University of Groningen
Rok Regulates yuaB Expression during Architecturally Complex Colony Development of
Bacillus subtilis 168
Kovacs, Akos T.; Kuipers, Oscar P.
Published in:
Journal of Bacteriology
DOI:
10.1128/JB.01170-10
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from
it. Please check the document version below.
Document Version
Publisher's PDF, also known as Version of record
Publication date:
2011
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):
Kovacs, A. T., & Kuipers, O. P. (2011). Rok Regulates yuaB Expression during Architecturally Complex
Colony Development of Bacillus subtilis 168. Journal of Bacteriology, 193(4), 998-1002.
https://doi.org/10.1128/JB.01170-10
Copyright
Other than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policy
If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons the number of authors shown on this cover page is limited to 10 maximum.
0021-9193/11/$12.00 doi:10.1128/JB.01170-10
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Rok Regulates
yuaB
Expression during Architecturally Complex
Colony Development of
Bacillus subtilis
168
䌤
#
A
´ kos T. Kova´cs
1and Oscar P. Kuipers
1,2*
Molecular Genetics Group, Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen,1and
Kluyver Centre for Genomics of Industrial Fermentation,2Kerklaan 30, 9751NN Haren, Netherlands
Received 30 September 2010/Accepted 11 November 2010
Transcriptome analysis of a Bacillus subtilis rok strain that showed reduced complex colony structure formation revealed significant downregulation of theyuaB gene. Overexpression ofyuaB restored structure formation in therokstrain. We show that transcription ofyuaBis indirectly regulated by Rok, independently from its previously described AbrB-dependent regulation.
The Gram-positive model organismBacillus subtilisis capa-ble of forming surface-associated communities of cells in a matrix of extracellular polymers organized in complex struc-tures (3). The main structural components ofB. subtilis bio-films, i.e., a protein component (TasA in theyqxM-sipW-tasA
operon) and an exopolysaccharide (EPS; produced by proteins encoded by the epsA-epsO operon), are produced by only a small proportion of cells (5). The aerial extensions formed in mature biofilms, often described as “fruiting body-like” struc-tures, present preferential sites for sporulation (3, 23).B. sub-tilisbiofilm development involves many regulators that deter-mine the cell fate and therefore the composition of the biofilm. Repression of biofilm genes by the master regulator SinR is relieved upon Spo0A-dependent expression of SinI (5). Simi-larly, Spo0A-regulated AbrB represses theyqxM operon (7). Besides theyqxMandepsoperons, several genes/operons have been described to be involved in the proper development of biofilms (4, 6, 13). The genesyuaBand yvcAwere identified through analysis of the role of phosphorylated DegU in biofilm development (14, 24). YuaB is a small secreted protein regu-lated by AbrB, and its expression depends on the biosynthesis of the polysaccharide component of the biofilm inB. subtilis
NCIB3610 (25).
Most studies of B. subtilis biofilm development have been performed in wild isolates (e.g., NCIB3610) that produce wrin-kled structures during architecturally complex colony develop-ment on solid surfaces or during pellicle formation at the air-liquid interface (3). It has been shown that certain labora-tory strains of B. subtilis (e.g., 168 1A700) are capable of forming architecturally complex colonies with vein-like struc-tures formed by elevated bundles of cells (23). Domesticated strains ofB. subtilis(e.g., PY79) have been shown to contain mutations in genes important for swarming motility as well as colony architecture formation (12, 22). Unlike 168 1A700,
PY79 produces flat colonies with no apparent bundle forma-tion (3, 23). Sequencing of thedegQ,sfp, andswrAloci of strain 168 1A700 revealed that these genes contain mutations iden-tical to those found in the PY79 strain, indicating that 168 1A700 must differ from PY79 at at least one other genetic locus.
