royalsocietypublishing.org/journal/rsif
Research
Cite this article:
Sedda L
et al
. 2019
Improved spatial ecological sampling using
open data and standardization: an example
from malaria mosquito surveillance.
J. R. Soc.
Interface
16
: 20180941.
http://dx.doi.org/10.1098/rsif.2018.0941
Received: 13 December 2018
Accepted: 19 March 2019
Subject Category:
Life Sciences – Earth Science interface
Subject Areas:
biogeography, biomathematics, environmental
science
Keywords:
remote sensing and field data, mosquito
sampling, stratification, adaptive and
non-adaptive sampling design, model-based
geostatistics, Sub-Saharan Africa
Authors for correspondence:
Luigi Sedda
e-mail: [email protected]
Martin J. Donnelly
e-mail: [email protected]
Electronic supplementary material is available
online at http://dx.doi.org/10.6084/m9.
figshare.c.4447709.
Improved spatial ecological sampling
using open data and standardization:
an example from malaria mosquito
surveillance
Luigi Sedda
1, Eric R. Lucas
2, Luc S. Djogbe´nou
2,3, Ako V. C. Edi
4,
Alexander Egyir-Yawson
5, Bilali I. Kabula
6, Janet Midega
7, Eric Ochomo
8,
David Weetman
2and Martin J. Donnelly
2,91Centre for Health Information, Computation and Statistics (CHICAS), Lancaster Medical School, Lancaster
University, Furness Building, Lancaster LA1 4YG, UK
2Department of Vector Biology, Liverpool School of Tropical Medicine, Pembroke Place, Liverpool L3 5QA, UK 3Institut Re´gional de Sante´ Publique/Universite´ d’Abomey – Calavi, BP 384 Ouidah, Benin
4Centre Suisse de Recherches Scientifiques en Cote d’Ivoire, 01 BP 1303 Abidjan 01, Cote d’Ivoire 5Department of Biomedical Sciences, University of Cape Coast, Cape Coast, Ghana
6National Institute for Medical Research (NIMR), Amani Centre, PO Box 81, Muheza, Tanzania
7Centre for Geographic Medicine Research, Kenya Medical Research Institute, PO Box 230, 80108 Kilifi, Kenya 8Centre for Global Health Research, Kenya Medical Research Institute, PO Box 1578 – 40100 Kisumu, Kenya 9Wellcome Sanger Institute, Hinxton, Cambridge CB10 1SA, UK
LS, 0000-0002-9271-6596; ERL, 0000-0003-3892-1668; LSD, 0000-0002-7381-6321; AVCE, 0000-0003-1564-4354; BIK, 0000-0002-7374-169X; EO, 0000-0002-4042-4970; DW, 0000-0002-5820-1388; MJD, 0000-0001-5218-1497
Vector-borne disease control relies on efficient vector surveillance, mostly carried out using traps whose number and locations are often determined by expert opinion rather than a rigorous quantitative sampling design. In this work we propose a framework for ecological sampling design which in its preliminary stages can take into account environmental conditions obtained from open data (i.e. remote sensing and meteorological stations) not necessarily designed for ecological analysis. These environmental data are used to delimit the area into ecologically homogeneous strata. By employing Bayesian statistics within a model-based sampling design, the traps are deployed among the strata using a mixture of random and grid locations which allows balancing predictions and model-fitting accuracies. Sample sizes and the effect of ecological strata on sample sizes are estimated from previous mosquito sampling campaigns open data. Notably, we found that a configuration of 30 locations with four households each (120 samples) will have a similar accuracy in the predictions of mosquito abundance as 200 random samples. In addition, we show that random sampling indepen-dently from ecological strata, produces biased estimates of the mosquito abundance. Finally, we propose standardizing reporting of sampling designs to allow transparency and repetition/re-use in subsequent sampling campaigns.
1. Introduction
Sampling design is a crucial step in any survey as it affects the quality of data col-lection and analysis [1]. Sampling strategies should therefore be designed to maximize the effectiveness of the study, using any relevant preliminary and back-ground data available [2]. Furthermore, because published sampling strategies frequently inspire designs for future studies, both the design details and justifica-tion should be rigorously reported. Despite improvement in recent years, both the
&
2019 The Authors. Published by the Royal Society under the terms of the Creative Commons Attributionuse of available informative data and the rigour with which sampling designs are reported continue to fall short of what could be achieved [3]. Specifically, the amount of environ-mental data available from open-data platforms is often acknowledged but rarely exploited to support sampling design, while the necessary information for study repeatability, comparability or usability are often inadequately reported. Such data, even when not collected for ecological analyses, can support representativeness in investigations of population dynamics, epidemiological processes and biological studies. Here we use an example from malaria vector surveillance to design a sampling strategy for collecting mosquitoes for whole genome sequencing based monitoring and evaluation.
Genomic technologies are radically transforming our understanding of vector-borne disease transmission dynamics [4] due to the capacity to unveil complex interaction between human, pathogen, vector and environment. Whole genome sequencing projects have revealed novel genetic loci associated with increased susceptibility to malaria in the human host [5,6] and made major contributions to our understanding of how anti-malarial and insecticide resistance evolves [6]. However, the impact of environment on genotype distri-butions is much more poorly understood, reflecting at least in part the use of insufficiently ecologically informed sampling strategies. Much of the sampling conducted in vector surveil-lance studies is opportunistic and lacks a rigorous sampling framework. Often, ecological and entomological sampling designs rely solely on resource availability rather than aiming to maximize representativeness and precision of the variable of interest, e.g. collectors target locations where disease vectors are known to be abundant.
Designing a field sampling strategy requires three decisions: what is the variable of interest (formally the estimator, e.g. vector density), the sampling approach (e.g. model-based or not) and sampling location distribution (e.g. the number and spatial / temporal allocation of sampling points). These decisions constitute the sampling strategy trinity [7] in which each element strictly depends on the other two. Sampling strategies are further complicated by deterministic (e.g. due to age, environment, socio-economic, etc.) and stochastic (i.e. spatio-temporal autocorre-lation) factors. Our literature search in Web of Science on spatial sampling of mosquitoes (search terms: mosquito OR anopheles AND sampling AND spatial, in title/keywords/ abstract—last access in August 2018) shows that while all studies provide a general description of the sampling design, only a limited number of papers (i.e. [8–11]) give a detailed description of the rationale, decisions and calcu-lations related to the ‘Where, When, How and How many’ samples to collect (see for example the reviews from [12] and [13]). In other literature, partial justification of the sampling design is provided. For example, [14– 22] used pre-vious surveillance information and remote sensing data to identify potential mosquito habitat types (or, in statistical terms, ‘strata’). However, the method used or assumptions made to choose the within-strata location and number of traps were not described, perhaps because these were entirely guided by practical considerations (i.e. [23]) or due to the high level of complexity or uncertainty in the scope of the sampling, which makes quantitatively-driven spatial sampling design very difficult (for example when the scope is describing a concurrent variety of species such as Ano-pheles, CulexandAedes) [24,25]. Conversely, descriptions of
sampling over time are often provided in detail, with explicit information on the frequency and length of the sampling campaign.
