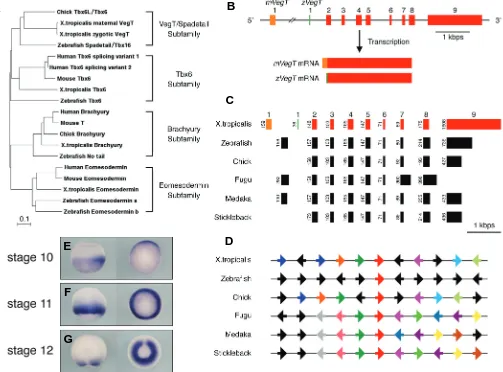
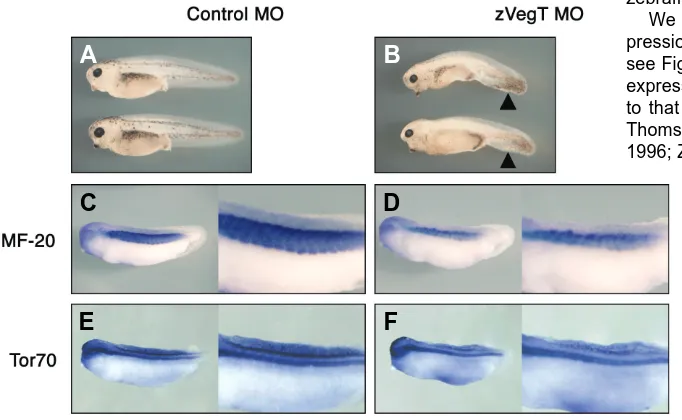
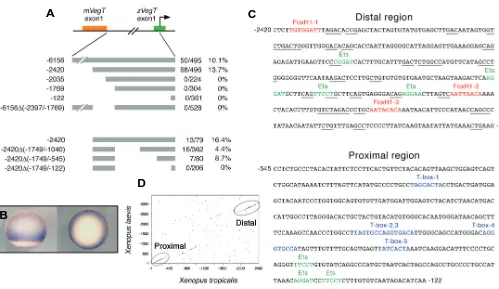
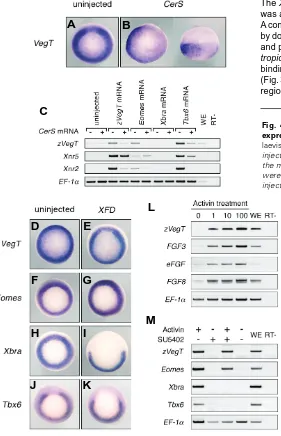
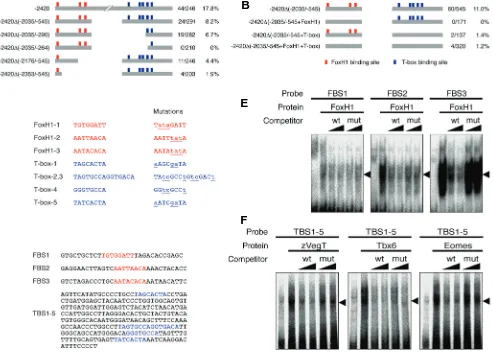
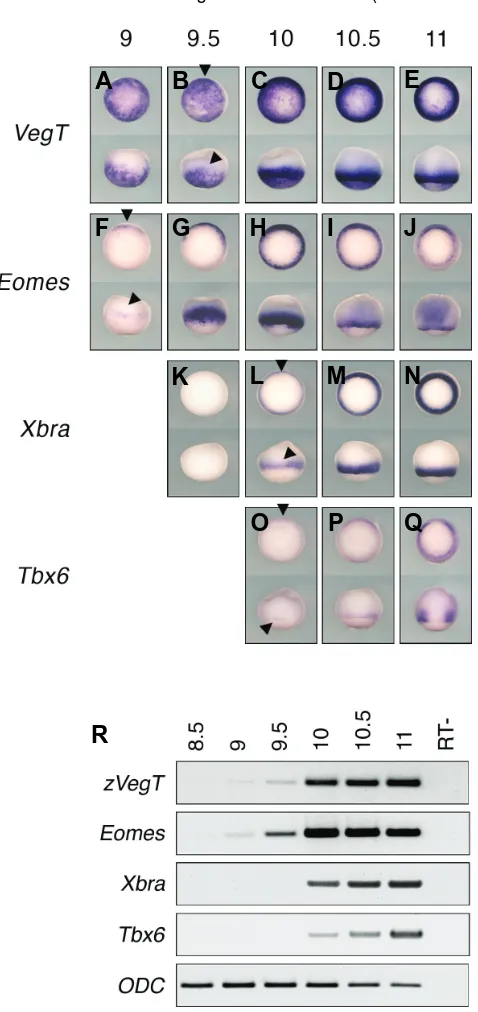
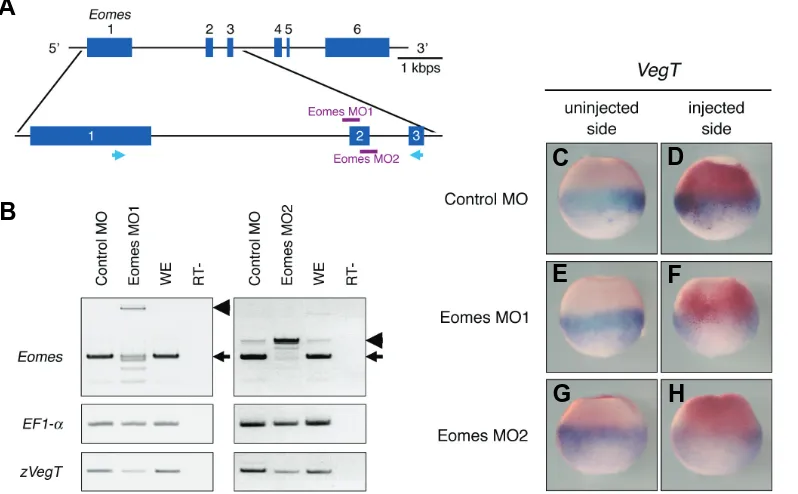
Zygotic VegT is required for Xenopus paraxial mesoderm formation and is regulated by Nodal signaling and Eomesodermin
Full text
Figure
Related documents
The growing diversity of the workforce on nineteenth century Brazilian and Cuban plantations _ estate slaves, leased slaves, emancipados, Asian, Mexican or Europeans employed
The interesting novelty which Gilbert added to the cosmological speculations of the time concerns his proposals of the magnetic nature of some of the forces
Based on the decision, the conclusion is that F arithmetic (18,203)> F table (3.09) then the halal label and price together have a positive and significant effect
Therefore, this study intends to investigate the phenomenon of how senior teachers are supported by the head teachers and explore strategies that are in place
The dam layout is situated across Bengoh River which is approximately 1.5 km upstream from the confluence of Sarawak Kiri River and Semadang River and majority of the
Second, a strand-specific library is necessary to prove that putative gRNA genes produce transcripts which are antisense relative to the protein gene transcript; however, the
In an effort to provide more certainty to foreign limited partnerships in light of the potentially harsh consequences resulting from failure to qualify, TRLPA
Inspired by the shape and function of nature, some architects have created great architectures by finding and applying new design sources from nature’s elements.. The beautiful