Here, we examine the role of the transcription factor Rok in architecturally complex colony development byB. subtilis168 1A700 (Table 1). Rok was previously identified as a repressor ofcomK (11), the master regulator of competence, but Rok also reduces the transcription of a family of genes that specify membrane-localized and secreted proteins, including a number of genes that encode products with antibiotic activity (1). Strains (Table 1) were grown in TY medium (1% Bacto tryp-tone, 0.5% Bacto yeast, 0.5% NaCl, and 0.1 mM Mncl2), and colonies were obtained by spotting 2l of an overnightBacillus
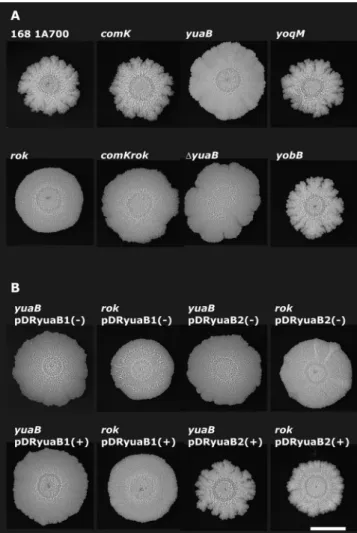
culture on 2⫻SG medium (13) solidified by 1.5% agar. Mu-tation inrokresulted in the reduction of theB. subtiliscolony structures (Fig. 1A), without disturbing the expression of genes important for the production of biofilm matrix, as was ob-served insinIorepsmutant strains (Fig. 2). Mutation inrok
also tempers pellicle formation (see Fig. S1 in the supplemen-tal material). The transcription factor Rok therefore raises the number of known transcription factors inB. subtilisthat influ-ence architecturally complex colony formation in addition to regulating various processes within the cells. Examination of genes that are affected byrokduring complex colony develop-ment would therefore likely reveal factors necessary for the formation of structures but not matrix production.
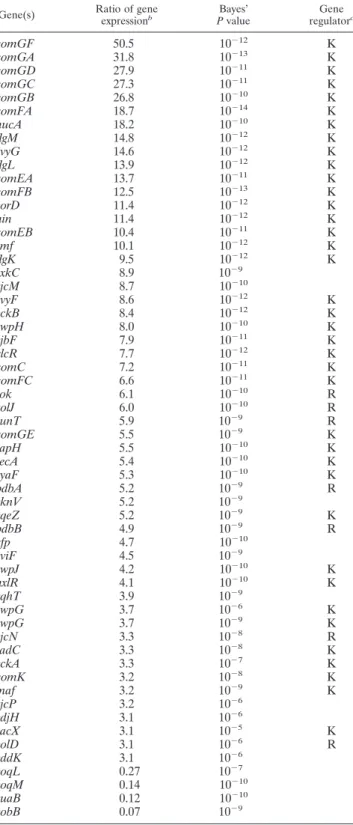
Transcriptome analysis of colonies of the rok mutant in comparison with those of 168 1A700 revealed the upregulation of previously identified genes (1) regulated directly by Rok or indirectly by ComK in liquid cultures (Table 2). The yuaB,
yobB, andyoqLMgenes were found to be significantly down-regulated in the rok strain. Even though many ComK-regu-lated genes were differentially expressed in the rok mutant, colony structure formation was not affected by acomK muta-tion (Fig. 1A). Although the list of upregulated genes (See Table 2) excludes the previously identified ComK-regulated genes in transcriptome analysis when colonies of acomKsingle mutant and acomK rokdouble mutant are compared, it con-tains the same downregulated genes (data not shown).
* Corresponding author. Mailing address: Molecular Genetics, Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Nijenborgh 7, 9747 AG Groningen, Neth-erlands. Phone: (31) 50 3632093. Fax: (31) 50 3632348. E-mail: [email protected].
# Supplemental material for this article may be found at http://jb .asm.org/.
䌤Published ahead of print on 19 November 2010.
To examine the genes that were downregulated in therok
strain under architecturally complex colony development, mu-tations were introduced into theyuaB,yobB, andyoqMgenes. Only a mutation in yuaB resulted in a reduction in colony structure, identical to that of therok mutant, while theyobB
andyoqMmutant strains had a colony structure similar to that of 168 1A700 (Fig. 1A). The BFA239 mutant of theyuaBgene, constructed previously by theBacillus subtilisgene Functional Analysis Project (15), was used in our experiments, since the deletion mutant ofyuaB (14) produced similar results (Fig. 1A). The phenotypic similarity ofyuaB and rok strains with regard to colony structure suggests that Rok affects colony development through alteration of the transcription ofyuaB.
To test whetheryuaBis the sole gene regulated by Rok that is required for complex colony architecture, we overexpressed
yuaBin therokmutant using 1 mM isopropyl--D -1-thiogalacto-pyranoside (IPTG). High expression of yuaB from the hy-perspank promoter in pDR111 restored complex colony struc-ture in bothyuaB and rok strains, while expression ofyuaB
from the spank promoter in pDR110 did not complement mutation in either yuaB orrok (Fig. 1B). Expression levels from the hyperspank promoter are around 7 times higher than those from the spank promoter when induced with IPTG (20). This suggests that a high level of expression is necessary for the
proper function of YuaB. Furthermore, the complementation of therokmutation by expression of yuaBsuggests that Rok affects the development ofB. subtiliscolony structure solely by regulating the transcription ofyuaB.