The picture that emerges from the literature is that using habitat stratification to inform sampling is a common pro-cedure in vector biology, but often based on subjective or qualitative decisions. However, stratification has a fundamen-tal role in describing and reducing the error in estimates of mosquito variation, which in turn influences surveillance success, assessment of epidemiological risk and genetic diversity [26]. Identifying a set of (independent) environ-mental variables homogeneous within strata allows a better representation and representativeness of the environment related to the property or properties under study (i.e. insect abundance and insecticide resistance) [13]. Unless the spatial or spatio-temporal autocorrelation of the property under study is tested and found negligible [27], these approaches often incorrectly assume independence between samples in space and time [28], which is an unrealistic assumption for most of the ecological processes. Spatial and spatio-temporal heterogeneity can be accounted for in sampling design by adopting a geostatistical model-based sampling design [8,29]. Ecological stratification of sampling designs is now facili-tated by web-based open data providers, allowing rapid access to large amounts of information on climate and land-use, which are commonly associated with biogeographic patterns of human and animal health and species distribution [30]. This availability of open data (largely remote sensing) for almost every global location, combined with appropriate spatio-temporal algorithms [15], make quantitative ecological stratification more accessible as a preliminary step to any sampling programme. Nevertheless ‘very few studies propose, at an early phase of research work, objective sampling strategies that are consistent with both study goals and constraints’ [13]. In this work we propose a framework for optimizing the sampling design of the spatial distribution of mosquito populations using open data, which we hope will be relevant to a wide range of ecological, disease monitoring and genomic studies.
This paper describes the stages constituting our sampling framework which is based on the following decisions:
1. The variable of interest. In our case is the presence of insecticide resistance genes in the mosquito genomes. To achieve this we will trap mosquitoes in areas with known or suspected insecticide resistance.
2. The sampling approach. The sample size is calculated based on previous mosquito surveys, and sample locations are defined to balance prediction and parametrization, i.e. the accuracy in predictions and the goodness of model fitting when limited amount of information of the variable of interest is available.
3. The stratification. The open data are used to ecologically characterize the area(s) under study and inform the location of each trap. The effect of ecological strata on sampling size is estimated from a previous malaria control surveillance campaign.
Finally, we discuss the necessity and benefits of a stan-dardization of the sampling design procedures and reports to make them repeatable and reusable.
2. Materials
2.1. GAARDian project
The sampling design described in this work has been developed within the UK-MRC-funded GAARDian project (https://www.anophelesgenomics.org/gaardian). The main objective of this project is to investigate the spatial and temporal scale of variation in mosquito gen-omes to improve our understanding of the processes underlying the spread of insecticide resistance. Insecticide resistance is a major threat to the sustained control of malaria, as 260 million averted clinical cases of malaria have been due to the use of insecticides that target the mosquito vector [31].
2.2. Study area
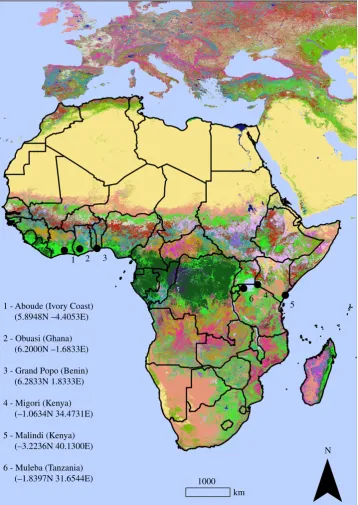
Six sites were chosen based on suspected use of insecticide or presence of insecticide resistance (figure 1) (see electronic supplementary material, appendix A, for detailed description of each site). Around each site, an operable area was deter-mined as the largest area where traps can be deployed and routinely checked by two operators. The operable area was a 6060 km square centred on the site.
2.3. Environmental data
We used open data information from several sources to stratify the ecological variations of each study site. These data include land cover, climate and topography [32]. 1 - Aboude (Ivory Coast)
(5.8948N –4.4053E)
2 - Obuasi (Ghana) (6.2000N –1.6833E)
3 - Grand Popo (Benin) (6.2833N 1.8333E)
4 - Migori (Kenya) (–1.0634N 34.4731E)
5 - Malindi (Kenya) (–3.2236N 40.1300E)
6 - Muleba (Tanzania) (–1.8397N 31.6544E)
N
1000
1 2 3
4 5 6
km
Figure 1.
Location of the GAARDian sampling sites, shown on a land cover background (GlobeLand30 land covers). Map was made using ArcMap 10.4 (http://
desktop.arcgis.com/en/arcmap/). Source administrative limits: http://www.maplibrary.org/library/index.htm. (Online version in colour.)
ro
yalsocietypublishing.org/journal/rsif
J.
R.
Soc.
Interfa
ce
16
:
20180941
GlobeLand30 (http://www.globallandcover.com/ GLC30Download/index.aspx) is a global land cover map of 30 m resolution produced by the National Geomatics Center of China and containing 10 land cover classes (full description of classes in [33]). The images used for GlobeLand30 classification are multispectral images, including the TM5 and ETMþ of America Land Resources Satellite (Landsat) and the multispectral images of China Environmental Disaster Alleviation Satellite (HJ-1). Globe-Land30 raster adopts WGS84 coordinate system, UTM projection, 6-degree zoning and the reference ellipsoid is WGS 84 ellipsoid.
The moderate-resolution imaging spectroradiometer (MODIS) satellite products are provided in monthly time-series at 0.05 degree (approx. 5 km) resolution from observations by the MODIS sensor on Terra (AM) for the period February 2000 to December 2013 inclusive and available at (https://ora.ox.ac.uk/objects/uuid:896bf37f-a56b-4bc0-9595-8c9201161973) [34]. The following MODIS products were used:
— MODIS Enhanced Vegetation Index (EVI) from the MOD13C2 product comprises monthly, global EVI. This resource provides consistent spatial and temporal compari-sons of vegetation canopy greenness, a composite property of leaf area, quantity of chlorophyll and canopy structure. EVI improves sensitivity over dense vegetation conditions or heterogeneous landscapes when compared to Normal-ized Difference Vegetation Index (NDVI).
— MODIS Air Temperature (Temp) from the MOD07_L2 Atmospheric Profile product comprises monthly, global temperature at the closest level to the earth’s surface. — MODIS Evapotranspiration (ET) from the MOD16 Global
Evapotranspiration product is calculated monthly as the ratio of Actual to Potential Evapotranspiration (AET/PET).
Precipitation was obtained from WorldClim v. 2 as average annual precipitation from 1970 to 2000 at 30 arcseconds (1 km2ca) (http://worldclim.org/version2) [35]. Finally, elevation was obtained from the NASA Shuttle Radar Topographic Mission (SRTM) 90 m Digital Elevation Database v. 4.1. The SRTM 90 m DEM’s have a resolution of 90 m at the equator. The DEM is available in geographical coordinate system—WGS84 datum (https://drive.google. com/drive/folders/0B_J08t5spvd8VWJPbTB3anNHamc).
3. Methods
Defining a standard spatial sampling design does not affect the multitude of choices necessary for each different problem, but it requires that three elements are fully described: sample size; stratification (if stratification is performed) and geographical allocation of the sampling points. A description of each element optimization is given below.
3.1. Sampling size optimization
For one of the collection areas (Migori, Kenya, location 4 in figure 1), additional data were available from entomological sur-veillance carried out from December 2015 to September 2017 as part of indoor residual spraying (IRS) (Abong’o et al. 2018, unpublished; http://www.africairs.net/about-airs/), which we will refer to as AIRS data hereafter. As in the GAARDian project, collections were made using CDC light traps [36], hung in each
house over the sleeping area, approximately 1.5 m from the ground, adjacent to an occupied bed net. The traps were run from 18.00 and mosquitoes were collected at 07.00 the next morn-ing. Placing the trap near sleeping space facilitates sampling female mosquitoes that are actively seeking a blood meal. We used this preliminary information about mosquito abundance to estimate the optimal sample size (in terms of mosquito distribution) to be used in all sites.
