271 331 365 336' 325' 2644
255 302' 296' 278' 274 277 279 269
267'
I11 .55 .43 D Y D 351 429 388 408 407 376
271 332 366 278 269 2644 258 302' 296*
2%' 278 280 265
'
XI1 .36 .17' D Y D 348 430 388 404 405* 371
267' 330 366' 262 355 258
255 298* 295' 266'
275 274 257
* Shoulders.
+
Bened = Benedict's solution; D = Dark; Y = Yellow.pound I being the most intense and compound XI1 the least. These twelve spots appeared dark in ultraviolet and yellow to yellow-brown in ultraviolet after spraying with AlC1,. With Benedict's reagent the twelve compounds were yellow in visible light and dark brown in ultraviolet. These results indicate the brown- ing type is distinguished by the presence of at least 12 ortho-dihydroxyflavones. Compound I, which occurred in the greatest quantity, was tested individually to determine if it would serve as a substrate for brown pigment €ormation. It was eluted from chromatograms, put into aqueous solution and mixed with a crude silk enzyme preparation free of polyphenolics. A brown pigment was formed which confirmed this compound as one of those contributing to silk browning in response to wounding.
Sufficient quantities of compounds I, 11, I11 and XI1 were isolated from chromatograms for visible and ultraviolet absorption spectrophotometry (Table
3 ) . Compounds I, 11, and I11 occur in large quantities in the silk of browning types while XI1 occurs in every small amounts. Compounds I, 11, and I11 gave similar spectral peaks. The double absorbance band at 270-271 (shoulder) and
255-258 nm and the single band at ca. 350 n m are characteristic of 5,7,3',4'-
droxyl groups in the B ring is indicated by the magnitudes of the hypsochromic shift in Band I with AlCl,/HCl relative to the AlC1, spectrum and the batho- chromic shift in Band
I
in NaOAc/H,BO, relative to the MeOH spectrum. The presence of a free 7-hydroxyl group in the A ring is demonstrated b y the batho- chromic shift in Band I1 in the presence of NaOAc. Spectra €or compound XI1 were similar to the spectra for compounds I, 11, and I11 in all solvents except the one containing NaOAc. No significant bathochromic shift was obtained with NaOAc which indicated the 7-position in the A ring was blocked. All spectral interpretations were based upon the results of JURD (1962) and MABRY et aZ.(1970).
All four compounds mere subjected to acid hydrolysis. However, insufficient quantities of aglycones were obtained from compounds I and I11 for identification. Hydrolysis of compound I1 yielded the C-glycosylflavones, orientin and isoorien- tin. Identification was based on cochromatography in two solvent systems (BAW and HOAc) with authenicated compounds and the demonstration of hydrolytic isomerization of the putative orientin and isoorientin. A milder hydrolysis of I1 gave a large amount of isoorientin and trace of orientin. This suggested that I1 is a derivative of isoorientin. Hydrolysis of XI1 produced luteolin which was identified by cochromatography in two solvent systems with authenicated luteolin. Based upon spectral, Rf and hydrolysis data, XI1 is idezltified as luteolin 7-0-glycoside.
DISCUSSION
Brown pigment formation in injured corn silks is due to enzymaiic oxidation of ortho-dihydroxyl compounds. At least 12 chromatographically different ortho- dihydroxyl compounds have been detected in silks from the lines which brown in response to wounding. The three compounds which occur most abundantly have been identified as luteolin derivatives. One, compound 11, is a C-glycosyl- flavone, an isoorientin derivative. To our knowledge, this is the first report of a C-glycosylflavone in corn. A fourth compound, XII, has been identified as luteolin 7-0-glycoside. Although the remsining 8 less abundant compounds have not been identified, it seems likely, based on chromatographic studies, that they will be found to be flavones. Clearly, ortho-dihydroxylflavones are the most important compounds in brown pigment formation in response to wounding. Silks from certain lines do not form a brown pigment when injured. Analyses indicate that these silks have polyphenol oxidase activity, but lack the ortho- dihydroxyl substrate necessary for pigment formation. None of the 12 ortho- dihydroxyl flavonoid compounds found in the browning phenotypes are present in the silks of nonbrowning types. One compound, chlorogenic acid, has been detected by cochromatography in browning and nonbrowning silks Although it
is an oxidizable ortho-dihydroxyl compound, i t occurs in insufficient quantities to contribute perceptibly to the brown pigment.
which results in browning of cut silks because of the presence of the 12 oxidizable o ~ h o - d i h y ~ o x y l compounds. Fu was chosen because the 3 compounds present in largest amounts have been identified as flavones. The recessive allele has been designated fu. I n the homozygous recessive condition, fufu, no browning of cut silks occurs because the 12 oxidizable compounds are absent. The dominant allele, Fu appears to behave as a completely dominant allele since a quantitative difference in brown pigment cannot be demonstrated between FvFu and Fufu
genotypes. The following commercial inbreds have been classified as FuFv geno- type: CI21, Hy2, L317, NC232, T61, T204, and WF9, while the Iollowing in- breds are of the genotype fvfu: Kys, NC34, NC45, and NC236.
Several points can be made concerning the block to flavonoid synthesis imposed by the fvfu genotype. First, the fufu genotype does not block all flavonoid syn- thesis since several inbred lines of the fufu genotype contain anthocyanin in their silks (e.g. NC236 and Jarvis line #52). Second, the fufu genotype does not block flavone synthesis in the leaves and tassel. Preliminary chromatographic studies of leaves and tassels from the fufu genotype showed the presence of flavones similar to those found in silks of the FvFu genotype. ~ o m p a ~ s o n s were based on Rr’s and color of compound in visible and ultraviolet light with AlC1, and Bene- dict’s reagent. Thus, the fvfu block to flavone synthesis appears limited to the silks. Finally, the fu locus may prove useful in studying flavone synthesis, par- ticularly if additional mutants in the pathway can be found.
The technical assistance of ME. PATSY BING is gratefully acknowledged.
SUMMARY
several fufu genotypes have anthocyanin in their silks, therefore, the fufu block
does not prevent all types of flavonoid synthesis. Second, the block to flavone synthesis in the fufu genotype appears confined to the silks since flavones are present in leaves and tassels of the fufu genotype.
LITERATURE CITED
ANDERSEN, R. A. and J. A. SOWERS, 1967 Optimum conditions f o r bonding of plant phenols to
insoluble polyvinylpyrrolidone. Phytochem. 7 : 293-301.
JURD, L., 1962 Spectral properties of flavonoid compounds. pp. 107-155. In: T h e chemistry O!
flauonoid compounds. Edited by T. A. GEISSMAN. Macmillan, New York.
LOOMIS, W. D. and J. BATTAILE, 1965 Plant phenolic compounds and the isolation of plant enzymes. Phytochem. 5 : 423-438.
MABRY, T. J., K. R. MARHHAM and M. B. THOMAS, 1970 T h e systematic identification of
flauonoids. Springer-Verlag, New York.
SEIKEL, M. K., 1962 Chromatographic methods of separation, isolation and identification of flavonoid compounds. pp. 34-69. In: T h e chemistry of flavonoid compounds. Edited by T. A. GEISSMAN. Macmillan, New York. -- , 1964 Isolation and identification of phenolic compounds in biological material. pp. 33-76. In: Biochemistry of phenolic componds. Edited by J. B. HARBOXNE. Academic Press, New York.
Electrons, defense and regulation. The 1969 W. 0. Atwater Memorial Lecture. U.S. Printing Office: 1970 0-375-692.