To address whether Rok regulatesyuaBexpression directly, gel mobility shift assays were performed using various amounts of purified Rok protein (1) and the promoter regions ofcomK
andyuaB (for details, see the supplemental material). While Rok bound to its previously identified target promoter of
comK, no DNA binding was observed inin vitrogel mobility shift assays when Rok was incubated with the promoter region of yuaB (see Fig. S2 in the supplemental material). In vivo
binding of Rok to theyuaBpromoter was also not observed in chromatin immunoprecipitation experiments (W. K. Smits, per-sonal communication). Therefore, Rok likely regulatesyuaB in-directly through a transcription factor that still needs to be identified. Extensive investigation of the differentially regu-lated genes in our transcriptome analysis did not reveal a
FIG. 1.B. subtiliscomplex colony structure depends on Rok-regu-latedyuaBtranscription. Strains were spotted on 2⫻SG plates and incubated for 3 days. The scale bar in the lower right corner represents 10 mm. (A) Colony morphology of variousB. subtilisstrains. (B)B. subtilis yuaBand rokmutants transformed with pDRyuaB1 (Pspank
-yuaB) and pDRyuaB2 (Phyperspank-yuaB) constructs in the absence
(up-per panels) or presence (lower panels) of 1 mM IPTG. TABLE 1. Strains and plasmids used in this study
Strain or plasmid Relevant genotype or descriptiona
Reference or source
B. subtilisstrainsb
168 1A700 trpC2/parental strain in this study
Bacillus Genetic Stock Center
rokmutant 168rok::Kmr 1
rokSp mutant 168rok::Spr
1
comKmutant 168comK::Spr 10
rok comKmutant 168rok::Kmr
comK::Spr
This study
yuaBmutant 168 BFA239yuaB::Eryr 15
yoqMmutant 168yoqM::Eryr This study
yobBmutant 168yobB::Eryr
This study
⌬yuaBmutant 168⌬yuaB::Cmr 14
abrBmutant 168abrB::Eryr 21
abrB rokmutant 168abrB::Eryr rok::Kmr
This study
degSUmutant 168⌬degSU::aphA3Kmr 19 degU146mutant 168degU146Kmr
8
degU32mutant 168degU(Hy)32Kmr 16 degU32 rokmutant 168degU(Hy)32Kmrrok::Spr This study sinImutant 168sinI::Spr
9
sinI rokmutant 168sinI::Sprrok::Kmr This study
sinRmutant 168sinR::Spr 21
epsGmutant 168epsG::pBL601 Spr
25
epsG rokmutant 168epsG::pBL601 Sprrok::Kmr This study
Plasmids
pSG1151 AmprCmrgfpmut1 17
pSGyuaB PyuaBcloned into pSG1151
Ampr
Cmr
This study
pDR110 AmprSprlacIⱊamyE amyE⬘
Pspank
Gift from D. Rudner
pDR111 AmprSprlacIⱊamyE amyE⬘
Phyperspank
Gift from D. Rudner
pDRyuaB1 yuaBcloned into pDR110
AmprSpr
This study
pDRyuaB2 yuaBcloned into pDR111
AmprSpr
This study
aKmr, kanamycin resistant; Spr, spectinomycin resistant; Eryr, erythromycin resistant; Cmr, chloramphenicol resistant, Ampr, ampicillin resistant.
bB. subtilisstrains were obtained by transforming genomic DNA of various
strains into parental strain 168 1A700.
candidate transcription factor that could play a role in the regulation ofyuaBtranscription.