From the AIRS data, we first estimated the spatial covariance function (via maximum-likelihood estimation, [37]) that was used to simulate a log Gaussian Cox process (LGCP) [38] mimicking the mosquito spatial distribution process found in Migori. This can be translated in lay words as a process (mosquito catches) that is environmentally driven but producing values of catches that can be considered independent (i.e. catch on one occasion does not predict subsequent catches in the same or nearby locations) although the average process is spatially dependent (hence the necessity to estimate the spatial covariance function above).
The Gaussian random field is of the form [39]
y(l)¼m(l)þZ(l)þ1, ð3:1Þ
wherelis the location,mis the mean,Zis the Gaussian process with Matern correlation function, and1is the error term (noise or nugget).
The Matern correlation function has the general form
Z(l)¼ 1
whereKh(.) is the modified Bessel function of orderhandris the
spatial range [40]. Bothhand rmust be positive and different from 0.
Finally the Poisson LGCP can be written as [41]
Y(l)P(l(l)) ð3:3Þ
and
l(l)¼exp(y(l)), ð3:4Þ
where Y is the mosquito density point process and l is the conditional mean. As can be easily noted, equation (3.4) links directly to equation (3.1).
From the LGCP we predicted the estimated variance in the parameters of the spatial covariance function and the prediction error for a set of sample sizes (15, 30, 75, 150, 200 and 300) assumed randomly allocated in the area of Migori.
This will allow the allocation of the (limited) resources to obtain the sample size that will produce the desired prediction error and variance in the spatial covariance parameters (if this is an objective of the sampling design).
3.2. Stratification (ecological delineation)
In many areas of physical, engineering, life and social sciences, inferential and predictive classification are prevalent tools to discriminate between classes and to interpret the differences. Examples range from identification of ecological niches to brain and bone anomalies. While the growing amount of open access information enables discrimination among a large number of ecological classes, many traditional algorithms fail for these data because of decreased classification performance (leading to overfitting) and mathematical/practical limitations [42]. One method is to describe ecological strata in terms of transformed environmental variation (i.e. factorial analyses) [13], but the results can be difficult to interpret. By contrast, discriminant analysis (DA) requires less computational time and resources because no parameter tuning is required [43]. Discriminant analysis [44] is a common multivariate statistical approach for data classification (for example, in 2017 2026 scientific articles
were published on the use or improvement of discriminant analysis—search terms used in Web of Science: ‘discriminant analysis’, in title/keywords/abstract).
The simplest forms of DA are linear (LDA) and quadratic (QDA). LDA can be seen as a regression line whose orientation divides a high-dimensional space, reducing the dimensionality while keeping each class separate from the other classes. In practice, the optimal orientation is the one that minimizes the within-class variance and maximizes the between-class variance [45]. The main assumption of LDA is that all the classes have a common variance –covariance matrix, i.e. the relationships between classes and explanatory variables are independent from class membership, while the differences between classes are dependent only on the mean.
When the variance –covariance matrices are not homo-geneous for two or more classes, linear discriminant analysis cannot be applied. Instead the QDA can be employed. The QDA discriminant function is
fiQ¼ 1
whereX is the matrix of variables,mthe vector containing the mean of each variable and S is the variance –covariance matrix, andpithe ‘prior’ probability of each point to belong to
the classi.iis the subscript for classi, withi¼1,. . .,Nwhere
Nis the total number of classes.
fis calculated based on a training dataset (class memberships are known). The larger the f value, the higher the probability that the point belongs to that group. For a training dataset, the
pican be calculated in several ways, usually by ‘equal priors’
method: each class has a prior probability equal to 1/N. In this analysis, and in order to take into account the spatial proxi-mity of the classes, a local frequency prior method was used. It estimates the classpiprior probability as the relative frequency
of i labels in the neighbourhood. Similarly, predicting a label for a new point means looking at the local proportion of each class (as classified from the training dataset) around the new point.
Oncef is maximized with the training dataset, a new data point can be classified by calculatingf for the new point and for each class (equivalent to calculating the position of a point with respect to all available class centroids), and assigning to it the class index at which corresponds the maximumf.
The QDA has been embedded into an algorithm that determines the optimal number of ecological classes and their geographical delimitation for each area (see electronic supplementary material, appendix B, for further details).
The analysis was carried out taking all the environmental variables at their original spatial resolution, and providing the output (classification) at 30 m resolution (the same as the land cover resolution).
3.3. Spatial allocation of the sample households
For the present study the malaria vector species we are targeting, within theAnopheles gambiaespecies complex, are usually highly anthropophilic and commonly found in houses. Therefore, the traps are located inside households.
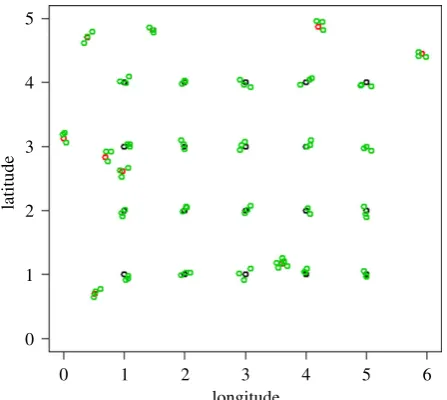
Locations of the sampling points, in each sampling site (figure 1), follow a ‘lattice plus close-pairs’ design [46] which combines regular lattice (efficient for predictions) and random points as close pairs (efficient for parameter estimation) [47].
For an easier understanding of the sampling design, we refer to the six locations distributed in West and East Africa as sampling sites (shown in figure 1). Each sampling site will con-tain M sampling points. Each sampling point contains V
households. Therefore, the total number of households sampled per sampling site isMV.
For all sites except Migori, the distribution of Msampling points are realized under two conditions: (i) 70% of sampling points are in grid (lattice) and 30% close-pairs randomly allocated (a proportion usually applied in simulation analyses, i.e. [46] and [48]) (figure 2); (ii) each stratum must contain a number of points proportional to the stratum size [49]:
ni¼M
Ai
AT
, ð3:6Þ
whereniis the number of points for classi;Aithe area of class
i;and ATis the total area. The term ‘close pairs’ here is used
loosely, since not all the points in the grid will have a close pair, and some close pairs may be shared between points.
In Migori alone (where a previous sampling campaign, AIRS, took place) an adaptive sampling design was trialled in which AIRS sampling served to inform the location of theMsampling points. From the LGCP model (see above), we estimated the predic-tion variances at each grid cell, and attributed theMlocations to the cells with highest prediction variance (uncertainty) [29,50–52].
3.4. Effect of stratification on sample size and
improvement of mosquito abundance models
In order to evaluate the effect of stratification:
(a) on the ratio between the Poisson rate parameter of mosquito counts from a survey (l1) and the Poisson rate parameter of
mosquito count from a sub-sample of it (l2) (random
versus stratified);
(b) and on the goodness of fitting of mosquito abundance models;
we have considered a mosquito sampling campaign from Uganda. This data is from 104 health subdistricts (HSD) where estimates of A. gambiae and Anopheles funestus densities (as determined by a standard collection method) for both male and female mosquitos are available. The sampling design was based on a cluster randomized trial with 10 houses selected at
longitude
random from each HSD, and mosquito collection was made every six months for 2 years (http://www.isrctn.com/ ISRCTN17516395).
For objective (a) we have employed a Poisson exact text on the null hypothesis that the ratio between l1 (obtained from
the entire Uganda mosquito collection data) andl2 (obtained
from a sub-sample of mosquito collections of the Uganda data) is equal to 1, i.e. the two conditional means are not different [53]. The test is performed by first randomly sampling 2, 3, 4 and 5 mosquito collection locations from each strata. For each of these sample sizes thel2is calculated and the test performed.
The process has been repeated 999 times, to randomize the location selection, and 95% confidence interval from the all bootstrapping are estimated. The procedure above was then com-pared with a sampling design that randomly extracts the same amount of locations from the entire dataset but independently from the strata to which they belong.
For objective (b) we fitted the total number of mosquitoes for each species and at each location (over the two years of collec-tion) using the ecological strata produced by performing the same methodology described in the above section ‘Stratification’. The model fitting employs a Poisson generalized linear model [53] and model comparison against the null model is performed using a MANOVA test [18].
4. Results
4.1. Sample size
In order to estimate the impact of sample size on model fitting and predictions, we simulated a log Gaussian Cox Process with known covariance function. The latter has been parametrized with the AIRS mosquito surveillance data via maximum-likelihood estimation. The obtained LGCP parameters were: intercept of 21.77, spatial variance (sill—i.e. the amount of variance dependent on distance) of 14 478, spatial range of 16 km (i.e. the maximum distance at which variance increases with distance) and 0 nugget (variance independent of distance that can be due to measurement errors or un-explained factors). Therefore, according to the model, all the variation is considered to be spatially-dependent up to a 16 km range. Finally, the Matern kappa parameter (shape parameter) was 1.5 (see electronic supplementary material, appendix C).
The simulation results of the above model with different sample size are reported in table 1. With 30 sampling locations, the prediction error and the total variance in the LGCP parameters was halved compared to 15 locations, and 20 times less when using 75 locations. The standard error in predictions is the maximum number of mosquitoes predicted in excess or in deficiency to the true mean. Therefore with 30 traps it is estimated a maximum error of 93 mosquitoes around the real mean and with 200 locations an error of 22 mosquitoes.
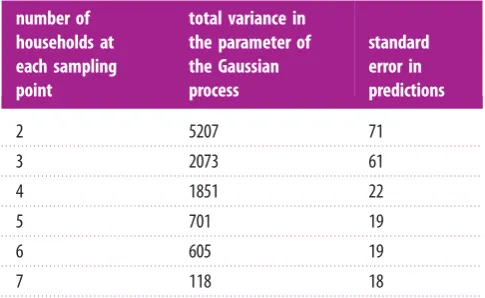
In order to improve local estimates, more than one house-hold within 2.5 km from each sampling point can be employed. Therefore if we take two households for each of the 30 sampling points, the total number of households is 60. The effect of the use of more than one household in model fitting and prediction is shown in table 2.
With four households and 30 sampling points we expect the same prediction error as using 200 random sampling points distributed across the entire area and each containing a single household (comparison of standard errors in
tables 1 and 2) but higher variance in the parameters. Using between five and seven households has little impact on the standard error in the predictions, although there is a significant improvement in the model fitting as the number of households increases (see total variance column in table 2). Consequently, the sampling design was chosen with 30 locations and four households which was considered a good balance in terms of standard errors, model fitting and economic feasibility.
4.2. Ecological classification
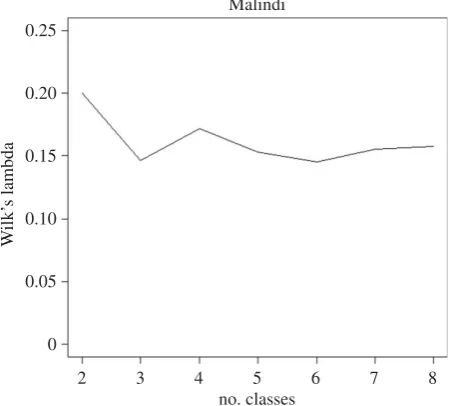
The stratification identified two ecological classes for Migori, Obuasi, Muleba and Aboude; three ecological classes in Malindi and four in Grand Popo. The Wilk’s criterion, measured as Wilk’s Lambda, for Malindi is shown in figure 3; for other sites, they are provided in the electronic supplementary material (appendix D). The biggest improve-ment (i.e. largest decrease of the Wilk’s Lambda) is in the change from 2 to 3 classes (figure 3).
A hierarchical numerical classification of the sites and classes is shown in figure 4. This dendrogram was obtained from the agglomerative method (classes are aggregated into progressively larger groups) group average [54]. The latter accounts for the average distances or similarities between all the members of the new group and those of the others.
Table 1.
Total variance in the parameters of the Gaussian process
(intercept, sill, nugget, range) and standard errors for the predictions at
different sample sizes.
sample
size
total variance in the
parameter of the
The heights in figure 4 represent the dissimilarities between classes (classes full description reported in electronic supplementary material, appendix E), which are very small for some intra-location comparisons (mg10 and mg55 in Migori (Kenya); and gp10 and gp35 in Grand Popo (Benin)) and indeed some inter-country comparisons (ob20 in Obuasi (Ghana) with ab85 in Aboude (Cote d’Ivoire)). The classification suggests geographical homogeneity for most of the sites, since close locations looks more similar (Migori and Muleba, Obuasi and Aboude) than same classes far apart. For example, Malindi and Grand Popo classes (with exception of class 95 in Grand Popo) form their own clusters. Both Grand Popo and Malindi are coastal sampling locations, albeit on opposite sides of the African continent.
Figure 5 shows the ecological classification and its uncertainty for the Malindi area. The same maps for the rest of the sites are shown in the electronic supplementary material (appendix F).
4.3. Sample locations
In figure 5 the sampling locations for Malindi are shown overlaid on the ecological classes. These locations are obtained from the lattice with close pairs sampling design, with a batch of four households (not shown in figure 5) at each sampling point. In this lattice with close pairs sampling design, 20 of the 30 locations are deployed in a 45 regular grid, and the rest allocated randomly as described in the methods. This general objective was modified to weight the number of locations by ecological classes; thus some of the points in the grid may have been adjusted slightly to be contained in the new class. In all the sites, each of these 30 locations constitutes a cluster of four households as in figure 2.
The sampling location in Migori followed an adaptive sampling design, in which only class 10 was sampled (see electronic supplementary material, appendix F) because of the constraint that previous samples (AIRS) only targeted this class. The allocation of 30 samples and households in class 10 in Migori were based on the prediction variance, i.e. new sampling points are allocated at the centre of 30 pixels with largest prediction variance [52] (figure 6).
4.4. Effect of stratification on sample size and
improvement of mosquito abundance models
For this objective we have used the Uganda dataset (see methods), which contains a large mosquito sampling cam-paign (104 districts, 1040 households) carried out over for 2 years. The first step was to identify the ecological strata for each cluster (of households) location. By applying the same stratification method described for here (see methods) we identified four ecological zones.The stratification was first used to evaluate if a sub-sample (up to 20% of the full dataset) of the mosquito collections, extracted from each strata, is still representative of the full mosquito collection obtained in Uganda. Stratifica-tion produced ratios that are not significantly different from 1 (i.e.l1andl2are not significantly different) for any sample size (see electronic supplementary material, appendix G, table G1). In contrast for random sampling all ratios between the two rate parameters are significantly higher than 1 for all the sample sizes (i.e. l1 and l2 are significantly different). Finally stratification improves the model fitting of mosquito counts when compared to a null model (see electronic sup-plementary material, appendix G, table G2).