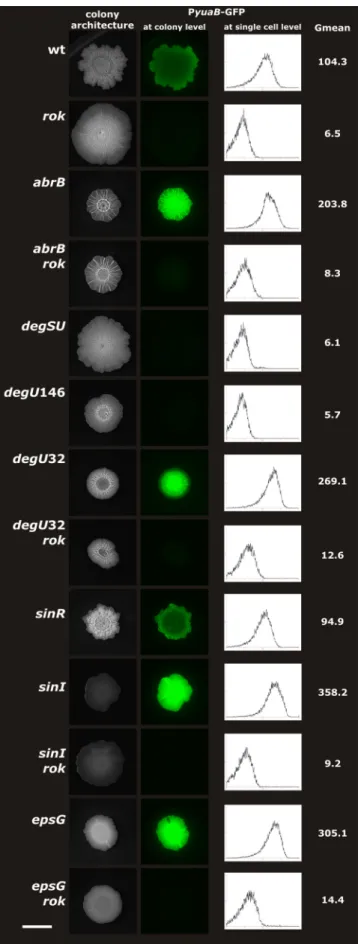
The transcriptional regulation of yuaB was studied using strains harboring ayuaB promoter-green fluorescent protein (GFP) fusion. Fluorescence was detected at both the colony level using a macrozoom fluorescence microscope and at the single-cell level using a cytometer (Fig. 2). Expression ofyuaB
was reduced in therokstrain, as observed in the array exper-iments (Fig. 2). Similarly,yuaB expression was diminished in the pellicles of the rok mutant compared to the pellicles of strain 168 1A700 (see Fig. S1 in the supplemental material). In agreement with previous reports (14, 24, 25), a mutation in
abrBenhanced, while deletion of thedegSUgenes reduced, the expression ofyuaB in complex colonies (Fig. 2). Abol-ishing (degU146) or increasing (degU32) the phosphoryla-tion of DegU resulted in reducphosphoryla-tion or elevaphosphoryla-tion ofyuaB tran-scription, respectively. When arok mutation was introduced into either theabrBordegU32background, minor changes in colony structure were observed. However, the expression of
yuaB was reduced in the abrB rok and degU32 rok double mutants to the same extent as whenrok alone was mutated. Thus, the transcriptional regulation ofyuaB by Rok is inde-pendent of the previously described regulatory pathways (14, 25). When the promoter-reporter fusion was introduced into a
sinRbackground, the reporter activity driven by theyuaB pro-moter region was comparable to that of the 168 1A700 strain. Previously, SinR was found to negatively affectyuaBregulation inB. subtilisNCIB3610 through the reduction of EPS produc-tion (25). In contrast, here we find that mutaproduc-tion of epsG, causing disruption of EPS production, increased expression of
yuaB in B. subtilis168 1A700 (Fig. 2). Similarly, deletion of
sinI, which results in a colony morphology comparable to that of the epsG strain, increased yuaB expression. Thus, in B. subtilis168 1A700, the absence of EPS has a positive rather than a negative effect on the transcription ofyuaB. Domesti-cated strains and undomestiDomesti-cated isolates were found previ-ously to differ in gene regulation during biofilm development; e.g., heterogeneity ofsrfAAexpression differs between strains NCIB3610 and 168 1A700 (18), while colony morphology de-pends on distinct polymers in various wild isolates, indicating that EPS has an important role in strain NCIB3610 (2) and poly-␥-DL-glutamate in strain RO-FF-1 (22). How the absence or presence of EPS regulates the expression ofyuaBin
differ-FIG. 2. Expression ofyuaBis altered inB. subtilisby Rok, AbrB, DegU, SinI, and EPS. Strains were spotted on 2⫻ SG plates and
incubated for 3 days. Fluorescence was detected in complex colonies using an Olympus MVX10 macrozoom fluorescence microscope equipped with a PreciseExcite LED fluorescence illuminator (470 nm), a GFP filter set (excitation at 460/480 nm and emission at 495/540 nm), and an Olympus XM10 monochrome camera. For the single-cell anal-ysis of complex colonies, a colony was scraped from the plate using a scalpel and homogenized for 1 min using a mini-beadbeater-8 (Bio-spec, Bartlesville, OK), and fluorescence was measured using a Coulter Epics XL-MCL flow cytometer (Beckman Coulter, Mijdrecht, Neth-erlands) operating an argon laser (488 nm). For each sample, at least 20,000 cells were analyzed using WinMDI 2.8 (http://facs.scripps.edu /software.html). The relative numbers of cells (0 to 75) are indicated on theyaxis of the graphs, and their relative fluorescence levels (0 to 104) are indicated on the x axis on a logarithmic scale. “Gmean”
indicates the mean fluorescence of PyuaB-GFP-containing cells at the single-cell level. The scale bar in the lower left corner represents 10 mm.
ent strains is not yet understood (25) and remains to be inves-tigated. However, deletion ofrokabolishes the highyuaB ex-pression in epsG or sinI mutants, demonstrating that the regulation ofyuaBby Rok depicted here is distinct from the ones previously described.
In summary, we have shown thatyuaBtranscription is influ-enced by the transcription factor Rok during architecturally complex colony development ofB. subtilisindependently from the previously described regulators.