5. Discussion
Vector-borne disease control and monitoring rely on vector surveillance, mostly carried out using trap-based indices and, more recently, remote sensing data [13,26]. Trap-based indices (density, population changes, distribution, etc.) are cal-culated from mosquito catches and require a system of traps dispersed in the field in sufficient numbers to represent mos-quito population ecology and dynamics. Conversely, remote sensing data can be used to define the ecological level of dis-ease risk based on mosquito ecological suitability [49]. This is a cheaper and quicker option, but may not have the spatial and temporal resolution necessary for practical interventions [55]. In a sampling design, trap-based indices and remote sen-sing analyses must be seen as complementary tools, since
no. classes Malindi
5 6 7
2 0 0.05 0.10
W
ilk’
s lambda
0.15 0.20 0.25
3 4 8
Figure 3.
Wilk’s Lambda criterion for Malindi. Graph was made using R-cran
3.5.0 (http://r-project.org).
14
12
10
8
6
height
4
2
mg 10 mg 55
mu 25
mu 60
ma 20 ma 45 ma 75 gp 95
ob 20 ab 85
ab 15
ob 65
gp 10 gp 35
gp 20
site’s ecological classes
Figure 4.
Dendrogram of agglomerative hierarchical clustering of the
ecologi-cal zones. mg, Migori; mu, Muleba; ma, Malindi; gp, Grand Popo; ob, Obuasi;
and ab, Aboude. For the description of the class number see electronic
sup-plementary material, appendix E. Graph was made using R-cran 3.5.0 (http://
r-project.org).
ro
yalsocietypublishing.org/journal/rsif
J.
R.
Soc.
Interfa
ce
16
:
20180941
remote sensing data contain the information necessary to define location and distance between sampling points.
Sample size, location, estimator and strategy (i.e. model based or not, adaptive or not) are the fundamental character-istics of a study design [27], which affect the likely success of describing the studied process (e.g. disease or organism distribution and abundance), its stochasticity, and, conse-quently, the accuracy of the estimates.
Ecologists are now equipped with algorithms, open infor-mation and datasets that enable a better understanding of the biology and spatial distribution of populations, which allows
optimization of collection site placement to best describe natural processes. Ecological/environmental classification is now possible for every region in the world [55]. Failure to exploit these data in ecological and genomic sampling frameworks ignores the spatial variability of favourable, unfavourable or neutral habitats, therefore random or trans-ect sampling designs may or may not be representative of the ground conditions and characteristics [56]. Even a grid design can be biased towards larger ecological classes and may miss linear features (i.e. a river passing between collection points) [1]. The consequence of which could be an over- or under-estimate of the true abundance, even when the population phenology is correctly delineated [56]. In some cases, however, a quantitatively determined sampling design may not be necessary, especially if the study intends to survey the entire area under study (as for example in [57,58]), or if it is a consequence of interventions recommended by national or international authorities [58,59] (i.e. WHO guidelines [60]).
In the illustrative example presented we demonstrate how these approaches may be used to develop an a priori
sampling strategy to sample malaria vectors for genomic and ecological studies. The ecological classification presented for each site returned a maximum uncertainty ranging from 0.37 to 0.44 depending on the site (figure 5 for Malindi and electronic supplementary material, appendix F, for the other sites), which can be interpreted as the probability that a grid node belongs to a different class. This level of uncertainty shows that classification identified dominant classes. In addition, the ecological classification also shows that areas (the six sampling sites) with putatively the same land cover are still ecologically different when consider-ing the full set of environmental variables (temperature, precipitation, elevation, evapo-transpiration and vegetation), sampling points
countries Africa
20 75 45
Malindi (East Kenya) N uncertainty
classes high: 0.5
low: 0
km 10
Figure 5.
Ecological classification and uncertainty for the area of Malindi. Map was made using ArcMap 10.4 (http://desktop.arcgis.com/en/arcmap/). Source
administrative limits: http://www.maplibrary.org/library/index.htm. (Online version in colour.)
–1.2 –1.1 –1.0 –0.9 –0.8
longitude
latitude
34.2 34.3 34.4 34.5 34.6
10 000 20 000 30 000 40 000 prediction variance
Figure 6.
Adaptive sampling for Migori in class 10. Black dots are the AIRS
mosquito surveillance locations. The grey dots (blue on the online version)
are the adaptive locations, which are targeting the cells with largest
predic-tion variance. Graph was made using R-cran 3.5.0 (http://r-project.org).
(Online version in colour.)
ro
yalsocietypublishing.org/journal/rsif
J.
R.
Soc.
Interfa
ce
16
:
20180941
and that geographical proximity is a dominant factor in ecological clustering [61] (figure 4). It is therefore not surprising that sites cluster much more strongly within country than within ecotype; e.g. forest (class 20) in Malindi (Kenya) is not equivalent to forest in Obuasi (Ghana) or Grand Popo (Benin). The ecological classes, while not often used in modelling mosquito populations and commu-nities for medium- and large-scale analyses, represent the complex interaction of environmental and socio-economic conditions [23].
Another factor that we accounted for during our sampling design is the spatial autocorrelation of mosquito catches (model-based sampling design) [12]. The effect of strong autocorrelation can reduce the overall statistical power (and the overall biological significance of the study) as it results in effectively a lower sample size (because the assumption of independence is violated), underestimates of variance, and increases in type I error [10]. Geostatistical approaches, such as the one applied here, can lead to unbiased estimates of population parameters and avoid the risks and limitations of random, or haphazard, selection of sampling locations. Given the requirements to satisfy both parametrization and predictions [47], the simulated inhibitory design adapted from [50] in order to contain clusters of households at each sampling point, has shown that with 120 sampling houses for each site distributed across 30 sampling points, we achieve the same prediction error (main goal) as from 200 points allocated at random, albeit at the expense of parameter accuracy. However, there were important limitations in the sample size/location calculation. Firstly, they are based on limited pre-existing mosquito surveillance data from Migori, which may not describe the different spatial scales of the mosquito abun-dance distribution [26]. This is a concern due to the large variation in abundance levels observed throughout the period, but that can be solved by deploying an adaptive sampling design, i.e. concentrating the new samples where we have the largest uncertainties (our knowledge is poor) in the process of interest (abundance or a level of abundance). In addition, we are assuming that the mosquito population dynamics in Migori are similar to those in the other sites. The ecological classification has the advantage of correcting for local mosquito population dynamics although this is not a full solution. Ideally, Migori could have been used to analyse the effect of the ecological classification on mos-quito estimates. Unfortunately, the pre-existing surveillance samples are located in the same ecological zone (electronic supplementary material, appendix F) making it impossible to simulate the effect of the ecological classification on the sample size/location optimization. For this reason we evalu-ated the effect of stratification on sample size using a different dataset (Uganda). This shows that 10–20% of mosquito col-lections randomly selected from strata are representative of the full survey. This result shows that stratification can be applied at any stage of the sampling campaign, and even if it was not considered at the initial (planning) phase, it can adaptatively inform the subsequent sampling phases or collections and optimize the sampling costs (subsequent sub-sampling of each strata). In addition, using the Uganda dataset, we have also shown that the ecological stratification improves model fitting, again representing a model feature that can be applied in both pre-analysis and post-analysis of sampling campaigns.
An element not considered in this analysis but that requires discussion is the temporal frequency and length of the sampling campaign. Designs for temporal sampling raise the same challenges as spatial designs, along with additional considerations. These include:
is it better to trap six times in each of two houses, or twice in each of six houses, or four times in each of three houses? And in the latter case, is it necessary that the nights should be at weekly intervals, or would the easier task of sampling over four consecutive nights yield a similar amount of information? Should the same ‘fixed’ houses be sampled on each occasion, or should a new set be chosen randomly on each occasion?(extracted from [62])
Answering these questions requires relatively lengthy longi-tudinal studies and a knowledge of Anopheles population dynamics. Fortnightly collections are common in mosquito sampling designs, and enable cost-effective descriptions of seasonality and variation in mosquito abundance [18]. On the other hand, positioning traps during peaks of mosquito abundance can significantly overestimate the rate of popu-lation increase and the level of abundance, and only sampling over two or more years may accurately account for cyclical fluctuations in vector abundance [62].
Our analysis provides an example of how to fully describe the assumptions, conditions and constraints of sampling strategies. We do not expect other researchers to precisely replicate our methodology, e.g. the use of four houses in 30 sampling locations depends on previous abundance analy-sis and may change when more information will be available (adaptive sampling). Instead we have shown how open-data sources and ecological information can be implemented in the initial steps of sampling design. Our literature review shows that the specifics of sampling design are poorly reported and we therefore suggest that even when sampling is based on expert-opinion decisions, a full description of the sampling design should be provided to make the sampling repeatable or comparable or usable for subsequent similar studies. For example, field constraints such as presence of the disease or vector or host, vegetation type and density, elevation, field hostility, logistic feasibility, potential interfer-ence, human proximity, breeding sites, and risk of trapping material theft [13] or the type of trap used [63], which often are the major influences in the sampling design, need to be declared and described. In fact, previous sampling cam-paigns are often used to inform future sampling design, and therefore standardization of sampling designs and protocols are now a priority.
Not only malaria studies lack sampling design infor-mation. For mosquito-borne arboviruses such as Dengue and West Nile Virus, or indeed other vectors such as ticks [64], sample size and locations can be solely based on economic and environmental constraints and/or expert-based decisions (see for example [65,66]), including national and sub-national sampling campaigns. In the Netherlands a national mosquito surveillance campaign [67] did stratify the sampling area based on land use and public health concerns (i.e. preferential sampling by oversampling urban areas) with number of locations depending on the pre-determined scale of the analysis. A similar, although less detailed, approach was taken in North Italy [68] for West Nile Virus. However, a recent study [69], employed an a priori G*Power analysis to determine the sample size, and then allocated the samples to (a) maximize the spatial spread-ing of mosquito sample sites, and (b) to sample at locations
that would reflect a large range of socioeconomic conditions, since the objective was to estimate the effect of socio-economic drivers in Aedes albopictus distribution. While conceptually similar to the one proposed here (although our goal was to find ecological homogeneous areas instead of mapping socioeconomic differences), the authors do not consider the spatial autocorrelation in their sample size and design (which for diseases transmission byAedes species is very important, see for example [27] and [70]), although they do use a maximum coverage approach.
Finally, our framework can be applied to other ecological studies. For example, Wang and colleagues [71] included a spatial autocorrelation index to improve the sampling design for crop acreage. This approach, however, does not model the spatial autocorrelation that is necessary to allocate spatial samples but simply tries to achieve independence between samples. Our framework may support systematic sampling when affected by spatial autocorrelation [72] or when geostatistical mapping is required [73].
In conclusion, big and open data and research outputs could enhance the power of ecological and genomic studies [3], facilitating the growth of complex and multidimensional algorithms. In the specific field of vector biology and genomics, there is an urgent need to establish standards for mosquito sampling design and description in scientific
reports. One of the first steps is to facilitate training and workshops [11] but also the improvement of publishing stan-dards (i.e. requiring authors to fully disclose the sampling design) in order to produce a collection of high quality and usable sampling designs along with their results.
Data accessibility.All data used in this research are available through the links described in the paper. Any additional data or R codes can be requested from the corresponding authors (L.S.).
Authors’ contributions.L.S., E.R.L., D.W. and M.J.D. conceived the study. L.S.D., A.V.C.E., A.E.-Y., B.I.K., J.M. and E.O. provided background information and data on the sampling sites and inspected the areas. L.S. performed the statistical analysis. L.S., E.R.L., D.W. and M.J.D. drafted the manuscript and all the authors contributed to its final version.
Competing interests.We declare we have no competing interests. Funding. This study was supported by funding from the Medical Research Council (grant no. MR/P02520X/1). This UK funded award is part of the EDCTP2 programme supported by the European Union. A.V.C.E. was also supported by Wellcome Trust (grant number 110430/Z/15/Z); and L.S. by EPSRC (grant number EP/ R01860X/1).
Acknowledgements.We thank Prof. Peter Diggle and Dr Michael Chipeta for the useful advices on the presented analyses. L.S. thanks the wonderful KEMRI/Kisumu (Kenya) staff for hosting the presentation and contributing to the discussions of this work.
References
1. Thorpe ASet al.2016 Introduction to the sampling designs of the National Ecological Observatory Network Terrestrial Observation System.Ecosphere
7, e01627. (doi:10.1002/ecs2.1627)
2. Golding N, Nunn MA, Purse BV. 2015 Identifying biotic interactions which drive the spatial distribution of a mosquito community.Parasit. Vectors8, 367. (doi:10.1186/s13071-015-0915-1) 3. Foley DH, Wilkerson RC, Rueda LM. 2009
Importance of the ‘what’, ‘when’, and ‘where’ of mosquito collection events.J. Med. Entomol.46, 717 – 722. (doi:10.1603/033.046.0401)
4. Daniels RFet al.2015 Modeling malaria genomics reveals transmission decline and rebound in Senegal.Proc. Natl Acad. Sci. USA112, 7067 – 7072. (doi:10.1073/pnas.1505691112)
5. Timmann Cet al.2012 Genome-wide association study indicates two novel resistance loci for severe malaria.Nature489, 443 – 446. (doi:10.1038/ nature11334)
6. Clarkson CS, Miles A, Harding NJ, Botta G, Lawniczak MK, Donnelly MJ, Kwiatkowski D, Genomes AG. 2017 The Anopheles Gambiae 1000 Genomes Project Phase 2: insecticide resistance, gene drive and genome variation in 1,142 malaria mosquitoes.Am. J. Trop. Med. Hyg.97, 223 – 224. 7. Wang JF, Jiang CS, Hu MG, Cao ZD, Guo YS, Li LF,
Liu TJ, Meng B. 2013 Design-based spatial sampling: theory and implementation.Environ. Model. Softw.40, 280 – 288. (doi:10.1016/j.envsoft. 2012.09.015)
8. McCann RSet al.2017 Assessing the spatial heterogeneity of malaria vectors in the context of
increasing vector control interventions.Am. J. Trop. Med. Hyg.95, 57.
9. McCann RSet al.2017 Assessment of the effect of larval source management and house improvement on malaria transmission when added to standard malaria control strategies in southern Malawi: study protocol for a cluster-randomised controlled trial.
BMC Infect. Dis.17, 639. (doi:10.1186/s12879-017-2749-2)
10. Rochlin I, Iwanejko T, Dempsey ME, Ninivaggi DV. 2009 Geostatistical evaluation of integrated marsh management impact on mosquito vectors using before-after-control-impact (BACI) design.Int. J. Health Geogr.8, 35. (doi:10.1186/1476-072x-8-35) 11. Regis LNet al.2013 Sustained reduction of the
dengue vector population resulting from an integrated control strategy applied in two Brazilian cities.PLoS ONE8, e67682. (doi:10.1371/journal. pone.0067682)
12. Bowman LR, Runge-Ranzinger S, McCall PJ. 2014 Assessing the relationship between vector indices and dengue transmission: a systematic review of the evidence.PLoS Negl. Trop. Dis.8, e2848. (doi:10. 1371/journal.pntd.0002848)
13. Roux E, Gaborit P, Romana CA, Girod R, Dessay N, Dusfour I. 2013 Objective sampling design in a highly heterogeneous landscape—characterizing environmental determinants of malaria vector distribution in French Guiana, in the Amazonian region.BMC Ecol.13, 45. (doi:10.1186/1472-6785-13-45)
14. Apostol BL, Black WCT, Reiter P, Miller BR. 1996 Population genetics with RAPD-PCR markers: the
breeding structure of Aedes aegypti in Puerto Rico.