Microarray data accession number. Microarray data have been deposited in the Gene Expression Omnibus database (GSE22370).
We thank Nicola Stanley-Wall for the kind gift ofB. subtilisstrains and David Rudner for plasmids pDR110 and pDR111. We also thank Claire E. Price for assisting with the English corrections and Jan Willem Veening for critical reading of the manuscript.
A.T.K. was financially supported by grant 818.02.004 from the ALW-NWO Open Programme. This project was carried out within the re-search program of the Kluyver Centre for Genomics of Industrial Fermentation, which is part of the Netherlands Genomics Initiative/ Netherlands Organization for Scientific Research.
REFERENCES
1.Albano, M., et al.2005. The Rok protein ofBacillus subtilisrepresses genes for cell surface and extracellular functions. J. Bacteriol.187:2010–2019. 2.Branda, S. S., F. Chu, D. B. Kearns, R. Losick, and R. Kolter.2006. A major
protein component of theBacillus subtilisbiofilm matrix. Mol. Microbiol. 59:1229–1238.
3.Branda, S. S., J. E. Gonzalez-Pastor, S. Ben-Yehuda, R. Losick, and R. Kolter.2001. Fruiting body formation byBacillus subtilis. Proc. Natl. Acad. Sci. U. S. A.98:11621–11626.
4.Branda, S. S., et al.2004. Genes involved in formation of structured multi-cellular communities byBacillus subtilis. J. Bacteriol.186:3970–3979. 5.Chai, Y., F. Chu, R. Kolter, and R. Losick.2008. Bistability and biofilm
formation inBacillus subtilis. Mol. Microbiol.67:254–263.
6.Chai, Y., R. Kolter, and R. Losick.2009. A widely conserved gene cluster required for lactate utilization inBacillus subtilisand its involvement in biofilm formation. J. Bacteriol.191:2423–2430.
7.Chu, F., et al.2008. A novel regulatory protein governing biofilm formation inBacillus subtilis. Mol. Microbiol.68:1117–1127.
8.Dahl, M. K., T. Msadek, F. Kunst, and G. Rapoport.1991. Mutational analysis of theBacillus subtilisDegU regulator and its phosphorylation by the DegS protein kinase. J. Bacteriol.173:2539–2547.
9.Hahn, J., A. Luttinger, and D. Dubnau.1996. Regulatory inputs for the synthesis of ComK, the competence transcription factor ofBacillus subtilis. Mol. Microbiol.21:763–775.
10.Hamoen, L. W., W. K. Smits, A. de Jong, S. Holsappel, and O. P. Kuipers. 2002. Improving the predictive value of the competence transcription factor (ComK) binding site inBacillus subtilisusing a genomic approach. Nucleic Acids Res.30:5517–5528.
11.Hoa, T. T., P. Tortosa, M. Albano, and D. Dubnau.2002. Rok (YkuW) regulates genetic competence inBacillus subtilisby directly repressingcomK. Mol. Microbiol.43:15–26.
12.Kearns, D. B., F. Chu, R. Rudner, and R. Losick.2004. Genes governing swarming inBacillus subtilisand evidence for a phase variation mechanism controlling surface motility. Mol. Microbiol.52:357–369.
13.Kobayashi, K.2007.Bacillus subtilispellicle formation proceeds through genetically defined morphological changes. J. Bacteriol.189:4920–4931. 14.Kobayashi, K.2007. Gradual activation of the response regulator DegU
controls serial expression of genes for flagellum formation and biofilm for-mation inBacillus subtilis. Mol. Microbiol.66:395–409.
15.Kobayashi, K., et al.2003. EssentialBacillus subtilisgenes. Proc. Natl. Acad. Sci. U. S. A.100:4678–4683.
16.Kunst, F., T. Msadek, J. Bignon, and G. Rapoport.1994. The DegS/DegU and ComP/ComA two-component systems are part of a network controlling degradative enzyme synthesis and competence inBacillus subtilis. Res. Mi-crobiol.145:393–402.
17.Lewis, P. J., and A. L. Marston.1999. GFP vectors for controlled expression and dual labelling of protein fusions inBacillus subtilis. Gene227:101–110. 18.Lopez, D., H. Vlamakis, R. Losick, and R. Kolter.2009. Paracrine signaling
in a bacterium. Genes Dev.23:1631–1638.
19.Msadek, T., et al.1990. Signal transduction pathway controlling synthesis of a class of degradative enzymes inBacillus subtilis: expression of the regula-tory genes and analysis of mutations indegSanddegU. J. Bacteriol.172:824– 834.
TABLE 2. Summary of transcriptional changes in biofilms of the
B. subtilis rokmutant compared toB. subtilis168 1A700a Gene(s) Ratio of gene
expressionb Bayes’ Pvalue Gene regulatorc comGF 50.5 10⫺12 K comGA 31.8 10⫺13 K comGD 27.9 10⫺11 K comGC 27.3 10⫺11 K comGB 26.8 10⫺10 K comFA 18.7 10⫺14 K nucA 18.2 10⫺10 K flgM 14.8 10⫺12 K yvyG 14.6 10⫺12 K flgL 13.9 10⫺12 K comEA 13.7 10⫺11 K comFB 12.5 10⫺13 K yorD 11.4 10⫺12 K nin 11.4 10⫺12 K comEB 10.4 10⫺11 K smf 10.1 10⫺12 K flgK 9.5 10⫺12 K yxkC 8.9 10⫺9 yjcM 8.7 10⫺10 yvyF 8.6 10⫺12 K yckB 8.4 10⫺12 K ywpH 8.0 10⫺10 K yjbF 7.9 10⫺11 K glcR 7.7 10⫺12 K comC 7.2 10⫺11 K comFC 6.6 10⫺11 K rok 6.1 10⫺10 R yolJ 6.0 10⫺10 R sunT 5.9 10⫺9 R comGE 5.5 10⫺9 K rapH 5.5 10⫺10 K recA 5.4 10⫺10 K yyaF 5.3 10⫺10 K bdbA 5.2 10⫺9 R yknV 5.2 10⫺9 yqeZ 5.2 10⫺9 K bdbB 4.9 10⫺9 R efp 4.7 10⫺10 yviF 4.5 10⫺9 ywpJ 4.2 10⫺10 K hxlR 4.1 10⫺10 K yqhT 3.9 10⫺9 ywpG 3.7 10⫺6 K ywpG 3.7 10⫺9 K yjcN 3.3 10⫺8 R radC 3.3 10⫺8 K yckA 3.3 10⫺7 K comK 3.2 10⫺8 K maf 3.2 10⫺9 K yjcP 3.2 10⫺6 ydjH 3.1 10⫺6 sacX 3.1 10⫺5 K yolD 3.1 10⫺6 R yddK 3.1 10⫺6 yoqL 0.27 10⫺7 yoqM 0.14 10⫺10 yuaB 0.12 10⫺10 yobB 0.07 10⫺9
aStrains were spotted on 2⫻SG plates and incubated for 3 days. Four
inde-pendent biological replicates, each containing 16 complex colonies, were har-vested for total RNA isolation. Array studies were performed as described in the supplemental material. Genes with a Bayes’Pvalue below 10⫺4with at least 3-fold differential expression were considered to be significantly affected.
bCalculated as expression in therokstrain to that in 168 1A700.
cK and R represent ComK and Rok, respectively, which were reported
pre-viously to regulate genes based on transcriptome analyses (1, 10).
20.Quisel, J. D., W. F. Burkholder, and A. D. Grossman.2001.In vivoeffects of sporulation kinases on mutant Spo0A proteins inBacillus subtilis. J. Bacte-riol.183:6573–6578.
21.Smits, W. K., et al.2005. Stripping Bacillus: ComK auto-stimulation is responsible for the bistable response in competence development. Mol. Microbiol.56:604–614.
22.Stanley, N. R., and B. A. Lazazzera.2005. Defining the genetic differences between wild and domestic strains ofBacillus subtilisthat affect poly-␥ -DL-glutamic acid production and biofilm formation. Mol. Microbiol.57:1143–1158.
23.Veening, J.-W., O. P. Kuipers, S. Brul, K. J. Hellingwerf, and R. Kort.2006. Effects of phosphorelay perturbations on architecture, sporulation, and spore resistance in biofilms ofBacillus subtilis. J. Bacteriol.188:3099–3109. 24.Verhamme, D. T., T. B. Kiley, and N. R. Stanley-Wall.2007. DegU co-ordinates multicellular behaviour exhibited byBacillus subtilis. Mol. Micro-biol.65:554–568.
25.Verhamme, D. T., E. J. Murray, and N. R. Stanley-Wall.2009. DegU and Spo0A jointly control transcription of two loci required for complex colony development byBacillus subtilis. J. Bacteriol.191:100–108.