Heredity (Edinb)76(Pt 4), 325 – 334. (doi:10.1038/ hdy.1996.50).
15. Jacob BG, Gu WD, Caamano EX, Novak RJ. 2009 Developing operational algorithms using linear and non-linear squares estimation in Python (R) for the identification of Culex pipiens and Culex restuans in a mosquito abatement district (Cook County, Illinois, USA).Geospat. Health3, 157 – 176. (doi:10. 4081/gh.2009.218)
16. Himeidan YE, Elzaki MM, Kweka EJ, Ibrahim M, Elhassan IM. 2011 Pattern of malaria transmission along the Rahad River basin, Eastern Sudan.Parasit. Vectors4, 109. (doi:10.1186/1756-3305-4-109) 17. Kweka EJ, Zhou GF, Lee MC, Gilbreath TM, Mosha F,
Munga S, Githeko AK, Yan GY. 2011 Evaluation of two methods of estimating larval habitat productivity in western Kenya highlands.Parasit. Vectors4, 110. (doi:10.1186/1756-3305-4-110) 18. Vanlalruia K, Senthilkumar N, Gurusubramanian G.
2014 Diversity and abundance of mosquito species in relation to their larval habitats in Mizoram, North Eastern Himalayan region.Acta Trop.137, 1 – 18. (doi:10.1016/j.actatropica.2014.04.026)
19. Le Goff G, Damiens D, Payet L, Ruttee AH, Jean F, Lebon C, Dehecq JS, Gouagna LC. 2016 Enhancement of the BG-sentinel trap with varying number of mice for field sampling of male and female Aedes albopictus mosquitoes.Parasit. Vectors9, 514. (doi:10.1186/s13071-016-1801-1) 20. Adde Aet al.2017 Spatial and seasonal dynamics
of anopheles mosquitoes in Saint-Georges de l’Oyapock, French Guiana: influence of
environmental factors.J. Med. Entomol.54, 597 – 605. (doi:10.1093/jme/tjx031)
21. Brugman VAet al.2017 How often do mosquitoes bite humans in southern England? A standardised summer trial at four sites reveals spatial, temporal and site-related variation in biting rates.
Parasit. Vectors10, 420. (doi:10.1186/s13071-017-2360-9)
22. Crocker W, Maute K, Webb C, French K. 2017 Mosquito assemblages associated with urban water bodies; implications for pest and public health threats.Landsc. Urban Plan.162, 115 – 125. (doi:10. 1016/j.landurbplan.2017.02.006)
23. Zittra C, Vitecek S, Obwaller AG, Rossiter H, Eigner B, Zechmeister T, Waringer J, Fuehrer HP. 2017 Landscape structure affects distribution of potential disease vectors (Diptera: Culicidae).Parasit. Vectors
10, 205. (doi:10.1186/s13071-017-2140-6) 24. Zio SD, Fontanella L, Ippoliti L. 2004 Optimal spatial
sampling schemes for environmental surveys.
Environ. Ecol. Stat.11, 397 – 414. (doi:10.1007/ s10651-004-4186-9)
25. Guisan A, Thuiller W. 2005 Predicting species distribution: offering more than simple habitat models.Ecol. Lett.8, 993 – 1009. (doi:10.1111/j. 1461-0248.2005.00792.x)
26. Lima AWS, Honorio NA, Codeco CT. 2016 Spatial clustering of Aedes aegypti (Diptera: Culicidae) and its impact on entomological surveillance indicators.
J. Med. Entomol.53, 343 – 348. (doi:10.1093/jme/ tjv187)
27. Azil AH, Bruce D, Williams CR. 2014 Determining the spatial autocorrelation of dengue vector populations: influences of mosquito sampling method, covariables, and vector control.J. Vector Ecol.39, 153 – 163. (doi:10.1111/j.1948-7134.2014. 12082.x)
28. Brus DJ, deGruijter JJ. 1997 Random sampling or geostatistical modelling? Choosing between design-based and model-design-based sampling strategies for soil (with discussion).Geoderma80, 1 – 44. (doi:10. 1016/S0016-7061(97)00072-4)
29. Chipeta M, Diggle PJ. 2015 Comment on article by Ferreira and Gamerman.Bayesian Anal.10, 737 – 739. (doi:10.1214/15-Ba944a) 30. Kao RHet al.2012 NEON terrestrial field
observations: designing continental-scale, standardized sampling.Ecosphere3, 1 – 17. (doi:10. 1890/Es12-00196.1)
31. Churcher Tet al.2017 malERA: an updated research agenda for insecticide and drug resistance in malaria elimination and eradication.PLoS Med.14, e1002450. (doi:10.1371/journal.pmed.1002450) 32. Hellwig E, Hijmans RJ. 2017 Spatio-temporal
variation in childhood growth in Nigeria: a comparison of aggregation and interpolation.
Int. J. Digit. Earth10, 1166 – 1176. (doi:10.1080/ 17538947.2017.1330905)
33. Chen J, Cao X, Peng S, Ren HR. 2017 Analysis and applications of GlobeLand30: a review.ISPRS Int. J. Geoinf.6, 230. (doi:10.3390/ijgi6080230) 34. Seddon AW, Macias-Fauria M, Long PR, Benz D,
Willis KJ. 2016 Sensitivity of global terrestrial
ecosystems to climate variability.Nature531, 229 – 232. (doi:10.1038/nature16986)
35. Fick SE, Hijmans RJ. 2017 WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas.Int. J. Climatol.37, 4302 – 4315. (doi:10. 1002/joc.5086)
36. Kilama Met al.2014 Estimating the annual entomological inoculation rate forPlasmodium falciparumtransmitted byAnopheles gambiae s.l.
using three sampling methods in three sites in Uganda.Malar. J.13, 111. (doi:10.1186/1475-2875-13-111)
37. Ribeiro Jr PJ, Diggle PJ. 2006 geoR: Package for Geostatistical Data Analysis an illustrative session.
Artif. Intell.1, 1 – 24.
38. Diggle PJ, Moraga P, Rowlingson B, Taylor BM. 2013 Spatial and spatio-temporal log-Gaussian Cox processes: extending the geostatistical paradigm.
Stat. Sci.28, 542 – 563. (doi:10.1214/13-Sts441) 39. Diggle P, Ribeiro PJ. 2007Model-based geostatistics.
New York, NY: Springer.
40. Hoeting JA, Davis RA, Merton AA, Thompson SE. 2006 Model selection for geostatistical models.Ecol. Appl.16, 87 – 98. (doi:10.1890/04-0576) 41. Simpson D, Illian JB, Lindgren F, Sørbye SH, Rue H.
2016 Going off grid: computationally efficient inference for log-Gaussian Cox processes.Biometrika
103, 49 – 70. (doi:10.1093/biomet/asv064) 42. Fujikoshi Y, Ulyanov VV, Shimizu R. 2011
Multivariate statistics: high-dimensional and large-sample approximations. Hoboken, NJ: Wiley. 43. Zhang X, Nansen C, Aryamanesh N, Yan G, Boussaid
F. 2015 Importance of spatial and spectral data reduction in the detection of internal defects in food products.Appl. Spectrosc.69, 473 – 480. (doi:10.1366/14-07672)
44. McLachlan, G. 2004Discriminant analysis and statistical pattern recognition. Hoboken, NJ: Wiley. 45. Sharma A, Paliwal KK. 2008 A gradient linear
discriminant analysis for small sample sized problem.Neural Process. Lett.27, 17 – 24. (doi:10. 1007/s11063-007-9056-7)
46. Diggle P, Lophaven S. 2006 Bayesian geostatistical design.Scand. J. Stat.33, 53 – 64. (doi:10.1111/j. 1467-9469.2005.00469.x)
47. Zimmerman DL. 2006 Optimal network design for spatial prediction, covariance parameter estimation, and empirical prediction.Environmetrics17, 635 – 652. (doi:10.1002/env.769)
48. Banerjee S, Gelfand AE, Finley AO, Sang H. 2008 Gaussian predictive process models for large spatial data sets.J. R. Stat. Soc. Series B Stat. Methodol.70, 825 – 848. (doi:10.1111/j.1467-9868.2008.00663.x) 49. Silver JB. 2007Mosquito ecology: field sampling
methods. Dordrecht, The Netherlands: Springer. 50. Chipeta M, Terlouw D, Phiri K, Diggle P. 2017
Inhibitory geostatistical designs for spatial prediction taking account of uncertain covariance structure.
Environmetrics28, e2425. (doi:10.1002/env.2425) 51. Chipeta MG, Kabaghe AN, McCann RS, Phiri KS, Van
Vugt M, Takken W, Terlouw DJ. 2017 Adaptive geostatistical sampling enables efficient identification of malaria hotspots in repeated
cross-sectional surveys in rural Malawi.Am. J. Trop. Med. Hyg.97, 105.
52. Chipeta MG, Terlouw DJ, Phiri KS, Diggle PJ. 2016 Adaptive geostatistical design and analysis for prevalence surveys.Spat. Stat.15, 70 – 84. (doi:10. 1016/j.spasta.2015.12.004)
53. Venables WN. 1999Modern applied statistics with PLUS, 3rd edn. New York City, NY: Springer. 54. Kaufman L, Rousseeuw PJ. 1990Finding groups in
data.An introduction to cluster analysis. Hoboken, NJ: Wiley.
55. Pettorelli N, Laurance WF, O’Brien TG, Wegmann M, Nagendra H, Turner W, Milner-Gulland EJ. 2014 Satellite remote sensing for applied ecologists: opportunities and challenges.J. Appl. Ecol.51, 839 – 848. (doi:10.1111/1365-2664.12261) 56. Reisen WK, Lothrop HD. 1999 Effects of sampling
design on the estimation of adult mosquito abundance.J. Am. Mosq. Control Assoc.15, 105 – 114.
57. Cardo MV, Rubio A, Junges MT, Vezzani D, Carbajo AE. 2018 Heterogeneous distribution ofCulex pipiens,Culex quinquefasciatusand their hybrids along the urbanisation gradient.Acta Trop.178, 229 – 235. (doi:10.1016/j.actatropica.2017.11.017) 58. Piovezan R, de Azevedo TS, Von Zuben CJ. 2012 Spatial evaluation of larvae of Culicidae (Diptera) from different breeding sites: application of a geospatial method and implications for vector control.Rev. Bras. Entomol.56, 368 – 376. (doi:10. 1590/S0085-56262012005000037)
59. Thomas SJet al.2015 Improving dengue virus capture rates in humans and vectors in Kamphaeng Phet Province, Thailand, using an enhanced spatiotemporal surveillance strategy.Am. J. Trop. Med. Hyg.93, 24 – 32. (doi:10.4269/ajtmh.14-0242) 60. World Health Organization. 2009Dengue: guidelines
for diagnosis, treatment, prevention and control. France, World Health Organization.
61. Blasi Cet al.2017 Ecosystem mapping for the implementation of the European Biodiversity Strategy at the national level: the case of Italy.
Environ. Sci. Policy78, 173 – 184. (doi:10.1016/j. envsci.2017.09.002)
62. Magbity EB, Lines JD. 2002 Spatial and temporal distribution ofAnopheles gambiaes.l. (Diptera: Culicidae) in two Tanzanian villages: implication for designing mosquito sampling routines.Bull. Entomol. Res.92, 483 – 488. (doi:10.1079/ Ber2002200)
63. Salazar FV, Chareonviriyaphap T, Grieco JP, Prabaripai A, Polsomboon S, Gimutao KA, Bangs MJ, Achee NL. 2017 Bg-Sentinel (Tm) trap efficacy as a component of proof-of-concept for push-pull control strategy for dengue vector mosquitoes.
J. Am. Mosq. Control Assoc.33, 293 – 300. (doi:10. 2987/16-6552.1)
65. Ng KC, Chaves LF, Tsai KH, Chuang TW. 2018 Increased adultAedes aegyptiandCulex quinquefasciatus(Diptera: Culicidae) abundance in a dengue transmission hotspot, compared to a coldspot, within Kaohsiung City, Taiwan.Insects9, 98. (doi:10.3390/insects9030098)
66. Montgomery BL, Shivas MA, Hall-Mendelin S, Edwards J, Hamilton NA, Jansen CC, McMahon JL, Warrilow D, van den Hurk AF. 2017 Rapid Surveillance for Vector Presence (RSVP): development of a novel system for detectingAedes aegyptiandAedes albopictus.PLoS Negl. Trop. Dis.
11, e0005505. (doi:10.1371/journal.pntd.0005505) 67. Ibanez-Justicia A, Stroo A, Dik M, Beeuwkes J,
Scholte EJ. 2015 National Mosquito (Diptera: Culicidae) Survey in The Netherlands 2010 – 2013.
J. Med. Entomol.52, 185 – 198. (doi:10.1093/jme/ tju058)
68. Calzolari Met al.2015 West Nile virus surveillance in 2013 via mosquito screening in northern Italy and the influence of weather on virus circulation.
PLoS ONE10, e0140915. (doi:10.1371/journal.pone. 0140915)
69. Whiteman A, Delmelle E, Rapp T, Chen S, Chen G, Dulin M. 2018 A novel sampling method to measure socioeconomic drivers ofAedes albopictus
distribution in Mecklenburg County, North Carolina.
Int. J. Environ. Res. Public Health15, 2179. (doi:10. 3390/ijerph15102179)
70. Williams CR, Long SA, Webb CE, Bitzhenner M, Geier M, Russell RC, Ritchie SA. 2007Aedes aegypti
population sampling using BG-Sentinel traps in
North Queensland Australia: statistical considerations for trap deployment and sampling strategy.J. Med. Entomol.44, 345 – 350. (doi:10.1603/0022-2585(2007)44[345:Aapsub]2.0.Co;2)
71. Wang D, Zhou QB, Yang P, Chen ZX. 2018 Design of a spatial sampling scheme considering the spatial autocorrelation of crop acreage included in the sampling units.J. Integr. Agric.17, 2096 – 2106. (doi:10.1016/S2095-3119(17)61882-3)
72. Strand GH. 2017 A study of variance estimation methods for systematic spatial sampling.Spat. Stat.
21, 226 – 240. (doi:10.1016/j.spasta.2017.06.008) 73. Falk MG, Denham RJ, Mengersen KL. 2011 Spatially
stratified sampling using auxiliary information for geostatistical mapping.Environ. Ecol. Stat.18, 93 – 108. (doi:10.1007/s10651-009-0122-3)
ro
yalsocietypublishing.org/journal/rsif
J.
R.
Soc.
Interfa
ce
16
:
20180941